切除單側(cè)眼柄對不同體質(zhì)量養(yǎng)殖斑節(jié)對蝦卵巢發(fā)育及產(chǎn)卵的影響
2016-12-19 06:41:52楊其彬邱麗華黃建華周發(fā)林溫為庚
水產(chǎn)科學 2016年5期
關(guān)鍵詞:質(zhì)量
楊其彬,邱麗華,黃建華,周發(fā)林,溫為庚
( 中國水產(chǎn)科學研究院 南海水產(chǎn)研究所,農(nóng)業(yè)部南海漁業(yè)資源開發(fā)利用重點實驗室,廣東 廣州 510300 )
切除單側(cè)眼柄對不同體質(zhì)量養(yǎng)殖斑節(jié)對蝦卵巢發(fā)育及產(chǎn)卵的影響
楊其彬,邱麗華,黃建華,周發(fā)林,溫為庚
( 中國水產(chǎn)科學研究院 南海水產(chǎn)研究所,農(nóng)業(yè)部南海漁業(yè)資源開發(fā)利用重點實驗室,廣東 廣州 510300 )
在鹽度31、水溫28~29 ℃下,進行了4次卵巢誘導試驗,研究了切除單側(cè)眼柄對體質(zhì)量56.4~105.4 g的養(yǎng)殖斑節(jié)對蝦卵巢發(fā)育和產(chǎn)卵的影響及卵巢發(fā)育、產(chǎn)卵與體質(zhì)量之間的關(guān)系。結(jié)果顯示,切除單側(cè)眼柄誘導養(yǎng)殖斑節(jié)對蝦卵巢發(fā)育成熟,成功率(y)與雌蝦體質(zhì)量(x)顯著相關(guān)(P<0.05),雌蝦體質(zhì)量越小,誘導成功率越低,回歸方程為:y=133.92lnx-535.63(r2=0.9297)。養(yǎng)殖斑節(jié)對蝦眼柄手術(shù)后,最快10 d產(chǎn)卵,部分雌蝦能重復(fù)產(chǎn)卵。產(chǎn)卵量(y)與雌蝦體質(zhì)量(x)顯著正相關(guān)(P<0.05),雌蝦體質(zhì)量越小,產(chǎn)卵量越小,回歸方程為:y=122.22lnx-472.2(r2=0.9738)。雌蝦累積死亡率、卵子直徑和卵子孵化率與雌蝦體質(zhì)量無顯著相關(guān)性(P>0.05)。建議用體質(zhì)量80 g以上的養(yǎng)殖斑節(jié)對蝦雌蝦作為親蝦。
斑節(jié)對蝦;卵巢;成熟;產(chǎn)卵;養(yǎng)殖
斑節(jié)對蝦(Penaeusmonodon)最大體長可達350 mm,體質(zhì)量達500 g,是對蝦屬中個體最大的養(yǎng)殖對蝦。斑節(jié)對蝦活力強、食性雜,廣泛分布于非洲南部和東部海岸、印度、巴基斯坦、馬來西亞、泰國、日本和中國等沿海,中國的浙江、福建、臺灣、廣東、廣西和海南沿海有分布[1]。
傳統(tǒng)的斑節(jié)對蝦親蝦均源于自然海區(qū),但親蝦資源日漸枯竭,不能滿足斑節(jié)對蝦養(yǎng)殖業(yè)的需求。有研究發(fā)現(xiàn),野生斑節(jié)對蝦親蝦攜帶病毒的比例極高,如泰國野生斑節(jié)對蝦樣品攜帶白斑綜合癥病毒的比例高達89.8%[2],這就增加了養(yǎng)殖斑節(jié)對蝦失敗的幾率。近年斑節(jié)對蝦有逐漸被經(jīng)過選育的“生長快、無特定病原”的凡納濱對蝦(Litopenaeusvannamei)改良品系所取代的趨勢[3],養(yǎng)殖面積顯著減少。因此,開展斑節(jié)對蝦良種培育,獲得生長快、無特定病原的斑節(jié)對蝦新品系,是斑節(jié)對蝦養(yǎng)殖業(yè)持續(xù)、穩(wěn)定發(fā)展的需要。而這首先要研究養(yǎng)殖斑節(jié)對蝦性成熟和卵巢誘導成熟及產(chǎn)卵的規(guī)律。已有的研究表明,養(yǎng)殖的斑節(jié)對蝦性成熟和繁殖十分困難,親蝦的成熟率、產(chǎn)卵量和孵化率都很低,難以滿足規(guī)模化苗種生產(chǎn)需求[4-6],也是斑節(jié)對蝦全人工繁殖技術(shù)的關(guān)鍵。黃建華等[7]跟蹤研究了養(yǎng)殖斑節(jié)對蝦的性成熟,認為養(yǎng)殖斑節(jié)對蝦雌蝦性成熟與體質(zhì)量顯著相關(guān)。這與養(yǎng)殖墨吉明對蝦(Fenneropenaeusmerguiensis)[8]和凡納濱對蝦[9]的性成熟與其個體大小密切相關(guān)一致。但目前尚無有關(guān)養(yǎng)殖斑節(jié)對蝦卵巢誘導成熟及產(chǎn)卵與體質(zhì)量關(guān)系的試驗支持這一觀點。
在前人研究基礎(chǔ)上,筆者推測,養(yǎng)殖斑節(jié)對蝦卵巢誘導發(fā)育成熟及產(chǎn)卵可能與體質(zhì)量密切相關(guān)。本研究設(shè)計了4批次試驗,切除不同體質(zhì)量(56.4~105.4 g)的養(yǎng)殖斑節(jié)對蝦雌蝦的單側(cè)眼柄,誘導其卵巢快速發(fā)育成熟并產(chǎn)卵,跟蹤分析養(yǎng)殖斑節(jié)對蝦卵巢誘導成熟及產(chǎn)卵與體質(zhì)量之間的關(guān)系,為斑節(jié)對蝦全人工繁殖提供參考。
1 材料與方法
1.1 材料
1.1.1 試驗條件
于2013和2014年春季和秋季在南海水產(chǎn)研究所三亞基地先后進行了4次誘導卵巢成熟試驗,每次持續(xù)30 d。試驗池為室內(nèi)水泥池,面積1 m2的80口,20 m2的4口,深度1.3 m。試驗用水為80 cm厚的細沙層過濾、沉淀24 h的自然海水,鹽度31,溫度28~29oC,pH 8.3,再經(jīng)250目篩絹袋過濾。餌料為市場購買的魷魚、蟹肉、貝肉和活沙蠶。
1.1.2 試驗蝦的準備及管理
斑節(jié)對蝦為試驗基地培育的蝦苗(全長1.2 cm),投放到三亞林旺養(yǎng)殖示范基地的石斑魚(Epinephelus)養(yǎng)殖池塘(面積0.5~0.8 hm2)中混養(yǎng)。蝦苗密度為15尾/m2, 魚種(全長5.0 cm)密度0.5 尾/m2。早晚各投喂一次雜魚磨成的魚漿。在15口塘養(yǎng)殖266 d后,挑選附肢完整、彈跳力強、色澤鮮亮、帶有精莢的不同體質(zhì)量的雌蝦60尾,最小56.4 g,最大105.4 g。10 g為一個梯度范圍,4次誘導卵巢成熟試驗雌蝦體質(zhì)量分布范圍見表1。同時挑選雄蝦30尾,運回試驗基地,分別投放到室內(nèi)2口20 m2的水池中,加入深度50 cm海水,暫養(yǎng)5 d,每日8:00和19:00過量投喂餌料。每日18:00用虹吸法清除殘餌及糞便,換水50%。
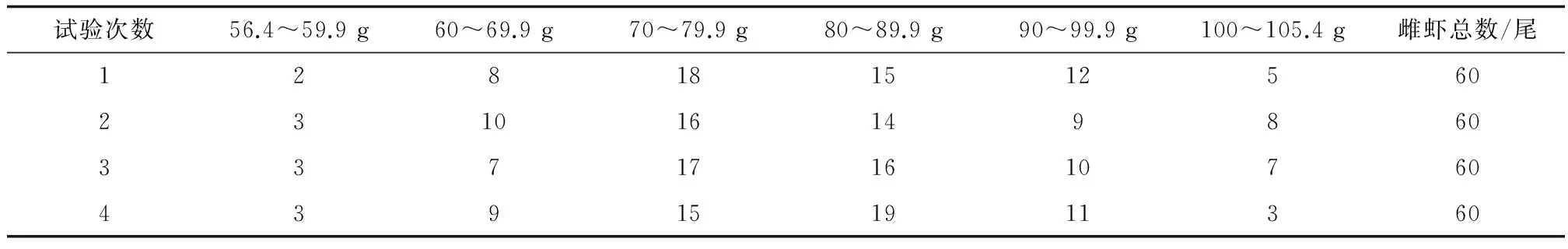
表1 各次試驗中雌蝦的體質(zhì)量范圍及數(shù)量
1.2 方法
根據(jù)文獻[10]的方法剪除單邊眼柄。剪眼柄蝦放在同一個20 m2水池(催熟池),同時投放30尾雄蝦,雌雄比例2∶1,最終養(yǎng)殖密度4.5 尾/m2。催熟水池用黑布遮光,光照度10 lx。每日8:00、19:00、21:00過量投喂餌料。清除殘餌及換水和準備階段相同。
卵巢的發(fā)育分期參照文獻[11-12],外觀觀察參照文獻[13]描述的方法進行。剪眼柄5 d后,每日19:00—20:00用潛水手電筒照射雌蝦腹部,仔細檢查卵巢發(fā)育情況。若卵巢達到Ⅲ期或以上即可移至產(chǎn)卵池(1 m2),加入試驗用水至1 m深。雌蝦產(chǎn)卵后,記錄其體質(zhì)量并移至另一個催熟池(20 m2)中,以區(qū)別于卵巢不發(fā)育的雌蝦,同時觀察卵巢再發(fā)育效果。未產(chǎn)卵的雌蝦移回催熟池或者讓其在產(chǎn)卵池繼續(xù)待產(chǎn)。
每日9:00左右,將已產(chǎn)卵的池水充分攪動,讓卵子盡可能分布均勻,從池四個角和中間的淺、深各處取樣100 mL,在顯微鏡下計數(shù)卵子數(shù)量,測量卵子直徑,計算樣品平均值(n)。卵子在產(chǎn)卵池中留2 d,孵化成無節(jié)幼體后,按照前述方法估算無節(jié)幼體數(shù)量。
每日記錄不同體質(zhì)量雌蝦的死亡數(shù)、產(chǎn)卵雌蝦數(shù)、卵巢達到Ⅲ期(及以上)雌蝦數(shù)(不含卵巢重復(fù)發(fā)育的雌蝦)、產(chǎn)卵量、無節(jié)幼體數(shù)量和卵子直徑。計算下列指標:
總產(chǎn)卵數(shù)量/104粒·池-1=S×n1
無節(jié)幼體數(shù)/104尾·池-1=S×n2
孵化率/%=n3/n4×100%
累積死亡率/%=n5/n6×100%
誘導成功率/%=n7/n6×100%
式中,S為產(chǎn)卵池面積(m2),n1為卵子樣品平均值(104粒/m2),n2為無節(jié)幼體樣品平均值(104尾/m2),n3為無節(jié)幼體估計數(shù),n4為卵子估計數(shù),n5為死亡雌蝦數(shù),n6為剪眼柄雌蝦數(shù),n7為卵巢Ⅲ期(及以上)雌蝦數(shù)。
1.3 數(shù)據(jù)處理
各次試驗中,將不同體質(zhì)量雌蝦的誘導成功率、產(chǎn)卵量和無節(jié)幼體數(shù)量逐個在Microsoft Excel 2010上作散點圖,進行回歸分析,得出回歸方程及其顯著性。以表1作為體質(zhì)量范圍,計算同一體質(zhì)量范圍該次試驗孵化率、卵子直徑、死亡率結(jié)果數(shù)值和最終4次試驗結(jié)果的平均值,在Microsoft Excel 2010上作散點圖,對最終結(jié)果進行回歸分析,得出其回歸方程及顯著性。
2 結(jié) 果
2.1 誘導后斑節(jié)對蝦卵巢的發(fā)育
在試驗中,不同體質(zhì)量的斑節(jié)對蝦雌蝦被剪除單側(cè)眼柄后,在適宜條件(暗光、高蛋白餌料等)下,卵巢發(fā)育較快,6 d就能觀察到清晰的卵巢。不同體質(zhì)量雌蝦卵巢發(fā)育到Ⅲ期(及以上)的尾數(shù)占手術(shù)雌蝦總數(shù)的比例(誘導成功率)不同,體質(zhì)量56.4~59.9 g的雌蝦平均誘導成功率為8.2%;60~69.9 g誘導成功率為8.4%;70~79.9 g誘導成功率為48.3%;80~89.9 g誘導成功率為67.4%;90~99.9 g誘導成功率為73.6%;100~105.4 g誘導成功率為81.6%。養(yǎng)殖斑節(jié)對蝦卵巢誘導成功率與其體質(zhì)量呈顯著的正相關(guān)(P<0.05):體質(zhì)量越小,卵巢誘導成功率越低。誘導成功率與體質(zhì)量成對數(shù)關(guān)系:y=133.92lnx-535.63(r2=0.9297)(圖1)。
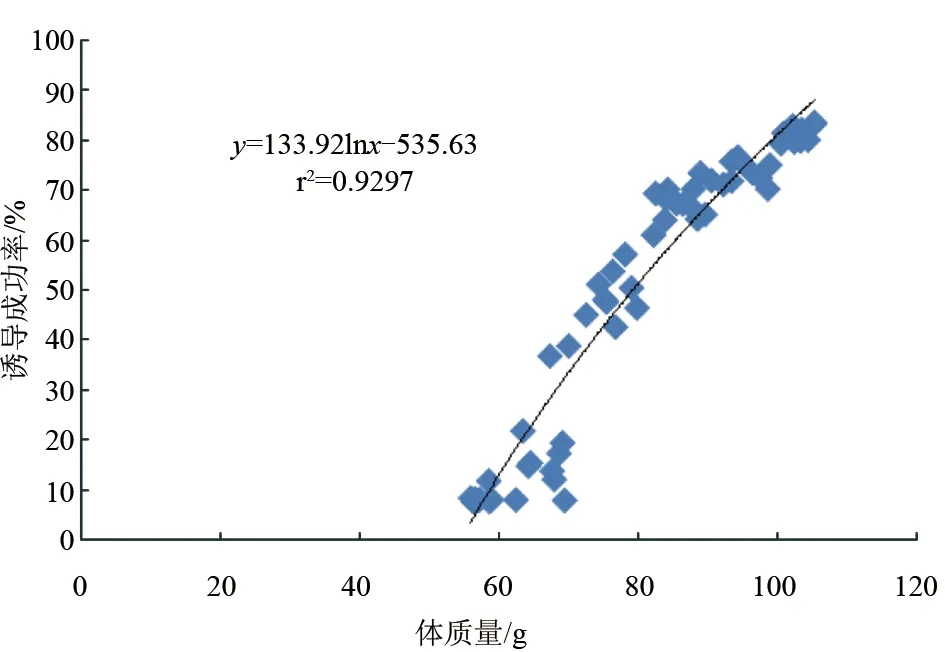
圖1 不同體質(zhì)量養(yǎng)殖斑節(jié)對蝦的誘導成功率
2.2 誘導后斑節(jié)對蝦產(chǎn)卵
養(yǎng)殖斑節(jié)對蝦雌蝦通過人工誘導(剪除單側(cè)眼柄)發(fā)育成熟加快,最快10 d就產(chǎn)卵。體質(zhì)量56.4~59.9 g雌蝦產(chǎn)卵1.5×105粒,60~69.9 g雌蝦產(chǎn)卵2.17×105粒(1.95×105~2.43×105粒),70~79.9 g雌蝦產(chǎn)卵4.38×105粒(3.61×105~5.16×105粒),80~89.9 g雌蝦產(chǎn)卵7.22×105粒(6.22×105~8.03×105粒),90~99.9 g雌蝦產(chǎn)卵7.75×105粒(6.54×105~8.95×105粒),100~105.4 g以上產(chǎn)卵量為8.73×105粒(7.80×105~9.66×105粒)。雌蝦產(chǎn)卵量與其體質(zhì)量呈顯著正相關(guān)(P<0.05),體質(zhì)量越大,產(chǎn)卵越多。產(chǎn)卵量與體質(zhì)量成對數(shù)關(guān)系:y=122.22lnx-472.2(r2=0.9738)(圖2)。
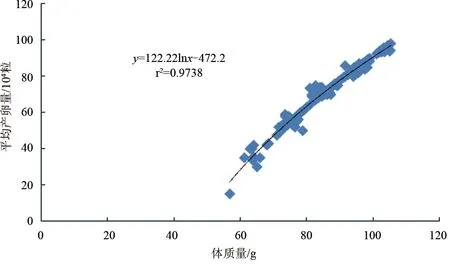
圖2 不同體質(zhì)量養(yǎng)殖斑節(jié)對蝦平均單次產(chǎn)卵量
斑節(jié)對蝦卵約28 h孵出無節(jié)幼體,平均單次無節(jié)幼體產(chǎn)量與雌蝦體質(zhì)量密切相關(guān)(P<0.05)。56.4~59.9 g雌蝦平均一次產(chǎn)無節(jié)幼體6.0×104尾,60~69.9 g產(chǎn)無節(jié)幼體1.25×105尾,70~79.9 g產(chǎn)無節(jié)幼體2.80×105尾,80~89.9 g產(chǎn)無節(jié)幼體5.17×105尾,90~99.9 g產(chǎn)無節(jié)幼體5.50×105尾,100~105.4 g產(chǎn)無節(jié)幼體6.81×105尾。無節(jié)幼體產(chǎn)量與雌蝦體質(zhì)量成對數(shù)關(guān)系:y=95.964lnx-374.51(r2=0.9475)(圖3)。整個試驗過程中,卵子孵化率最高82.2%,最低40.0%。各體質(zhì)量梯度范圍(由低到高)平均孵化率依次是:10%、57.6%、63.7%、71.6%、70.7%、75.6%。卵子孵化率與雌蝦體質(zhì)量的線性或?qū)?shù)關(guān)系均不顯著(P>0.05)(圖4)。斑節(jié)對蝦卵直徑為268.7~312.1 μm,各體質(zhì)量梯度范圍(由低到高)卵直徑平均值依次為:287.3 μm、291.0 μm、291.1 μm、293.7 μm、290.3 μm、291.6 μm,與雌蝦體質(zhì)量無顯著線性或?qū)?shù)關(guān)系(P>0.05)(圖5)。
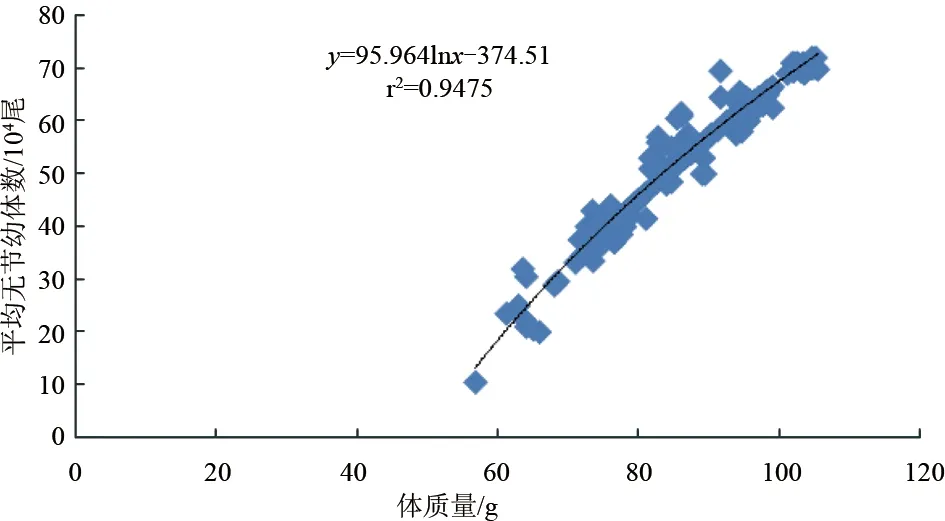
圖3 不同體質(zhì)量養(yǎng)殖斑節(jié)對蝦平均單次無節(jié)幼體產(chǎn)量
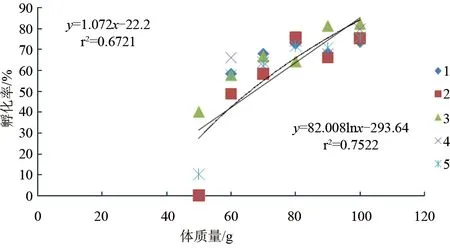
圖4 不同體質(zhì)量養(yǎng)殖斑節(jié)對蝦卵子孵化率注:系列1、2、3和4是該次試驗結(jié)果平均值;系列5是4次試驗結(jié)果的總平均值。為簡潔,只畫出“5”的趨勢線.下同.
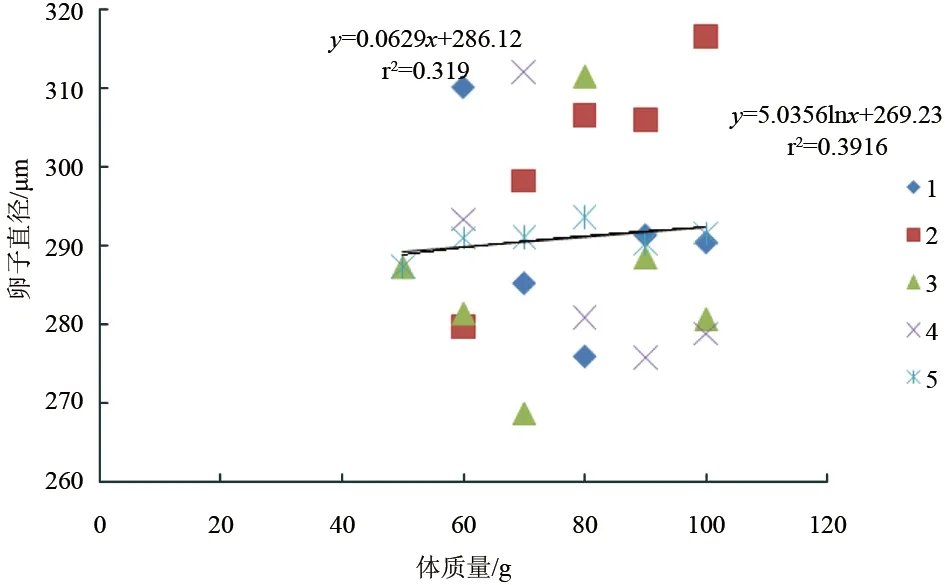
圖5 不同體質(zhì)量養(yǎng)殖斑節(jié)對蝦卵子直徑
2.3 斑節(jié)對蝦的誘導死亡率
剪除單側(cè)眼柄能誘導養(yǎng)殖斑節(jié)對蝦卵巢快速發(fā)育,也會導致對蝦死亡,最高死亡率為50%,平均值13.8%~23.0%。各體質(zhì)量梯度范圍(由低至高)死亡率平均值依次是18.8%、17.6%、13.8%、16.8%、16.5%和23.0%,與雌蝦體質(zhì)量無顯著線性或?qū)?shù)關(guān)系(P>0.05)(圖6)。

圖6 不同體質(zhì)量養(yǎng)殖斑節(jié)對蝦雌蝦眼柄手術(shù)后的死亡率
3 討 論
3.1 關(guān)于誘導成功率
本試驗跟蹤觀察到,養(yǎng)殖斑節(jié)對蝦的誘導成功率與其體質(zhì)量顯著相關(guān)(P<0.05),體質(zhì)量越小誘導成功率越低。體質(zhì)量80 g以上的雌蝦誘導成功率67.4%以上,與野生蝦的60%~90%相當[10],而體質(zhì)量80 g(不含80 g)以下的雌蝦誘導成功率為48.3%以下。黃建華等[7]的跟蹤研究顯示,養(yǎng)殖斑節(jié)對蝦雌蝦性成熟的最小體質(zhì)量為75.0 g,只有性成熟后的養(yǎng)殖斑節(jié)對蝦雌蝦才容易誘導成熟產(chǎn)卵,體質(zhì)量80 g以上的雌蝦誘導成功率較高。本試驗結(jié)果與Chotipuntu等[14]的結(jié)果“養(yǎng)殖斑節(jié)對蝦第一次卵巢成熟平均體質(zhì)量為(78 ± 4) g (n=2)”一致。對其他對蝦如墨吉明對蝦[8]和凡納濱對蝦[9]的研究也發(fā)現(xiàn),性成熟與體質(zhì)量密切相關(guān)。另外,卵巢誘導成熟還有其他條件配合。Hoang等[8,10]認為,暗光有利于卵巢成熟;Xu等[15]認為,優(yōu)質(zhì)餌料(如魚肉、維生素C和魚油)在水生動物繁殖中起重要作用。該試驗采用了暗光和鮮活餌料,可能這是導致少數(shù)體質(zhì)量較小的雌蝦被誘導發(fā)育成熟的原因。試驗蝦的養(yǎng)殖日齡對卵巢誘導效果的影響,有待以后深入研究。
3.2 關(guān)于產(chǎn)卵量
本試驗跟蹤觀察到,剪除單側(cè)眼柄后,養(yǎng)殖斑節(jié)對蝦的產(chǎn)卵量與體質(zhì)量顯著相關(guān)(P<0.05):體質(zhì)量80 g以上的雌蝦平均單次產(chǎn)卵7.22×105粒,最高9.66×105粒;體質(zhì)量70 g以下雌蝦平均單次產(chǎn)卵4.38×105粒,最低1.5×105粒。高于Arnold等[16]的研究結(jié)果“第八代養(yǎng)殖斑節(jié)對蝦平均每次產(chǎn)卵量4.13×105粒”。Peixoto等[17]認為,卵巢成熟度顯著影響產(chǎn)卵量。同一成熟度的卵巢,性腺指數(shù)相似,雌蝦體質(zhì)量越大,其卵巢就越大,所含的卵子越多,產(chǎn)卵量越大,與Menasevta等[18]的研究結(jié)果一致。所以,筆者建議用體質(zhì)量80 g以上的養(yǎng)殖斑節(jié)對蝦雌蝦作為親蝦。
3.3 關(guān)于雌蝦死亡率
本試驗中,剪除單側(cè)眼柄導致試驗雌蝦累積死亡率為13.8%~23.0%,略高于Emerenciano等[19]的研究結(jié)果(6%~12%),雌蝦死亡率與其體質(zhì)量無顯著關(guān)系(P>0.05)。剪除眼柄是破壞性手術(shù),損傷試驗蝦,甚至死亡。用夾燙法剪除眼柄效果最好,試驗蝦存活率幾乎達到100%[10]。本試驗中,養(yǎng)殖斑節(jié)對蝦雌蝦的死亡可能是剪眼柄手術(shù)所致,與體質(zhì)量無顯著關(guān)系。目前,已有學者研究了降低卵巢誘導發(fā)育中蝦死亡率的新方法,如Meeratana等[18]用注射血青素誘導羅氏沼蝦(Macrobrachiumrosenbergii)卵巢成熟,Vaca等[20]用血青素誘導凡納濱對蝦卵巢成熟,溫為庚等[13,21]用血青素誘導斑節(jié)對蝦卵巢成熟。此方法雖然能降低蝦類的死亡率,但操作復(fù)雜,還有待深入研究。
3.4 關(guān)于孵化率
本試驗中,卵子孵化率為40.0%~82.2%,與雌蝦體質(zhì)量無顯著關(guān)系(P>0.05)。影響卵子孵化率的因素很多[22],但Arnold等[23]認為,精莢質(zhì)量、精子數(shù)量和質(zhì)量顯著影響孵化率,而Coman等[24]認為,卵子的質(zhì)量顯著影響孵化率;Ramesh[25]認為,天然餌料能改善卵子質(zhì)量;Meunpol等[26]則認為,餌料是影響雄蝦繁殖性能的重要因子。養(yǎng)殖斑節(jié)對蝦生存環(huán)境局限在池塘中,不同體質(zhì)量的對蝦攝食的餌料主要是人工投喂的餌料,差異不顯著,對精子和卵子質(zhì)量影響不顯著,對孵化率影響不顯著。
經(jīng)過多年選育,斑節(jié)對蝦對餌料的利用率有所提高[27],繁殖力有改善[28],但繁殖效率依然較低。如何提高斑節(jié)對蝦的繁殖效率和培育無特定病原斑節(jié)對蝦親蝦,有待深入研究。
[1] 劉瑞玉,鐘振如. 南海對蝦類[M]. 北京:農(nóng)業(yè)出版社,1986:106-108.
[2] 楊麗詩,黃建華,孫苗苗,等. 3種來源的野生斑節(jié)對蝦攜帶病毒情況調(diào)查[J]. 上海海洋大學學報,2011, 20(4):546-552.
[3] Wyban J. Domestication of Pacific white shrimp revolutionizes aquaculture [J]. G Aqua Ad, 2007(4):42-44.
[4] Coman G J, Crocos P J, Amold S J, et al. Growth, survival and reproductive performance of domesticated Australian stock of the giant tiger shrimp,Penaeusmonodon, reared in tanks and raceways [J]. J W Aqua So, 2005, 36(4):464-479.
[5] Coman G J, Amold S J, Peixoto S, et al. Reproductive performance of reciprocally crossed wild-caught and tank rearedPenaeusmonodonbroodstock [J]. Aquaculture, 2006, 252(2/4):372-384.
[6] Coman G J, Amold S J, Callaghal T R, et al. Effect of two maturation diet combination on reproductive performance of domesticatedPenaeusmonodon[J]. Aquaculture, 2007, 263(1/4):75-83.
[7] 黃建華,楊其彬,馬之明,等. 池塘養(yǎng)殖斑節(jié)對蝦的生長、發(fā)育與性成熟[J]. 水產(chǎn)學報,2013,37(3):397-406.
[8] Hoang T, Lee S Y, Keenan C P,et al. Maturation and spawning performance of pond-rearedPenaeusmerguiensisin different combinations of temperature, light intensity and photoperiod [J]. Aqua Res, 2002, 33(15):1243-1252.
[9] Ceballos-Vazquez B P, Rosas C, Racotta I S. Sperm quality in relation to age and weight of white shrimpLitopenaeusvannamei[J]. Aqua, 2003,228(1/4):141-151.
[10] 陳明耀,馮玉愛,梁華芳,等. 斑節(jié)對蝦親蝦卵巢催熟若干問題的探討[J]. 湛江水產(chǎn)學院學報,1992, 12(1):1-6.
[11] 黃建華,馬之明,周發(fā)林,等. 南海北部野生斑節(jié)對蝦卵巢解剖結(jié)構(gòu)及組織學的研究[J]. 南方水產(chǎn),2005,1(3):49-53.
[12] 黃建華,周發(fā)林,馬之明,等. 南海北部斑節(jié)對蝦卵巢發(fā)育的形態(tài)及組織學觀察[J]. 熱帶海洋學報, 2006,25(3):47-52.
[13] 溫為庚,黃建華,楊其彬,等. 血清素在斑節(jié)對蝦催熟過程中的作用[J]. 南方水產(chǎn),2009,5(1):59-63.
[14] Chotipuntu P, Wuthisuthimethayee S, Direkbusrakom S,et al.Reproductive aspects of SPFPenaeusmonodongrown in closed culture captivity[J]. Ag Tech Bios, 2013,10(3):227-236.
[15] Xu J Y, Wang T T, Wang Y F, et al. Effect of combined fish meal, soyabean meal ratio, vitamin-C, and fish oil supplementations in diet on the growth and reproduction of red shrimp, crayfish,Procambarusclarkii(Crustacea Decapoda)[J]. Aquaculture Research, 2010,41(9):252-259.
[16] Arnold S J,Coman G J, Emerenciano M. Constraints on seedstock production in eighth generation domesticatedPenaeusmonodonbroodstock[J].Aqua, 2013(410/411):95-100.
[17] Peixoto S, Coman G J, Arnold S J, et al. Histological examination of final oocyte maturation and atresia in wild and domesticatedPenaeusmonodon(Fabricius) broodstock[J]. Aqua Res, 2005, 36(7):666-673.
[18] Meeratana P, Withyachamnarnkul B, Damrongphol P,et al. Serotonin induces ovarian maturation in giant freshwater prawn broodstock,Macrobrachiumrosenbergiide Man[J]. Aqua, 2006,160 (1/2):315-325.
[19] Emerenciano M, Cuzon G, Mascaro M,et al. Reproductive performance, biochemical composition and fatty acid profile of wild-caught and 2nd generation domesticatedFarfantepenaeusduorarum(Burkenroad,1939) broodstock [J]. Aqua, 2012(344/349):194-204.
[20] Vaca A A, Alfaro J. Ovarian maturation and spawning in the white shrimp,Penaeusvannamei, by serotonin injection [J]. Aqua, 2000,182 (3/4):373-385.
[21] Wongprasert K, Asuvapongpatana S, Poltana P, et al. Serotonin stimulates ovarian maturation and spawning in the black tiger shrimp,Penaeusmonodon[J].Aqua, 2006, 261(3):1447-1454.
[22] Wouter R, Lavens P, Nieto J,et al. Penaeid shrimp broodstock nutrition: an update review on research and development[J]. Aqua, 2001,202(1/2):1-21.
[23] Arnold S J,Coman G J, Burridge C, et al. A novel approach to evaluate the relationship between measures of male fertility and egg fertilization inPenaeusmonodon[J]. Aqua, 2012(338/341):181-189.
[24] Coman G J, Arnold, S J, Callaghan T R, et al.Effect of two maturation diet combinations on reproductive performance of domesticatedPenaeusmonodon[J]. Aqua, 2007, 263(1):75-83.
[25] Ramesh B K. Improved maturation of wild and pond-reared black tiger shrimpPenaeusmonodon(Fabricius) using different combinations of live and wet feeds[J]. Asia Jour Ex, 2013,27(2):37-42.
[26] Meunpol O, Meejing P,Piyatiratitivorakul S. Maturation diet based on fatty acid content for malePenaeusmonodon(Fabricius) broodstock[J]. Aquac Res,2005,36(12):1216-1225.
[27] Glencross B, Tabrett S, Irvin S, et al. An analysis of the effect of diet and genotype on protein and energy utilization by the black tiger shrimp,Penaeusmonodon—why do genetically selected shrimp grow faster? [J]. Aqua Nut, 2013, 19(2):128-138.
[28] Coman, G J, Arnold, S J, Wood, A T, et al. Evaluation of egg and nauplii production parameters of a single stock of domesticatedPenaeusmonodon(giant tiger shrimp) across generations[J]. Aqua, 2013(400/401):125-128.
EffectsofUnilateralEyestalkAblationonOvaryDevelopmentandSpawningofCulturedTigerShrimpPenaeusmonodonwithDifferentBodyWeight
YANG Qibin, QIU Lihua, HUANG Jianhua, ZHOU Falin, WEN Weigeng
( Key Laboratory of South China Sea Fishery Resources Exploitation and Utilization, Ministry of Agriculture, South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Guangzhou 510300, China )
The effects of unilateral eyestalk ablation on ovary development and spawning were studied in cultured tiger shrimpPenaeusmonodonwith body weight from 56.4 g to 105.4 g in 4 trials to evaluate the relationship between ovary development and spawning and body weight of cultured tiger shrimp. The results showed that ovary maturation of cultured tiger shrimp was induced by unilateral eyestalk ablation, with maturation success rate (%), significantly related to the body weight of female logarithmically (P<0.05):y=133.92lnx-535.63(r2=0.9297). The smaller female was, the lower maturation success rate was. The tiger shrimp females with eyestalk ablated spawned as early as the 10th day after ablation, some females spawning repeatedly. The fecundity was found to be significantly positively related to the body weight of female logarithmically (P<0.05):y=122.22lnx-472.2(r2=0.9738), the smaller the female was, the less egg production was. The accumulative mortality of female, egg diameter and hatching rate were not significantly related to the body weight of female. The cultured tiger shrimp females with body weight of ≥80.0 g is recommended for broodstock.
Penaeusmonodon; ovary; maturation; spawning; culture
10.16378/j.cnki.1003-1111.2016.05.010
S968.22
A
1003-1111(2016)05-0516-06
2016-01-06;
2016-04-05.
海南省重點科技計劃項目(ZDXM2014057);海南省社會發(fā)展項目(SF201439).
楊其彬(1972—),男,助理研究員;研究方向:對蝦遺傳育種. E-mail:yangqibin1208@163.com. 通信作者:溫為庚(1963—),男,副研究員;研究方向:對蝦基礎(chǔ)生物學.E-mail: wenweigeng@163.com.
猜你喜歡
中學生數(shù)理化·中考版(2022年10期)2022-11-10 09:37:42
中學生數(shù)理化·八年級物理人教版(2022年12期)2022-02-14 07:08:42
中學生數(shù)理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數(shù)理化·中考版(2020年10期)2020-11-27 01:59:48
中國生殖健康(2019年2期)2019-08-23 08:12:08
石油化工建設(shè)(2018年6期)2018-04-22 03:16:54
產(chǎn)品可靠性報告(2017年7期)2017-09-05 09:49:12
中學生數(shù)理化·八年級物理人教版(2017年12期)2017-04-18 12:59:38
汽車觀察(2016年3期)2016-02-28 13:16:26
民生周刊(2014年7期)2014-03-28 01:30:54
