塔里木河下游多枝檉柳幼苗生長和光合生理指標對輸水漫溢的響應特征
2016-12-17 02:12:09王明慧馬曉東張瑞群邢旭明
西北植物學報 2016年10期
關鍵詞:生長
王明慧,馬曉東,張瑞群,邢旭明
(新疆師范大學 生命科學學院,新疆特殊環境物種保護與調控生物學重點實驗室,烏魯木齊 830054)
?
塔里木河下游多枝檉柳幼苗生長和光合生理指標對輸水漫溢的響應特征
王明慧,馬曉東*,張瑞群,邢旭明
(新疆師范大學 生命科學學院,新疆特殊環境物種保護與調控生物學重點實驗室,烏魯木齊 830054)
以塔里木河下游天然漫溢區多枝檉柳幼苗為研究對象,分析連續漫溢干擾下多枝檉柳幼苗生長和種群變化特征,以及不同漫溢強度下幼苗的光合特性。結果表明:(1)在連續2次較強輸水漫溢的干擾下,多枝檉柳幼苗種群密度從2013年3月到2014年7月呈先增后降再增的趨勢,而其冠面積、株高和基徑則呈先降后增再降的趨勢。(2)在第2次輸水漫溢較弱的Ⅲ監測區,多枝檉柳幼苗種群密度呈先增加后逐漸降低的趨勢,而其冠面積、株高和基徑則呈先降低后逐漸增加的趨勢。(3)隨第2次輸水漫溢量逐漸減弱,多枝檉柳幼苗葉片相對含水量呈先降后增的趨勢,葉綠素含量、實際光化學量子產量(ΦPSⅡ)、光化學猝滅系數(qp)均呈逐漸降低的趨勢,而非光化學猝滅系數(NPQ)呈逐漸增加的趨勢,但潛在最大光化學量子效率(Fv/Fm)總體處于相對適宜的狀態。研究認為,連續2次較強的輸水漫溢條件雖能夠使生長季期間漫溢區多枝檉柳幼苗種群得到更新,但同時也導致大量幼苗受淹死亡,從而使漫溢區的多枝檉柳幼苗種群不能形成一定的規模;建議采取強弱交替的方式進行生態輸水,既能提高新發生幼苗的存活率,又能使幼苗穩定的生長。
多枝檉柳;漫溢干擾;種群;幼苗生長;葉綠素熒光
塔里木河下游地處極端干旱區,其荒漠河岸植被群落主要依靠人工生態輸水維系[1]。在生態輸水過程中,通過生態閘或河道人工缺口的作用在河岸兩側一定范圍內出現漫溢現象,河水漫溢作為河流生態系統的一個重要的環境干擾因子,是干旱和半干旱區河岸林生態系統的主要“組建者”之一[2]。白元等采用點格局分析方法對漫溢干擾下胡楊(PopuluseuphraticaOliv.)幼苗庫在空間分布格局上的變化及分布規律進行了研究,表明塔里木河下游胡楊幼樹主要分布在生態輸水的河道附近和河水漫溢區[3]。徐海量等研究表明洪水漫溢干擾對半干旱區域河岸林物種豐富度具一定的影響[4]。
多枝檉柳(TamarixramosissimaL.)廣泛分布于中國西北部平原荒漠、沙地和鹽堿地,具有很強的耐鹽、耐旱、耐沙埋的特性,作為塔里木河下游荒漠河岸林灌木層建群種,它對該地區河岸林的穩定起重要作用[5,6]。隨著河水消退,多枝檉柳以種子散播的方式,沿塔里木河流兩岸的漫溢帶發生大量幼苗,形成新的種群。多枝檉柳幼苗期是植物單株存活和種群擴展的決定性生活期,它們在極端干旱環境中發生和定居要更為艱難[7]。以往有關學者對多枝檉柳幼苗的研究主要集中在其幼苗生物量、形態、生理和根系分布對水分脅迫的響應上,以及其種群的空間分布格局方面[8-11],也有學者對洪水漫溢區多枝檉柳實生苗的更新進行了研究[12]。雖然趙振勇等[12]對漫溢干擾下多枝檉柳實生幼苗的密度情況也進行了考察,但主要是集中在1次漫溢后的效應,而有關連續漫溢干擾下多枝檉柳幼苗種群生長的動態特征,以及不同漫溢干擾強度后續將對多枝檉柳幼苗光合生長產生和影響方面的研究較少。基于此,為探究漫溢干擾作用下多枝檉柳幼苗種群是否會形成一定的規模效應以及不同漫溢強度下多枝檉柳幼苗的光合生理狀態,本研究對連續漫溢干擾下多枝檉柳幼苗種群地上形態特征和種群密度,以及不同漫溢強度下多枝檉柳幼苗的葉綠素熒光特征進行了探討,旨在從個體和種群水平揭示漫溢干擾對多枝檉柳幼苗生存狀態的影響,進而為塔里木河下游荒漠河岸林的更新生長和生態恢復提供科學依據。
1 研究區概況與方法
1.1 研究區概況
研究區位于塔里木河下游英蘇斷面,地處塔克拉瑪干沙漠和庫魯克沙漠之間,屬典型的極端干旱溫帶大陸氣候,區域內荒漠河岸林生態系統極為脆弱和不穩定。研究區內年降水量僅17.4~42.0 mm,年蒸潛在發量2 500~3 000 mm,氣候極度干燥,風沙危害嚴重。自20世紀50年代以來,由于人類不合理的水土資源開發,又加之1972年大西海子水庫的興建,使塔里木河下游321 km河道斷流,地下水位大幅度下降,塔里木河下游的荒漠河岸林生態系統退化嚴重,檉柳群落逐漸斑塊化,呈不均勻的條帶、斑塊狀分布。自2000年4月開始向塔里木河下游實施生態輸水工程,由于水量較大,從而出現了較大范圍的河水漫溢現象,河水消退的地方常常出現大量茂密的植被,河水漫溢區萌生了多枝檉柳和胡楊的實生苗以及黑果枸杞(LyciumruthenicumMurr.)、疏葉駱駝刺(AlhagisparsifoliaShap.)、鈴鐺刺(Halimodendronhalodendron(Pall.) Voss)和花花柴(Kareliniacaspia(Pall.) Less.)等一年生草本植物。
1.2 樣地設置
2013年3月底在塔里木河下游英蘇斷面河水漫溢區,選擇3個具有多枝檉柳分布的典型區域設置為幼苗庫監測點,分別標記為Ⅰ、Ⅱ和Ⅲ,每個監測區設置3個10 m×10 m調查樣方。2012年10月底和2013年8月初河水漫溢發生,形成一定面積的漫溢區,并于2013年11月對3個監測點處的漫溢情況進行調查,調查數據作為各監測點漫溢情況的強弱指標。其中,Ⅰ監測區水深1 m、漫溢帶最大寬度10 m;Ⅱ監測區水深0.15 m、漫溢帶最大寬度3 m;Ⅲ監測區無水淹情況、土壤含水量為22.8%。在2013年3月、2013年7月、2014年3月、2014年7月和2014年11月,分別對3個監測區9個樣方中多枝檉柳幼苗種群的株高、冠幅、基徑和密度以及0~150 cm土層土壤含水量進行了連續調查取樣。同時,為監測各漫溢干擾下幼苗的光合生理狀態,于2014年7月中旬植物生長旺季,對各監測區多枝檉柳幼苗葉綠素熒光參數日變化和葉綠素含量等指標進行了測定。
1.3 測定指標及方法
1.3.1 葉綠素熒光數據的采集與處理 于2014年7月中旬分別在Ⅰ、Ⅱ和Ⅲ3塊監測區每個調查樣方中隨機選取20株生長狀態相近且健康的多枝檉柳幼苗,于晴好的天氣下,采用調制葉綠素熒光儀(JUNIOR PAM)野外實地測定葉綠素熒光數據。每日于8:00~20:00每隔4 h測定1次光適應下最大熒光值Fm′、初始熒光Fo′和實時熒光值F′,每個時段重復6次,并于采樣當天凌晨4:00測量暗適應后葉片的Fo和Fm。潛在最大光化學量子效率(Fv/Fm)、實際光化學效率(ΦPSⅡ)、光化學淬滅系數(qp)和非光化學猝滅系數(NPQ)等參數通過以下公式計算。
Fv/Fm=(Fm-Fo)/Fm
ΦPSII= (Fm′-F′)/Fm′
NPQ=Fm/Fm′-1
qP=(Fm′-F′)/(Fm′-Fo′)
1.3.2 葉綠素含量和組織含水量的測定 于2014年7月中旬多枝檉柳幼苗葉綠素熒光參數測定的同時,采用隨機取樣法,對多枝檉柳幼苗的葉綠素和組織含水量進行測定。于10:00在Ⅰ、Ⅱ和Ⅲ3塊監測區每個調查樣方中隨機摘取若干健康的多枝檉柳幼苗葉片,分別稱取剪碎的新鮮葉片0.15 g,放入15 mL的具塞試管中,加入丙酮-乙醇混合提取液(80%丙酮與無水乙醇的體積比為1∶1),每塊樣地共有9個重復,標好記號置于避光的條件下浸提48 h,待剪碎的葉片變白,溶液呈綠色后搖勻,用紫外可見分光亮度計分別在波長663和645 nm下定OD值,計算葉綠素a和葉綠素b含量。再另取3 g新鮮葉片,浸入蒸餾水中數小時,使葉片吸水成飽和狀態,稱取葉片飽和重,再放入85 ℃干燥箱烘至恒重,稱取干重,每塊樣地共有9個重復。葉片相對含水量=(葉片鮮重-葉片干重)/(葉片飽和重-葉片干重)。
1.3.3 比根長的測定 于2014年7月中旬分別在Ⅰ、Ⅱ和Ⅲ3塊監測區每個調查樣方中隨機選取3株多枝檉柳幼苗,對其采用挖掘法挖出整株根系,而且在挖掘的過程中按照0~10、10~20、20~40、40~60、60~90、90~120、120~150 cm的土層來進行。采用沖根器將根系沖洗干凈后,用吸水紙吸干根系表面水分,并采用WINRDO根系分析系統測量不同土層的根系長度。根系經過分層測定后分別裝入標記好的信封,置于80 ℃烘箱中烘干至恒重,用電子天平稱其干重。由其根系長度與干重的比值得到不同土層根系的比根長。
1.3.4 土壤含水量的測定 土壤含水量的測定采用人工土鉆分層取樣法,分別在3塊監測區每個調查樣方中各鉆取3個土壤剖面,每個剖面采集0~10、10~20、20~40、40~60、60~90、90~120、120~150 cm土層土樣,每層3個重復,用烘干法(105 ℃)烘至恒重,稱重后計算土壤含水量。
1.4 數據處理
所得數據均使用SPSS17.0軟件進行統計分析,采用單因素方差分析(one-way ANOVA)和LSD多重比較對數據差異是否顯著(P<0.05)進行比較分析,并應用Excel軟件進行數據處理和制圖。
2 結果與分析
2.1 輸水漫溢下多枝檉柳立地土壤含水量的變化
實驗期間第1次輸水后至下次輸水到來之前(2012年10月底至2013年7月底)及第2次輸水后(2013年8月初至2014年11月),各監測區(Ⅰ~Ⅲ)0~150 cm土壤含水量均隨著時間推移總體呈持續減少趨勢;同時,Ⅰ和Ⅱ監測區土壤含水量隨著土層深度的增加而呈逐漸增加的趨勢,而Ⅲ監測區60~150 cm層含水量則呈急劇降低的趨勢(圖1)。第1次和2次輸水后,Ⅰ和Ⅱ2塊監測區各土層平均土壤含水量在2013年3月份分別為29%和28.1%,在2014年3月份分別為28.3%和30.2%,且Ⅰ、Ⅱ區之間差異不顯著(P>0.05),但兩者均顯著高于同期Ⅲ監測區的22.1%、12.3%(P<0.05)。

Ⅰ.監測區水深1 m,漫溢帶最大寬度10 m;Ⅱ. 監測區水深0.15 m,漫溢帶最大寬度3 m;Ⅲ. 監測區無水淹情況,土壤含水量為22.8%;下同圖1 不同監測區0~150 cm土壤含水量的變化(平均值±標準誤)Ⅰ. The monitoring area at a water depth of 1 m, and flood plain with maximum width 10 m; Ⅱ. The monitoring area at a water depth of 0.15 m, and flood plain with maximum width 10 m; Ⅲ. The monitoring area without flodding water, the soil water content is 22.8%; The same as belowFig.1 The change of soil water content of 0-150 cm soil depth at different monitoring areas(mean±SE)
Ⅰ和Ⅱ2塊監測區各土層平均土壤含水量在2013年7月份分別為16.9%和9.1%,分別比Ⅲ監測區高出了52.7%(P<0.05)和12%(P>0.05);它們在2014年7月份分別為19.2%和12.8%,分別比Ⅲ監測區高出了55.7%(P<0.05)和33.6%(P>0.05);而兩者在2014年11月份分別為12.9%和7.7%,分別比Ⅲ監測區高出了59.7%(P<0.05)和48.1%(P>0.05)。但同期Ⅰ和Ⅱ2塊監測區間土層平均土壤含水量始終差異不顯著(P>0.05)。
2.2 輸水漫溢下多枝檉柳生長旺季幼苗比根長的變化
在多枝檉柳生長旺季(2014年7月份),Ⅰ和Ⅱ監測區0~40 cm土層內多枝檉柳幼苗的比根長顯著高于Ⅲ監測區(P<0.05),且Ⅰ監測區幼苗的比根長又顯著高于Ⅱ監測區的(P<0.05),而Ⅲ監測區60 cm土層以下幼苗的比根長隨土層深度的增加而增大(圖2)。這表明Ⅰ和Ⅱ監測區幼苗的根系主要分布在0~40 cm的土層范圍內,而Ⅲ監測區幼苗的根系主要分布在60 cm土層以下。而由圖1可知,Ⅰ和Ⅱ監測區0~40 cm深度范圍內平均土壤含水量分別為15%和6.6%,而Ⅲ監測區60 cm土層以下的平均土壤含水量僅為2.8%,故對2014年7月份Ⅰ、Ⅱ和Ⅲ監測區幼苗生長起主要作用的平均土壤含水量分別為15%、6.6%和2.8%,其間差異顯著(P<0.05),基于此本研究主要對2014年7月份多枝檉柳幼苗的光合生理指標進行了監測。
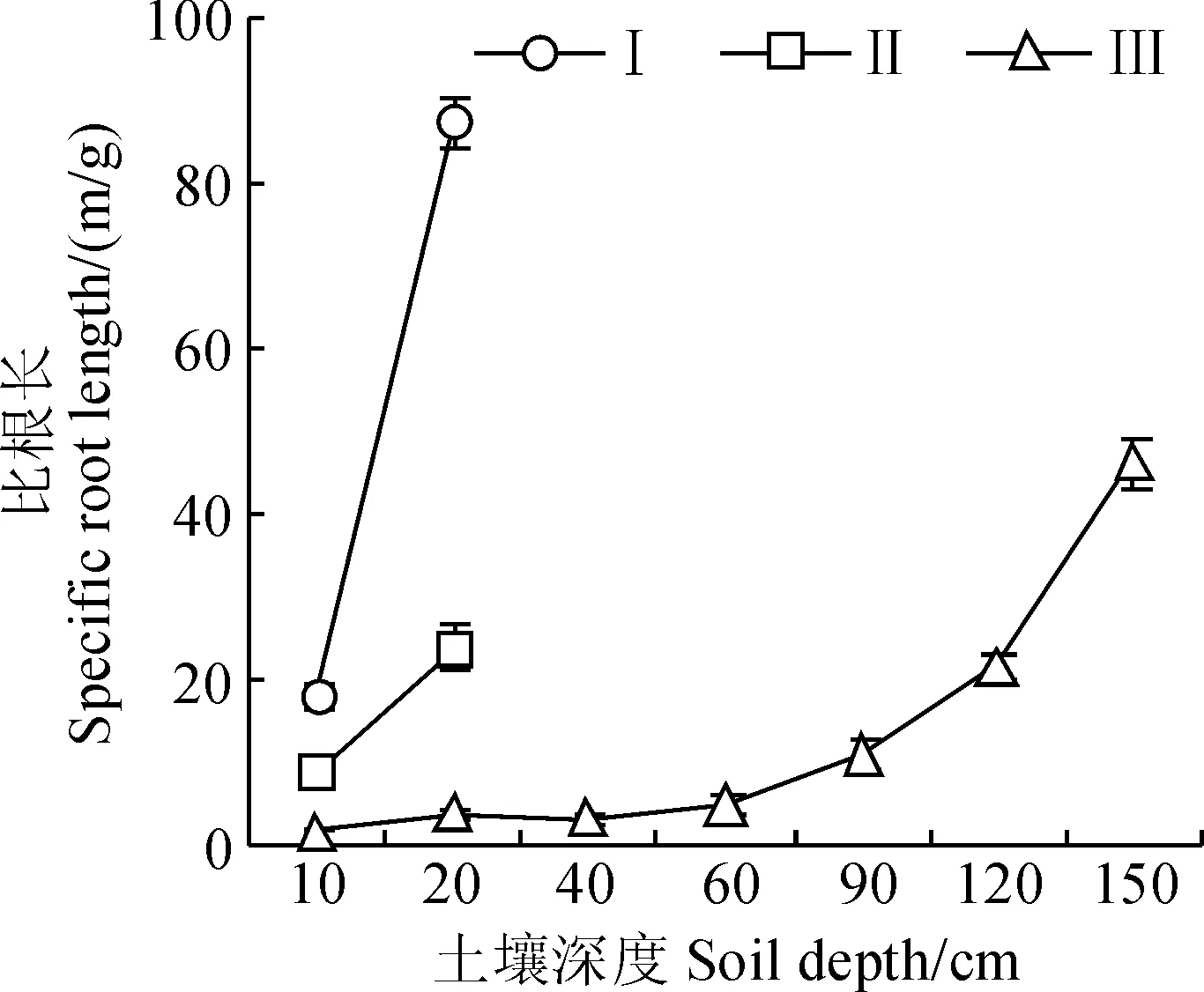
圖2 不同監測區0~150 cm多枝檉柳幼苗比根長的變化(平均值±標準誤)Fig.2 The change of specific root length of Tamarix ramosissima seedlings of 0-150 cm soil depth at different monitoring areas(mean±SE)
2.3 輸水漫溢對多枝檉柳幼苗生長及種群密度的影響
實驗期輸水漫溢后,Ⅰ、Ⅱ和Ⅲ監測區多枝檉柳幼苗種群密度在2013年7月要分別比2013年3份和2014年3月份顯著高出80.1%、78.9%、76.2%和96.9%、66.7%、72.7%(P<0.05);Ⅰ和Ⅱ監測區2014年7月份的種群密度又顯著比2014年3月份高出了96.1和54.4%(P<0.05),而Ⅲ監測區2014年7月份的種群密度卻比2014年3月份降低了3.9%(圖3)。這表明Ⅰ和Ⅱ兩監測區幼苗發生量基本呈現出春低夏高的波動態勢,而Ⅲ監測區的幼苗種群密度在第2次輸水漫溢后呈降低的趨勢。
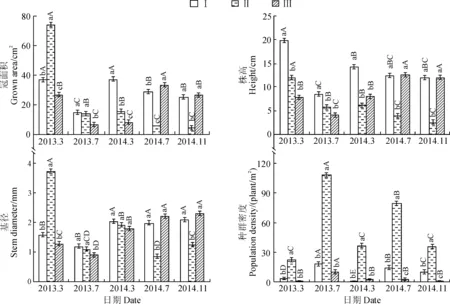
不同大寫字母表示同一監測區不同日期間在0.05水平上的顯著性差異,不同小寫字母表示同期監測區間在0.05水平存在顯著性差異圖3 不同監測區多枝檉柳幼苗種群冠面積、株高、基徑和種群密度動態變化(平均值±標準誤)Different capital letters represent significant difference between different dates of the same monitoring area at 0.05 level, while different normal letters indicate significant difference among different monitoring area within the same date at 0.05 levelFig.3 The dynamic change of crown area, height, stem diameter, population density of T. ramosissima seedlings at different monitoring areas(mean±SE)
Ⅰ、Ⅱ和Ⅲ監測區2014年7月到2014年11月幼苗種群密度下降的幅度明顯低于從2013年7月到2014年3月份的下降幅度,這表明水淹是導致幼苗種群密度降低的主要因素。
同時,2次輸水漫溢后,2013年和2014年7月份的Ⅰ和Ⅱ兩監測區多枝檉柳幼苗的冠面積、株高和基徑均比當年3月份低,且2013年7月幼苗的3個生長指標均低于次年3月的均值;而在第2次輸水漫溢后,2014年7月Ⅰ監測區生長指標冠面積、株高和基徑分別比Ⅱ監測區顯著高出80.2%、67.3%和57.4%(P<0.05)。Ⅲ監測區有所不同,2013年7月份該區幼苗種群冠面積、株高和基徑均分別低于2013年3月份和2014年3月份;但2014年7月份相應值均顯著高于2014年3月份的值(P<0.05),同時也高于Ⅰ和Ⅱ監測區2014年7月份的值(圖3)。3個生長指標比較,Ⅰ、Ⅱ和Ⅲ3個監測區由2014年7月到2014年11月份多枝檉柳幼苗冠面積和株高呈下降趨勢,而其基徑則呈上升趨勢。上述分析表明,Ⅰ和Ⅱ監測區的水過程較強,使多枝檉柳幼苗種群得到較大更新,但隨后死亡較多;Ⅲ監測區條件雖不利于幼苗種群的更新,卻使僅存的幼苗穩定生長。
2.4 輸水漫溢對多枝檉柳生長旺季幼苗葉綠素含量及葉片相對含水量的影響
葉綠素是截獲光能的主要色素,其衰減是植物在逆境條件下的一種反應,而植物組織相對水量是反映植物水分狀況的重要參數。由圖4可知,Ⅲ監測區多枝檉柳幼苗葉綠素含量最低(0.74 mg/g),分別比Ⅰ和Ⅱ監測區顯著降低48.3%和43.9%(P<0.05),而Ⅰ監測區多枝檉柳幼苗葉綠素含量雖比Ⅱ監測區的高,但差異不顯著(P>0.05);同時,從Ⅰ區、Ⅱ區到Ⅲ區,隨著輸水漫溢量的變弱,多枝檉柳幼苗葉片相對含水量呈先降低后增加的趨勢,其值分別是85.2%、81.7%和82.6%,而其間差異不顯著(P>0.05)。以上分析表明輸水漫溢量最弱的Ⅲ監測區多枝檉柳幼苗水分狀況仍處于良好的狀態,但該區水分條件已不利于多枝檉柳幼苗葉綠素含量的積累,從而可能影響到光合作用的效率。
2.5 輸水漫溢對多枝檉柳生長旺季幼苗葉綠素熒光參數的影響
自8:00開始,隨著光合有效輻射(PAR)和氣溫(T)的上升,各監測區多枝檉柳幼苗葉綠體內PSⅡ反應中心開放程度均快速降低,實際光化學量子產量(ΦPSⅡ)呈顯著下降的趨勢,下降的幅度表現為Ⅲ>Ⅱ>Ⅰ,并在12:00~16:00時降至最低,之后隨著PAR和T的下降而呈顯著上升的趨勢(圖5)。另外,整個光合作用過程中,在12:00~16:00的高光強時段,各監測區間幼苗ΦPSⅡ整體差異顯著(P<0.05),其中Ⅲ監測區幼苗ΦPSⅡ均值顯著較低;而在8:00和20:00的中低光強下,各監測區幼苗ΦPSⅡ差異不顯著(P>0.05)。這表明Ⅲ監測區多枝檉柳幼苗的PSⅡ光化學效率雖較低,但隨著PAR和T下降而得到了完全的恢復,其光合機構的光合活性在強光和高溫下只是受到了暫時的抑制。
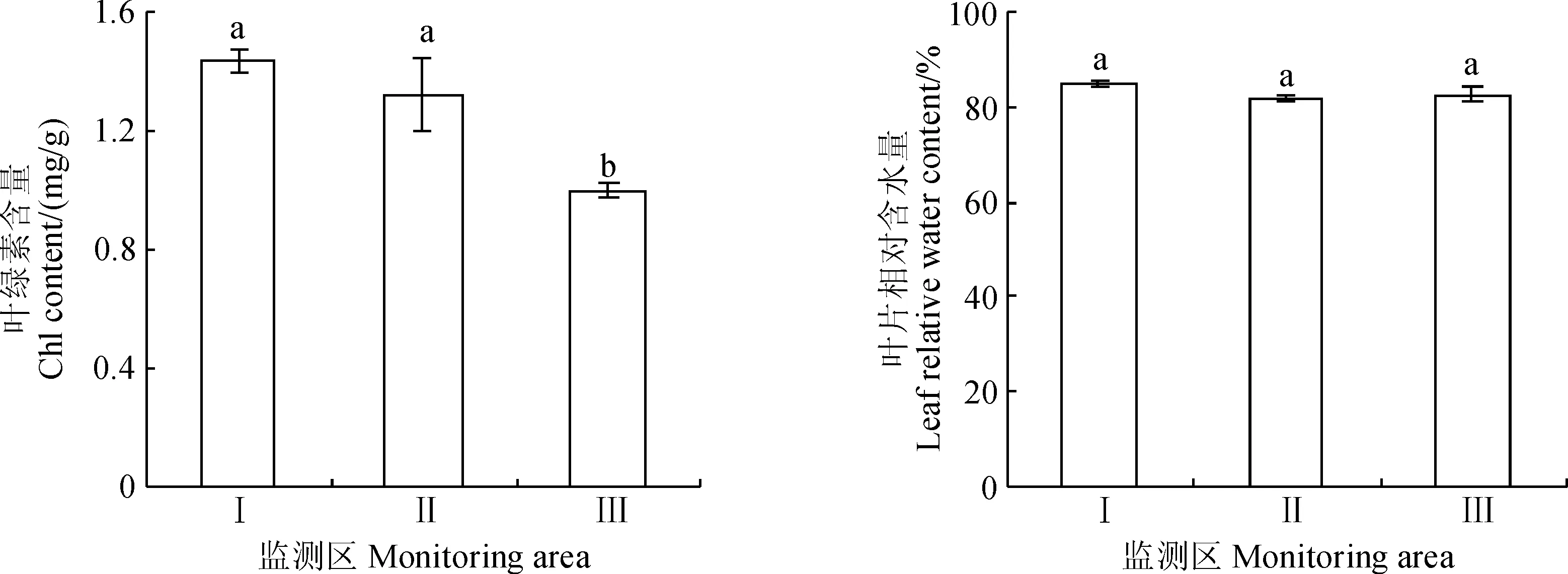
不同小寫字母表示不同監測區間0.05水平上的差異顯著;下同圖4 不同監測區多枝檉柳幼苗的葉綠素和葉片相對含水量(平均值±標準誤)Different normal letters represent significant difference between different monitoring areas at 0.05 level; The same as belowFig.4 The chlorophyll content and leaf relative water content of T. ramosissima seedlings at different monitoring areas(mean±SE)

圖5 不同監測區多枝檉柳幼苗葉綠素熒光參數日變化和PSⅡ潛在最大光化學量子效率(平均值±標準誤)Fig.5 Diurnal variation of chlorophyll fluorescence parameters and maximum photochemical efficiencies of T. ramosissima seedlings at different monitoring areas(mean±SE)
同時,各監測區多枝檉柳幼苗的光化學猝滅(qp)日變化趨勢與ΦPSⅡ較為一致(圖5),在12:00~16:00高光強時段,各監測區間幼苗的qp值差異顯著(P<0.05),并表現為Ⅰ>Ⅱ>Ⅲ,而在8:00和20:00中低光強時段,各監測區間幼苗的qp值較接近。而各監測區幼苗非光化學猝滅(NPQ)的變化趨勢與qp的相反(圖5)。隨著PAR和T的變化,各監測區幼苗NPQ呈先上升后穩定再下降的趨勢,并以Ⅲ監測區變化幅度最大。一天內,Ⅲ監測區多枝檉柳幼苗的NPQ值整體上顯著高于Ⅰ和Ⅱ監測區(P<0.05),平均增幅分別為43.6%和36.6%,且僅在12:00~16:00高光強時段,Ⅲ監測區多枝檉柳幼苗的NPQ值就分別比Ⅰ和Ⅱ監測區高出40.5%和34.8%。可見,與Ⅰ和Ⅱ監測區相比,Ⅲ監測區水分狀態雖不利于多枝檉柳幼苗PSⅡ反應中心開放部分比例的提高,但在高光照下能將葉片吸收的光能較多地分配到熱耗散,從而進行有效的光保護,體現了幼苗對高溫的適應能力。
另外,當植物處于非逆境條件時,其PSⅡ最大光化學量子效率(Fv/Fm)值一般為0.75~0.85,不受物種和生長條件的影響,而在脅迫生長條件下該值會顯著降低[13-15]。圖5顯示,各監測區多枝檉柳幼苗Fv/Fm差異均不顯著,Ⅰ和Ⅱ監測區多枝檉柳幼苗Fv/Fm值分別比Ⅲ監測區(0.759)僅高出2.6%和1.3%(P>0.05)。這表明Ⅲ監測區多枝檉柳幼苗PSⅡ光合活性雖受到一定程度的抑制,但總體仍處于較為適宜的狀態。
3 討 論
干旱區河水漫溢是河流生態系統的一個重要的環境干擾因子,河水漫溢后形成的河漫灘是河岸林實生苗補充的最佳場所,而漫溢的時間和強度以及漫溢后各種復雜土壤含水量所產生的滯后效應會對河岸林實生苗更新和光合生長產生一定的影響[16,12,4,17]。
種群密度是種群結構的基本要素,是種群數量特征的一個重要指標,通過測定密度可獲知種群的動態[18]。植物的冠面積、株高和基徑等形態特征是指示植物生長狀態對生境響應的重要指標[9,19]。本研究中,輸水漫溢后Ⅰ和Ⅱ2個監測點的多枝檉柳幼苗種群密度從3月份到當年的7月份均明顯增加,而多枝檉柳幼苗種群冠面積、株高和基徑等生長指標則均呈降低的趨勢。種群密度增加主要是因為2次輸水漫溢均給多枝檉柳種子的萌發提供了良好的水分條件,從而在河漫灘處萌生了大量的實生幼苗,這與趙振勇等研究一致[12];種群冠面積、株高和基徑等生長指標降低可能是因為在3月份的調查中,部分較大幼苗地上部分雖存在,但根系卻因長期水淹缺氧已經死亡,但仍統計在內,故使3月份的生長指標值偏高[12,20]。從2013年7月到2014年3月,Ⅰ、Ⅱ和Ⅲ3塊監測區多枝檉柳幼苗種群密度出現大幅降低趨勢,而幼苗種群冠面積、株高和基徑均表現出不同程度增加,這可能是因為輸水漫溢水淹時間從2013年8月持續到當年11月,甚至更長,導致大量較小幼苗因漫溢水淹缺少光照而大量死亡[21,22],而部分個體較大的幼苗存活下來,使幼苗種群地上形態指標偏大。此外,Ⅲ監測區多枝檉柳幼苗種群密度在2014年3月至7月則呈不顯著的降低趨勢,而種群生長指標則呈顯著增加的趨勢,這主要是因為第2次漫溢后2014年3月份該區0~10 cm表層土壤含水量僅為13.8%,從而不適合多枝檉柳種子的萌發和幼苗的定居[23],導致7月份新生幼苗占比較少,大部分幼苗穩定生長,生長指標值隨之增加。
葉綠素是植物在光合作用過程中進行光能吸收和傳遞的重要功能物質,其衰減是植物在逆境條件下的一種反應。已有研究表明葉綠素含量隨著水分脅迫的加重而減少[24],本研究結果與之一致。本研究中,Ⅲ監測區多枝檉柳幼苗葉綠素含量均明顯比Ⅰ和Ⅱ監測區低,這主要是因為Ⅰ和Ⅱ監測區多枝檉柳幼苗是新萌生的,其根系多分布在深度約40 cm的土層范圍內,且0~40 cm土壤平均含水量較高,但Ⅲ監測區多枝檉柳幼苗根系多分布在60 cm土層以下,且Ⅲ監測區60 cm以下土壤平均含水量僅為2.8%,故在水分缺乏條件下,葉綠素合成受阻而分解加快,導致葉綠素含量下降。植物葉片相對含水量在一定程度上能夠反映組織的抗脫水能力,較高的相對水分含量是植物對干旱的適應性選擇[25,26]。本研究中,隨著輸水漫溢量逐漸變弱,多枝檉柳幼苗的相對含水量呈先降低后增加的趨勢,這與楊鑫光等的研究一致[27]。這表明漫溢水分補給量最弱的Ⅲ監測區,多枝檉柳幼苗可通過調節自身葉片的水分狀況來適應干旱脅迫,表現出較強的抗脫水能力。
干旱脅迫常常會使植物的PSⅡ光能捕獲效率和電子傳遞速率發生顯著降低, 而強光合有效輻射常常會與環境脅迫相互疊加而加重彼此脅迫程度[28]。本研究中,隨著PAR的持續增加,以及輸水漫溢量的逐漸減弱,各監測區多枝檉柳幼苗ΦPSⅡ、qp、Fv/Fm呈降低的趨勢,而NPQ則呈增加的趨勢,但Fv/Fm值均處于一個相對適宜的水平(0.75~0.85),這與朱成剛等的研究結果一致[29]。這說明漫溢量較弱處多枝檉柳幼苗光能捕獲效率與光化學反應能量雖較低,但其熱耗散能力強,進而可以緩解光能過剩帶來的損傷,實現光保護,從而使其PSⅡ反應中心避免發生不可逆的損傷。
綜上所述,連續較大強度的輸水漫溢雖能夠使漫溢區多枝檉柳幼苗種群在很大程度上得到更新,但同時也導致了大量幼苗的受淹死亡,故在這種連續的輸水狀態下,多枝檉柳種群總是處于幼苗期,而不能穩定生長。因此,在實踐中建議塔里木河下游生態輸水采取強弱交替的方式進行,即在1次強輸水漫溢促進種群更新的基礎上,隨后進行1次較弱的輸水漫溢,以補充之前更新的幼苗生長所需的水分使其穩定生長,從而使幼苗種群規模持續擴大。
[1] CHEN Y N, PANG Z, CHEN Y P,etal. Response of riparian vegetation to water-table changes in the lower reaches of Tarim River, Xinjiang Uygur, China[J].HydrogeologyJournal, 2008, 16(7): 1 371-1 379.
[2] VALETT H M, BAKER M, MORRICE J A,etal. Biogeochemical and metabolic responses to the flood pulse in a semiarid floodplain[J].Ecology, 2005, 86(1): 220-234.
[3] 白 元, 徐海量, 趙新風, 等. 河水漫溢對胡楊(Populuseuphratica)分布格局的影響[J]. 中國沙漠, 2013, 33(5): 1 356-1 362.
BAI Y, XU H L, ZHAO X F,etal. Influence of river flooding on the distribution pattern ofPopuluseuphraticain the lower reaches of the Tarim River[J].JournalofDesertResearch, 2013, 33(5): 1 356-1 362.
[4] 徐海量, 葉 茂, 李吉玫, 等. 河水漫溢對荒漠河岸林植物群落生態特征的影響[J]. 生態學報, 2007, 27(12): 4 990-4 998.
XU H L, YE M, LI J M,etal. The influence of the river flooding disturbance on the native vegetation restoration in the lower reaches of Tarim River[J].ActaEcologicaSinica, 2007, 27(12): 4 990-4 998.
[5] BUSCH D E, SMITH S D. Effects of fire on water and salinity relations of riparian woody taxa[J].Oecologia, 1993, 94: 186-194.
[6] HAO X M., LI W H, HUANG X,etal. Assessment of the groundwater threshold of desert riparian forest vegetation along the middle and lower reaches of the Tarim River, China[J].HydrologicalProcesses, 2010, 24: 178-186.
[7] 蔣高明. 植物生理生態學[M]. 北京: 高等教育出版社, 2004: 137.
[8] 曾凡江, 郭海峰, 劉 波, 等. 多枝檉柳和疏葉駱駝刺幼苗生物量分配及根系分布特征[J]. 干旱區地理, 2010, 33(1): 59-64.
ZENG F J, GUO H F, LIU P,etal. Characteristics of biomass allocation and root distribution ofTamarixramosissimaLedeb. andAlhagisparsifoliaShap. seedlings[J].AridLandGeography, 2010, 33(1): 59-64.
[9] 馬曉東, 王明慧, 李衛紅, 等. 極端干旱區多枝檉柳幼苗對人工水分干擾的形態及生理響應[J]. 生態學報, 2013, 33(19): 6 081-6 087.
MA X D, WANG M H, LI W H,etal. The morphological and physiological responses ofTamarixramosissimaseedling to different irrigation methods in the extremely arid area[J].ActaEcologicaSinica, 2013, 33(19): 6 081-6 087.
[10] 單立山, 張希明, 柴仲平, 等. 多枝檉柳幼苗根系分布對灌溉量的響應[J]. 干旱區研究, 2007, 24(2): 213-218.
SHAN L S, ZHANG X M, CHAI Z P,etal. Response of the distribution of roots ofTamarixramosissimaseedlings to irrigation with different volumes[J].AridZoneResearch, 2007, 24(2): 213-218.
[11] 趙峰俠, 尹林克. 荒漠內陸河岸胡楊和多枝檉柳幼苗種群空間分布格局及種間關聯性[J]. 生態學雜志, 2007, 26(7): 972-977.
ZHAO F X, YIN L K. Spatial distribution pattern and interspecific association ofPopuluseuphraticaandTamarixramosissimaseedlings populations along desert inland river[J].ChineseJournalofEcology, 2007, 26(7): 972-977.
[12] 趙振勇, 張 科, 盧 磊, 等. 塔里木河中游洪水漫溢區荒漠河岸林實生苗更新[J]. 生態學報, 2011, 31(12): 3 322-3 329.
ZHAO Z Y, ZHANG K, LU L,etal. Seedling recruitment in desert riparian forest following river flooding in the middle reaches of the Tarim River[J].ActaEcologicaSinica, 2011, 31(12): 3 322-3 329.
[13] 張守仁. 葉綠素熒光動力學參數的意義及討論[J]. 植物學通報, 1999, 16(4): 444-448.
ZHANG S R. A discussion on chlorophyll fluorescence kinetics parameters and their significance[J].ChineseBulletinofBotany, 1999, 16(4): 444-448.
[14] 何炎紅, 郭連生, 田有亮. 白刺葉不同水分狀況下光合速率及其葉綠素熒光特性的研究[J]. 西北植物學報, 2005, 25(11): 88-95.
HE Y H, GUO L S, TIAN Y L. Photosynthetic rates and chlorophyll fluorescence ofNitrariatangutorumat different leaf water potentials[J].ActaBotanicaBoreali-occidentaliaSinica, 2005, 25(11): 88-95.
[15] KRAMER D M, JOHNSON G, KIIRATS O,etal. New fluorescence parameters for the determination of QA redox state and excitation energy fluxes[J].PhotosynthesisResearch, 2004, 79(2): 209-218.
[16] VALETT H M, BAKER M, MORRICE J A,etal. Biogeochemical and metabolic responses to the flood pulse in a semiarid floodplain[J].Ecology, 2005, 86(1): 220-234.
[17] 王 鵬, 趙成義, 李 君. 地下水埋深及礦化度對多枝檉柳幼苗光合特征及生長的影響[J]. 水土保持通報, 2012, 32(2): 84-89.
WANG P, ZHAO C Y, LI J. Effects of groundwater depth and mineralization degree on photosynthesis and growth ofTamarixramosissimaseedlings[J].BulletinofSoilandWaterConservation, 2012, 32(2): 84-89.
[18] 倪紅偉, 高玉慧, 高亦珂, 等 三江平原不同群落類型小葉章種群密度季節動態比較[J]. 東北林業大學學報, 1999, 20(1): 40-42.
NI H W, GAO Y H, GAO Y K,etal. Dynamic comparison of population density between differentDeyeuxiaangustisfoliacommunities in Sanjiang Plain[J].JournalofNortheastForestryUniversity, 1999, 20(1): 40-42.
[19] 魏 疆, 張希明, 馬文東, 等. 甘蒙檉柳幼苗生長動態及其對沙漠腹地生境條件的適應策略[J]. 干旱區地理, 2007, 30(5): 666-673.
WEI J, ZHANG X M, MA W D,etal. Seedling growth dynamics ofTamarixaustromongolicaand its acclimation strategy in hinterland of desert[J].AridLandGeography, 2007, 30(5): 666-673.
[20] 趙可夫. 植物對水澇脅迫的適應[J]. 生物學通報, 2003, 38(12): 11-14.
ZHAO K F. Plants adaptation to waterlogging stress[J].BulletinofBiology, 2003, 38(12): 11-14.
[21] 陳家寬, 陳中義. 不同生境內瀕危植物長喙毛茛澤瀉種群數量動態比較[J]. 植物生態學報, 1999, 23(1): 9-14.
CHEN J K, CHEN ZH Y. Population dynamics ofRanalismarostratum, an endangered species growing in different habitats[J].ChineseJournalofPlantEcology, 1999, 23(1): 9-14.
[22] 曹秋玉. 北固山濕地蘆葦和虉草種群的生長規律及其生態功能的研究[D]. 江蘇鎮江:江蘇大學, 2006.
[23] 徐海量, 葉 茂, 李吉枚, 等. 不同水分供應對塔里木河下游土壤種子庫種子萌發的影響[J]. 干旱區地理, 2008, 31(5): 650-658.
XU H L, YE M, LI J M,etal. Effect of different water treatments on the germination of soil seed bank at lower reaches of Tarim River[J].AridLandGeography, 2008, 31(5): 650-658.
[24] 孫 龍, 彭祚登, 王 沖, 等. 干旱脅迫對兩種沙地灌木能源樹種光合特性的影響[J]. 南京林業大學學報(自然科學版), 2014, 38(2): 99-104.
SUN L, PENG Z D, WANG C,etal. Effect of drought stress on photosynthetic characteristics of two energy resource sandy shrubs[J].JournalofNanjingForestryUniversity, 2014, 38(2): 99-104.
[25] 李雪華, 蔣德明, 阿拉木薩, 等. 科爾沁沙地4種植物抗旱性的比較研究[J]. 應用生態學報, 2002, 13(11): 1 385-1 388.
LI X H, JIANG D M, ALAMUSA,etal. A comparative study on drought-resistance of four plant species in Kerqin sandy land[J].ChineseJournalofAppliedEcology, 2002, 13(11): 1 385-1 388.
[26] 李向義, THOMAS F M, FOETZKI A, 等. 自然狀況下頭狀沙拐棗對水分條件變化的響應[J]. 植物生態學報, 2003, 27(4): 516-521.
LI X Y, THOMAS F M, FOETZKI A,etal. The responses ofCalligonumcaput-medusaeto changes of water conditions under natural environment[J].ChineseJournalofPlantEcology, 2003, 27(4): 516-521.
[27] 楊鑫光, 傅 華, 李曉東. 干旱脅迫對霸王水分生理特征及細胞膜透性的影響[J]. 西北植物學報, 2009, 29(10): 2 076-2 083.
YANG X G, FU H, LI X D. Water physiological characteristics and membrane permeability in the leaves ofZygophyllumxanthoxylumunder drought stress[J].ActaBotanicaBoreali-occidentaliaSinica, 2009, 29(10): 2 076-2 083.
[28] REMORINI D, MELGAR J C, GUIDI L,etal. Interaction effects of root-zone salinity and solar irradiance on the physiology and biochemistry of Oleaeuropaea[J].EnvironmentalandExperimentalBotany, 2009, 65(2/3): 210-219.
[29] 朱成剛, 李衛紅, 馬建新, 等. 塔里木河下游地下水位對檉柳葉綠素熒光特性的影響[J]. 應用生態學報, 2010, 21(7): 1 689-1 696.
ZHU CH G, LI W H, MA J X,etal. Effects of groundwater level on chlorophyll fluorescence characteristics ofTamarixhispidain lower reaches of Tarim River[J].ChineseJournalofAppliedEcology, 2010, 21(7): 1 689-1 696.
(編輯:裴阿衛)
Response of Growth Dynamic and Photosynthetic Characteristics of Tamarix ramosissima Seedlings to River Flooding in the Lower Reaches of Tarim River
WANG Minghui, MA Xiaodong*, ZHANG Ruiqun, XING Xuming
(Key Laboratory of Species Conservation and Control in Xinjiang, College of Life Sciences, Xinjiang Normal University, Urumqi 830054, China)
TakingTamarixramosissimaseedlings in the natural flooding area along lower reaches of Tarim River as objects, we analyzed the growth of seedlings and variation of the population under continuous flooding disturbance, and the photosynthetic dynamic responses of seedlings under different flood intensities. Conclusions were as following: (1) the population density ofT.ramosissimaseedlings under continuous twice strong flooding disturbance, from March 2013 to July 2014, increased firstly, then decreased and increased lastly, but the seedlings’ crown, height and stem diameter decreased firstly, then increased and decreased. (2) The population density of the seedlings increased firstly and then decreased in monitoring area Ⅲ in which the second flooding disturbance was weaker, but its crown, height and stem diameter decreased firstly and then increased. (3) The leaf relative water content of the seedlings decreased firstly with the second flooding disturbance decreasing and then increased. At the same time, the chlorophyll content, actual photochemical efficiency of photosystem Ⅱ in the light (ΦPSⅡ), photochemical quenching (qP) of the seedlings decreased, the non-photochemical quenching (NPQ) increased, but the maximal photochemical efficiencies of PSⅡ (Fv/Fm) maintained optimum value. The research means that although the continuous twice strong flooding disturbance could make the population ofT.ramosissimaseedlings regenerated, it also cause death to a large number of seedlings for flooding. As a result, the population ofT.ramosissimaseedlings could not form a certain scale. We suggest that we can adopt the strong and weak alternately manner to conduct ecological water conveyance, which can improve the survival rate of seedlings, and make the seedlings to grow stably.
TamarixramosissimaL.; flooding disturbance; the population; the growth of seedlings; the chlorophyll fluorescence parameters
1000-4025(2016)10-2038-09
10.7606/j.issn.1000-4025.2016.10.2038
2016-04-04;修改稿收到日期:2016-10-23
國家自然科學基金(41261103);國家科技支撐計劃(2014BAC15B02);新疆師范大學研究生科技創新基金(20131208);國家自然科學基金NSFC-新疆聯合基金(U-1303102)
王明慧(1991-),女,在讀碩士研究生,主要從事植物生態方面的研究。E-mail:907009956@qq.com
*通信作者:馬曉東,博士,碩士生導師,主要從事植物生態方面的研究。E-mail:mxd1107@126.com
Q945.79
A
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
