海南省2個(gè)冬青葉馬尾藻種群的多樣性及分子系統(tǒng)學(xué)研究
2016-12-17 08:27:32鄒瀟瀟劉鑫鑫黃惠琴鮑時(shí)翔
廣東海洋大學(xué)學(xué)報(bào) 2016年6期
鄒瀟瀟,劉鑫鑫,朱 軍,林 勇,黃惠琴,鮑時(shí)翔
(中國(guó)熱帶農(nóng)業(yè)科學(xué)院熱帶生物技術(shù)研究所,海南 海口 571101)
海南省2個(gè)冬青葉馬尾藻種群的多樣性及分子系統(tǒng)學(xué)研究
鄒瀟瀟,劉鑫鑫,朱 軍,林 勇,黃惠琴,鮑時(shí)翔
(中國(guó)熱帶農(nóng)業(yè)科學(xué)院熱帶生物技術(shù)研究所,海南 海口 571101)
為了解海南省冬青葉馬尾藻不同地理種群的多樣性,從潭門鎮(zhèn)和長(zhǎng)坡鎮(zhèn)收集到12份冬青葉馬尾藻樣品,通過PCR擴(kuò)增和測(cè)序獲得了其cox1(細(xì)胞色素C氧化酶亞基1)、rbcL(1,5-二磷酸核酮糖羧化酶/加氧酶大亞基)及ⅠTS(內(nèi)轉(zhuǎn)錄間隔區(qū))序列。多序列比對(duì)顯示,2個(gè)種群的冬青葉馬尾藻的cox1、rbcL序列很保守,未發(fā)現(xiàn)多態(tài)性位點(diǎn);而ⅠTS序列變異較大,在堿基組成和序列長(zhǎng)度上均有豐富的多態(tài)性。根據(jù)ⅠTS的序列特征將所分析樣品分為A、B和C 3個(gè)類群。系統(tǒng)進(jìn)化分析顯示,3個(gè)類群能以較高的置信度與數(shù)據(jù)庫(kù)中其他地域來源的冬青葉馬尾藻聚為一支,其中A、B類群的親緣關(guān)系非常近,而C類群與其他地域來源的冬青葉馬尾藻更相似。
褐藻;冬青葉馬尾藻;種群;多樣性;系統(tǒng)進(jìn)化
馬尾藻(Sargassum spp.)廣泛分布于熱帶和亞熱帶海域的近岸低潮帶和潮下帶[1-2],是褐藻門(Phaeophyta)墨角藻目(Fucales)中一類重要經(jīng)濟(jì)海藻[3],在工業(yè)、農(nóng)業(yè)、食品、醫(yī)藥及生態(tài)修復(fù)等方面都具有廣闊的開發(fā)與應(yīng)用前景[4-9]。
馬尾藻屬是墨角藻目中形態(tài)和分類最為復(fù)雜的屬之一[3],屬以下可再細(xì)分為亞屬、組、亞組、系等分類階元。傳統(tǒng)的馬尾藻分類主要依據(jù)其外部的形態(tài)特征,如葉、氣囊、生殖托以及初生分枝、次生分枝等進(jìn)行區(qū)分[10-11],然而由于馬尾藻的形態(tài)易隨時(shí)間、環(huán)境等因素的影響而變化,甚至有些種類在個(gè)體間的變異很大,導(dǎo)致馬尾藻屬的物種分類非常困難。迄今為止,馬尾藻屬有記錄的物種(包括同種異名)已有近1 000個(gè),但是被證實(shí)的物種僅353個(gè)[12]。
冬青葉馬尾藻(S.ilicifolium)是馬尾藻屬分布較為廣泛的物種之一,印度尼西亞、爪哇、馬來西亞、澳大利亞及我國(guó)臺(tái)灣、海南等地均有報(bào)道[1]。冬青葉馬尾藻能有效吸附重金屬元素[13-14],并且其活性提取物還具有良好的鎮(zhèn)痛消炎作用和抗氧化、抗腫瘤等作用[15-16],因此在生態(tài)修復(fù)和醫(yī)藥等領(lǐng)域都有很大的應(yīng)用潛力。
在傳統(tǒng)的分類中,冬青葉馬尾藻隸屬于真馬尾藻亞屬刺托組冬青葉系(S.sect.Acanthocarpicae,ser.Ilicifolia)[1,17]。隨著DNA條形碼技術(shù)在馬尾藻分類鑒定中的應(yīng)用,研究者發(fā)現(xiàn)刺托組物種具有明顯的多系性,因此建議對(duì)該組內(nèi)物種形態(tài)鑒定的主要依據(jù)進(jìn)行修訂,并對(duì)刺托組以下的分類階元進(jìn)行重新劃分[17-18]。Mattio等[17]結(jié)合形態(tài)鑒定及分子系統(tǒng)學(xué)研究結(jié)果,提出了針對(duì)刺托組的修訂方案,其中包括將冬青葉系(S.ser.Ilicifoliae)提升為冬青葉組(S.sect.Ilicifoliae)的建議。
海南的馬尾藻資源非常豐富,冬青葉馬尾藻是其中較為常見的一種。由于對(duì)其資源分布規(guī)律和遺傳多樣性的認(rèn)識(shí)十分有限,目前海南的馬尾藻資源并沒有得到很好的應(yīng)用。為研究海南省冬青葉馬尾藻不同地理種群的多樣性,筆者從長(zhǎng)坡鎮(zhèn)和潭門鎮(zhèn)采集了冬青葉馬尾藻樣品,測(cè)定其cox1、rbcL及ⅠTS的序列,并對(duì)其種群的多樣性和系統(tǒng)進(jìn)化關(guān)系進(jìn)行分析,以期豐富對(duì)海南省冬青葉馬尾藻遺傳多樣性的認(rèn)識(shí),以及為進(jìn)一步研究冬青葉馬尾藻的系統(tǒng)進(jìn)化提供基礎(chǔ)。
1 材料與方法
1.1 樣品采集及處理
本研究的馬尾藻實(shí)驗(yàn)材料,TM1~TM6采集于海南省潭門鎮(zhèn),CP1~CP6采集于長(zhǎng)坡鎮(zhèn)。樣品采集時(shí),選擇成熟、完整的藻體,連同固著器將整個(gè)藻株一起鏟下。用海水反復(fù)沖洗,去除表面附著的泥沙、雜藻及多毛類、甲殼類等動(dòng)物后,帶回實(shí)驗(yàn)室暫養(yǎng),以便形態(tài)鑒定及DNA提取。
1.2 馬尾藻基因組DNA提取
將馬尾藻的藻體清洗干凈,選取約1.0 g新鮮幼嫩的藻葉,吸干表面水分后,用液氮充分研磨。根據(jù)改良的 CTAB 法提取馬尾藻基因組DNA[19]。
1.3 PCR擴(kuò)增與序列測(cè)定
將馬尾藻基因組DNA的濃度稀釋至50ng/μL,以作為PCR反應(yīng)的模板。cox1、rbcL及ⅠTS的擴(kuò)增引物如表1。

表1 引物信息Table 1 Primers used in this study
用10mg/mL瓊脂糖凝膠電泳對(duì)PCR反應(yīng)產(chǎn)物進(jìn)行檢測(cè),并利用SanPrep柱式DNA膠回收試劑盒(生工生物工程上海股份有限公司)進(jìn)行回收。回收產(chǎn)物與pMD18-T載體(大連寶生物有限公司)連接后,轉(zhuǎn)化至大腸桿菌感受態(tài)細(xì)胞。隨機(jī)挑選3-5個(gè)陽(yáng)性克隆,送至生工生物工程(上海)股份有限公司進(jìn)行測(cè)序。
1.4 數(shù)據(jù)分析
利用chromas軟件檢查測(cè)序結(jié)果的峰圖,然后用DNAman軟件對(duì)序列進(jìn)行校正和拼接,以獲得各基因片段的一致序列。另外,從GenBank數(shù)據(jù)庫(kù)中下載馬尾藻屬部分物種的相關(guān)序列、并以喇叭藻屬(Turbinaria)的T.ornata作為外群(表2),采用最大似然法(Maximum Likelihood)構(gòu)建系統(tǒng)進(jìn)化樹(Bootstrap設(shè)為1 000次重復(fù)),然后基于Kimura 2-parameter模型計(jì)算種間的遺傳距離。

表2 GenBank下載的馬尾藻物種的序列Table 2 GenBank accession numbers of Sargassum spp.used in the present study
2 結(jié)果與分析
2.1 冬青葉馬尾藻的cox1、rbcL及ⅠTS的序列多態(tài)性
通過PCR擴(kuò)增和測(cè)序,得到12份冬青葉馬尾藻樣品的cox1、rbcL及ⅠTS序列。其中cox1擴(kuò)增產(chǎn)物為561 bp,rbcL擴(kuò)增產(chǎn)物為488 bp;ⅠTS序列為1 498~1 491 bp。通過多序列比對(duì)發(fā)現(xiàn)12份冬青葉馬尾藻樣品的cox1和rbcL均非常保守,沒有找到多態(tài)性位點(diǎn),并且分別與數(shù)據(jù)庫(kù)中冬青葉馬尾藻的cox1(GenBank序列登錄號(hào):HQ416039)和rbcL(GenBank序列登錄號(hào):KF282003)的序列相似性分別達(dá)99.9%和100%。而ⅠTS序列的變異相對(duì)較大,在序列長(zhǎng)度和堿基組成上均發(fā)現(xiàn)了較為豐富的多態(tài)性。由于目前未見冬青葉馬尾藻ⅠTS序列全長(zhǎng)的報(bào)道,因此對(duì)獲得的ⅠTS序列進(jìn)行了重點(diǎn)分析。
2.2 冬青葉馬尾藻的ⅠTS序列特征
12份冬青葉馬尾藻樣品的ⅠTS序列可以分為3類(GenBank序列登錄號(hào):KU857032~KU857034),分別標(biāo)記為ⅠTS-A、ⅠTS-B和ⅠTS-C(表3)。通過與GenBank中其他馬尾藻物種的ⅠTS序列進(jìn)行比對(duì)后,確定了ⅠTS1和ⅠTS2的長(zhǎng)度范圍。
12條冬青葉馬尾藻ⅠTS序列的GC含量基本一致,其中ⅠTS1和ⅠTS2的GC含量分別為 57.4%~57.5% 和55.9%~56.3%。ⅠTS-A全長(zhǎng)為1 489 bp,包括采自長(zhǎng)坡鎮(zhèn)的5份和潭門鎮(zhèn)的3份冬青葉馬尾藻;ⅠTS-B的序列全長(zhǎng)為1 490 bp,包括采自潭門鎮(zhèn)的另外4份冬青葉馬尾藻;此外,長(zhǎng)坡鎮(zhèn)和潭門鎮(zhèn)還各有1份冬青葉馬尾藻樣品的ⅠTS序列全長(zhǎng)為1 488 bp,標(biāo)記為ⅠTS-C(表1)。與ⅠTS-A相比,ⅠTS-B存在1處堿基顛換(C-A),以及1處單堿基插入(1468位點(diǎn)發(fā)生C的插入);相對(duì)于前兩種ⅠTS序列,ⅠTS-C的變異較大,共發(fā)生1處單堿基丟失和6處堿基變異(圖1)。發(fā)生變異位點(diǎn)多分布在ⅠTS1區(qū)(66.7%),而5.8s 極為保守,未出現(xiàn)變異位點(diǎn)。根據(jù)ⅠTS序列的種類將本研究的冬青葉馬尾藻樣品分成A、B和C 3個(gè)不同的類群,以便于后續(xù)的系統(tǒng)進(jìn)化分析。

表3 不同類群的冬青葉馬尾藻的ⅠTS序列特征及樣品來源信息Table 3 Features ofⅠTS sequences and sample information of different S.ilicifolium types
2.3 系統(tǒng)進(jìn)化分析
目前 GenBank數(shù)據(jù)庫(kù)中馬尾藻屬物種的序列信息十分有限,僅7個(gè)物種同時(shí)檢索到了ⅠTS2、cox1和rbcL 3個(gè)片段的序列信息。由于ⅠTS2序列是馬尾藻屬的系統(tǒng)進(jìn)化分析中最常用的標(biāo)記之一,有序列信息的物種相對(duì)豐富,且大多具有憑證標(biāo)本作為驗(yàn)證,增加了數(shù)據(jù)的可靠性。因此本研究分別采用3個(gè)基因片段聯(lián)合分析和ⅠTS2序列單獨(dú)分析兩種方法,與數(shù)據(jù)庫(kù)中的參考序列進(jìn)行了比對(duì),并以喇叭藻屬(Turbinaria)作為外群,采用最大似然法構(gòu)建系統(tǒng)進(jìn)化樹(圖2—3)。結(jié)果顯示,兩種方法構(gòu)建的系統(tǒng)進(jìn)化樹中,不同的冬青葉馬尾藻均能優(yōu)先聚為一支,其中A、B類群首先聚在一起,說明它們之間的親緣關(guān)系更近;另外,葉囊馬尾藻(S.phyllocystum)也顯示出與冬青葉馬尾藻之間較近的親緣關(guān)系。不同的是,在3個(gè)片段聯(lián)合構(gòu)建的系統(tǒng)進(jìn)化樹中,匍枝馬尾藻(S.polycystum) 與冬青葉馬尾藻之間的親緣關(guān)系很遠(yuǎn),而在以ⅠTS2聯(lián)合構(gòu)建的系統(tǒng)進(jìn)化樹中,兩者的親緣關(guān)系較近。

圖1 三種冬青葉馬尾藻的ⅠTS序列比對(duì)Fig.1 Alignment of three types of S.ilicifoliumⅠTS fragments
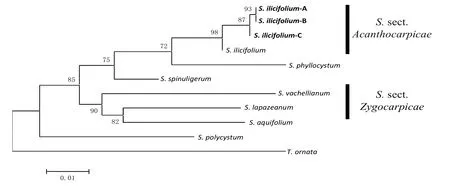
圖2 基于ⅠTS2-cox1-rbcL序列的系統(tǒng)進(jìn)化分析Fig.2 Phylogenetic analysis based onⅠTS2-cox1-rbcL sequences

圖3 基于ⅠTS2序列的系統(tǒng)進(jìn)化分析Fig.3 Phylogenetic analysis based onⅠTS2 sequences
基于Kimura 2-parameter模型計(jì)算各馬尾藻的種間遺傳距離,結(jié)果顯示,利用ⅠTS2計(jì)算的遺傳距離比利用cox1、rbcL及ⅠTS2這3個(gè)片段計(jì)算的平均遺傳距離略大(表 3)。海南的冬青葉馬尾藻3個(gè)類群中,A、B類群的之間的遺傳距離最小,平均遺傳距離為0.000,它們與C類群的平均遺傳距離為0.003;而C類群與數(shù)據(jù)庫(kù)中冬青葉馬尾藻親緣關(guān)系較近,平均遺傳距離為 0.003。與所分析的其他馬尾藻屬物種相比,冬青葉馬尾藻與葉囊馬尾藻的平均遺傳距離較近,為0.019~0.031;與瓦氏馬尾藻(S.vachellianum)的平均遺傳距離相對(duì)較遠(yuǎn),為0.054~0.078(表4)。

表4 不同馬尾藻的種間遺傳距離Table 4 Pairwise genetic distances between different Sargassum species
3 討論
3.1 不同冬青葉馬尾藻地理種群的多樣性
ⅠTS、cox1和rbcL等是藻類分子系統(tǒng)學(xué)研究中應(yīng)用較多的基因片段。其中ⅠTS因進(jìn)化速度相對(duì)較快,已廣泛用于大型藻類近緣種、甚至同物種不同種群間的系統(tǒng)進(jìn)化分析[23],并且在馬尾藻屬物種的研究中也逐漸應(yīng)用起來[24-27];此外,分別位于線粒體和葉綠體的cox1和rbcL基因,因具有引物通用性強(qiáng)且變異速率適宜的優(yōu)點(diǎn),也被認(rèn)為是較為理想的DNA條形碼標(biāo)記[28-29]。本研究利用這3個(gè)片段對(duì)海南省2個(gè)地理種群的冬青葉馬尾藻的遺傳多樣性進(jìn)行了初步分析。其中cox1和rbcL未能找到變異位點(diǎn),而ⅠTS序列在長(zhǎng)度和堿基組成上均發(fā)現(xiàn)了較為豐富的多態(tài)性。其原因可能是cox1和rbcL分別在呼吸作用和光合作用中起著重要的作用,承受的選擇壓力比非編碼的ⅠTS略大,因而在種內(nèi)的分辨率不夠高。
本研究根據(jù)ⅠTS序列將2個(gè)地理種群的冬青葉馬尾藻樣品分為3個(gè)類群。系統(tǒng)進(jìn)化分析顯示,與C類群相比,A、B類群間的親緣關(guān)系更近;而C類群與數(shù)據(jù)庫(kù)中來源于南太平洋新喀里多尼亞的冬青葉馬尾藻(憑證標(biāo)本編號(hào);ⅠRD1617)[17]更相似。但是,由于目前具有來源和憑證標(biāo)本信息的馬尾藻的數(shù)據(jù)非常有限,筆者暫時(shí)無(wú)法對(duì)海南的冬青葉馬尾藻的與其他地域來源的樣品之間的關(guān)系進(jìn)行進(jìn)一步研究。
3.2 冬青葉馬尾藻的分類地位
在墨角藻目中,馬尾藻屬是物種最為豐富,分類最為復(fù)雜的屬之一。目前,馬尾藻屬被分為真馬尾藻亞屬(S.subgen.Sargassum)、反曲葉亞屬(S.subgen.Bactrophycus)、節(jié)葉亞屬( S.subgen.Arthrophycus)和葉枝亞屬(S.subgen.Phyllotrichia)4個(gè)亞屬,其中以真馬尾藻亞屬的物種最為豐富多樣[10]。在傳統(tǒng)的分類中,冬青葉馬尾藻是真馬尾藻亞屬刺托組冬青葉系的代表性物種。本研究利用兩種方法構(gòu)建的系統(tǒng)進(jìn)化樹中,不同地域來源的冬青葉馬尾藻物種都能首先聚在真馬尾藻亞屬的一個(gè)小分支內(nèi),這與形態(tài)學(xué)分類的結(jié)果一致,也暗示這些冬青葉馬尾藻的起源可能具有單系性。
此外,本研究中利用兩種方法構(gòu)建的系統(tǒng)進(jìn)化樹顯示,傳統(tǒng)分類上同屬于刺托組的硬葉馬尾藻和冬青葉馬尾藻均位于不同的分支,印證了刺托組物種為多系發(fā)生的觀點(diǎn)[17,18]。由于具有ⅠTS2序列的物種信息相對(duì)豐富,從本研究利用ⅠTS2序列構(gòu)建的系統(tǒng)進(jìn)化樹可以看出,硬葉馬尾藻與瓦氏馬尾藻在同一個(gè)分支,且此分支與冬青葉馬尾藻所在的分支處于并列的地位,因此進(jìn)一步支持了Mattio等[17]提出的將冬青葉系提升為冬青葉組(S.sect.Ilicifoliae),以及將原來屬于刺托組的硬葉馬尾藻劃分到新組S.sect.Binderianae的建議。
3.3 馬尾藻傳統(tǒng)分類和分子分類中存在的問題
本研究中利用3個(gè)基因片段和ⅠTS2分別構(gòu)建的系統(tǒng)進(jìn)化樹上,除了匍枝馬尾藻所處的位置變化較大之外,其他物種的聚類結(jié)果基本一致,但是利用3個(gè)基因片段構(gòu)建的系統(tǒng)進(jìn)化樹的支持度比ⅠTS2序列單獨(dú)構(gòu)建的略高,可見多基因片段聯(lián)合分析有利于提高分析結(jié)果的準(zhǔn)確性。另外,利用ⅠTS2序列的系統(tǒng)進(jìn)化分析顯示山德馬尾藻(S.sandei)與冬青葉馬尾藻聚在一個(gè)小分支內(nèi),在傳統(tǒng)分類中,山德馬尾藻和冬青葉馬尾藻同屬于冬青葉系,該結(jié)果證實(shí)了它們之間的親緣關(guān)系非常近,但也說明依靠單一標(biāo)記對(duì)馬尾藻近緣物種進(jìn)行分辨時(shí)具有一定局限性。
此外,在ⅠTS2序列構(gòu)建的系統(tǒng)進(jìn)化樹上,冬青葉馬尾藻重緣變種(S.ilicifolium var.conduplicatum)沒有與冬青葉系的物種聚在一起,而與硬葉馬尾藻(S.aquifolium)和斯氏馬尾藻(S.swartzii)顯示出較近的親緣關(guān)系,這與傳統(tǒng)分類不一致。但由于數(shù)據(jù)庫(kù)中沒有冬青葉馬尾藻重緣變種的其他基因序列信息,筆者無(wú)法對(duì)其與冬青葉馬尾藻的近緣關(guān)系做進(jìn)一步分析。
由于馬尾藻物種的形態(tài)變異很大,依據(jù)其形態(tài)特征的分類系統(tǒng)中一直存在很多有爭(zhēng)議的問題,因而出現(xiàn)分子分類和傳統(tǒng)分類不一致的現(xiàn)象也比較常見。隨著分子系統(tǒng)學(xué)研究在馬尾藻分類學(xué)中越來越多地應(yīng)用,傳統(tǒng)的馬尾藻屬分類系統(tǒng)發(fā)生了一些重大改變,如對(duì)先前的一些分類標(biāo)準(zhǔn)進(jìn)行重新評(píng)估,以及去除一些不合理的分類方法等[17-18,30]。但是,由于目前還沒有確定適合于馬尾藻分子系統(tǒng)學(xué)研究的分子標(biāo)記,馬尾藻物種的準(zhǔn)確分類和其系統(tǒng)演化關(guān)系的明確,還需要進(jìn)行大量的工作。
[1]曾呈奎.中國(guó)海藻志 [M].北京:科學(xué)出版社,2000.
[2]曾呈奎.中國(guó)黃渤海海藻 [M].北京:科學(xué)出版社,2009.
[3]PHⅠLLⅠPS N.Biogeography of Sargassum (Phaeophyta) in the Pacific basin[M]//ABBOTTⅠ A.Taxonomy of economic seaweeds with reference to some Pacific species,vol.5.La Jolla,California:California Sea Grant College,1995:107-144.
[4]文竹青,高偉,何少華.藻類吸附法去除廢水中的重金屬 [J].江蘇環(huán)境科技,2006,19(2):49-51.
[5]De SOUSA A P A,TORRES M R,PESSOA C,et al.Ⅰn vivo growth-inhibition of Sarcoma 180 tumor by alginates from brown seaweed Sargassum vulgare [J].Carbohydrate Polymers,2007,69(1):7-13.
[6]羅先群,王新廣,楊振斌.馬尾藻的營(yíng)養(yǎng)成分測(cè)定及多糖的提取 [J].化學(xué)與生物工程,2007,24(4):64-66.
[7]張永亮,張浩江,謝水波,等.藻類吸附重金屬的研究進(jìn)展 [J].鈾礦冶,2009,28(1):31-37.
[8]王博,黃和,熊皓平,等.硇洲馬尾藻的營(yíng)養(yǎng)成分分析與評(píng)價(jià) [J].食品研究與開發(fā),2010,31(10):195-197.
[9]ERMAKOⅤA S,SOKOLOⅤA R,KⅠM S,et al.Fucoidans from brown seaweeds Sargassum hornery,Eclonia cava,Costaria costata:Structural characteristics and anticancer activity [J].Applied Biochemistry & Biotechnology,2011,164(6):841-850.
[10]MATTⅠO L,PAYRⅠ C E,STⅠGER-POUⅤREAU Ⅴ.Taxonomic revision of Sargassum( Fucales,Phaeophyceae) from French Polynesia based on morphological and molecular analyses [J].Journal of Phycology,2008,44(6):1541–1555.
[11]YOSHⅠDA T.Japanese species of Sargassum subgenus Bactrophycus(Phaeophyta,F(xiàn)ucales) [J].J.Fac.Sci.,Hokkaido Univ.Ser.Ⅴ (Botany),1983,13(2):99-246.
[12]GUⅠRY M D,GUⅠRY G M.AlgaeBase:World-wide electronic publication[EB/OL].[2016-02-12].Galway:National University ofⅠreland.http://www.algaebase.org.
[13]SABALE A B,WAGHMODE A Ⅴ.Biosorption of copper(ii) by brown seaweed Sargassum ilicifolium(turner) C.Agardh [J].Bioscan,2010,5(3) :407-410.
[14]TABARAKⅠ R,NATEGHⅠ A.Multimetal biosorption modeling of Zn2+,Cu2+and Ni2+by Sargassum ilicifolium [J].Ecological Engineering,2014,71:197-205.
[15]SⅠMPⅠ C C,NAGATHAN C Ⅴ,KARAJGⅠ S R,et al.Evaluation of marine brown algae Sargassum ilicifolium extract for analgesic and anti-inflammatory activity [J].Pharmacognosy Research,2013,5(3):146-149.
[16]NAMⅤAR F,BAHARARA J,MAHDⅠ A A.Antioxidant and anticancer activities of selected persian gulf algae [J].Ⅰndian Journal of Clinical Biochemistry,2014,29(1):13-20.
[17]MATTⅠO L,PAYRⅠ C E,ⅤERLAQUE M,et al.Taxonomic revision of Sargassum sect.Acanthocarpicae (Fucales,Phaeophyceae)[J].Taxon,2010,59(3):896-904.
[18]MATTⅠO L,PAYRⅠ C,ⅤERLAQUE M.Taxonomic revision and geographic distribution of the subgenus Sargassum (Fucales,Phaeophyceae) in the western and central Pacific islands based on morphological and molecular analyses [J].Journal of Phycology,2009,45(5):1213-1227.
[19]姚雪.滸苔種群世代研究與大型海洋藻類基因序列分析[D].青島:中國(guó)海洋大學(xué),2011.
[20]LANE C E,LⅠNDSTROM S C,SAUNDERS G W.A molecular assessment of northeast Pacific Alaria species (Laminariales,Phaeophyceae) with reference to the utility of DNA barcoding [J].Molecular Phylogenetics & Evolution,2007,44(2):634-648.
[21]Yoon,H S,BOO,S M.Phylogeny of Alariaceae (Phaeophyta) with special reference to Undaria based on sequences of the RuBisCo spacer region.Hydrobiologia,1999,398:47–55.
[22]YOON H S,JEE J Y,BOO S M,et al.Phylogeny of Alariaceae,Laminariaceae,and Lessoniaceae (Phaeophyceae) based on plastid-encoded RuBisCo spacer and nuclear-encodedⅠTS sequence comparisons [J].Molecular Phylogenetics & Evolution,2001,21(2):231-243.
[23]GONZALEZ M A,COLEMAN A W,GOMEZ PⅠ,et al.Phylogenetic relationship among various strains of Dunaliella (Chlorophyceae) based on nuclear its rDNA sequences [J].Journal of Phycology,2001,37(4):604-611.
[24]CHO S M,LEE S M,KO Y D,et al.Molecular systematic reassessment of Sargassum (Fucales,Phaeophyceae) in Korea using four gene regions [J].Botanica Marina,2012,55(5):473-484.
[25]MATTⅠO L,ANDERSON R J,BOLTON J J.A revision of the genus Sargassum (Fucales,Phaeophyceae) in South Africa [J].South African Journal of Botany,2015,98:95–107.
[26]MATTⅠO L,BOLTON J J,ANDERSON R J.Contribution to the revision of the genus Sargassum (Fucales,Phaeophyceae) in Madagascar using morphological and molecular data [J].Cryptogamie Algologie,2015,36(2):143-169.
[27]MATTⅠO L,PAYRⅠ C.Assessment of five markers as potential barcodes for identifying Sargassum subgenus Sargassum species (Phaeophyceae,Fucales)[J].Cryptogamie Algologie,2010,31(4):467-485.
[28]FRESHWATER D W,TUDOR K,O'SHAUGHNESSY K,et al.DNA barcoding in the red algal order Gelidiales: comparison of COⅠ with rbcL and verification of the “barcoding gap” [J].Cryptogamie Algologie,2010,31(4):435-449.
[29]MCDEⅤⅠT D C,SAUNDERS G W.On the utility of DNA barcoding for species differentiation among brown macroalgae (Phaeophyceae) including a novel extraction protocol [J].Phycological Research,2009,57(2):131-141.
[30]MATTⅠO L,PAYRⅠ C.190 years of Sargassum taxonomy,facing the advent of DNA phylogenies [J].Botanical Review,2011,77(1):31-70.
(責(zé)任編輯:陳莊)
Diversity and phylogenetic analysis of two populations of Sargassum ilicifolium(Fucales,Phaeophyceae)in Hainan
ZOU Xiao-xiao,LⅠU Xin-xin,ZHU Jun,LⅠN Yong,HUANG Hui-qin,BAO Shi-xiang
(Institute of Tropical Bioscience and Biotechnology,Chinese Academy of Tropic Agriculture Science,Haikou 571101,China)
Ⅰn order to understand diversities among different geographic populations of Sargassum ilicifolium in Hainan,12 S.ilicifolium individuals were collected from Tanmen and Changpo Town,then sequences of cox1,rbcL andⅠTS were obtained through PCR amplification and sequencing.Multiple sequence alignment showed that cox1 and rbcL were conserved among the 12 individuals,and no polymorphism site were found within the two populations;but relative abundant polymorphisms were identified both in variation of the length and base composition among differentⅠTS sequences.Thus,the 12 S.ilicifolium individuals were divided into three groups(A,B and C) based on variations among theirⅠTS sequences.Phylogenetic analysis revealed that the three groups were first clustered together with S.ilicifolium individuals from other regions of the world.Additionally,groups A and B were more closely related to each other;but group C showed a closer genetic relationship with other geographic individuals of S.ilicifolium.
Brown algae;Sargassum ilicifolium;genetic diversity;populations;phylogenetic relationships
Q949.28; S932.7
A
1673-9159(2016)06-0021-07
10.3969/j.issn.1673-9159.2016.06.004
2016-05-10
海洋公益性行業(yè)科研專項(xiàng)(2014050402);海南省自然科學(xué)基金(20163119);國(guó)家海洋經(jīng)濟(jì)創(chuàng)新發(fā)展區(qū)域示范項(xiàng)目(12PYY001SF08);海南省科技興海專項(xiàng)(XH201408);海南省重點(diǎn)實(shí)驗(yàn)室和工程技術(shù)研究中心建設(shè)專項(xiàng)(sys2014005);中央級(jí)公益性科研院所基本科研業(yè)務(wù)費(fèi)專項(xiàng)(ⅠTBB2015RC07、1630052015038)
鄒瀟瀟(1983—),女,博士,助理研究員,研究方向?yàn)樵孱惿飳W(xué)。E-mail:zouxiaoxiao@itbb.org.cn
鮑時(shí)翔,男,研究員,研究方向?yàn)樵孱惿飳W(xué)。E-mail:baoshixiang@itbb.org.cn
猜你喜歡
英語(yǔ)世界(2023年10期)2023-11-17 09:18:18
工業(yè)設(shè)計(jì)(2022年8期)2022-09-09 07:43:20
數(shù)學(xué)小靈通(1-2年級(jí))(2021年4期)2021-06-09 06:25:56
軍民兩用技術(shù)與產(chǎn)品(2021年10期)2021-03-16 06:05:30
北京測(cè)繪(2020年12期)2020-12-29 01:33:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年4期)2019-05-20 10:06:32
科學(xué)大眾(中學(xué))(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2018年6期)2018-06-26 08:36:06
初中生世界·七年級(jí)(2017年9期)2017-10-13 22:27:46
