飛蝗海藻糖酶基因的分子特性及功能
2016-12-23 06:40:01劉曉健孫亞文馬恩波張建珍
中國農(nóng)業(yè)科學(xué) 2016年22期
劉曉健,孫亞文,2,崔 淼,馬恩波,張建珍
?
飛蝗海藻糖酶基因的分子特性及功能
劉曉健1,孫亞文1,2,崔 淼1,馬恩波1,張建珍1
(1山西大學(xué)應(yīng)用生物學(xué)研究所,太原 030006;2山西大學(xué)生命科學(xué)學(xué)院,太原 030006)
【目的】海藻糖酶(trehalase)可專一性地將一分子海藻糖水解成為兩分子葡萄糖,在昆蟲能量代謝和幾丁質(zhì)合成過程中發(fā)揮重要作用,因此,海藻糖酶已成為害蟲控制的潛在靶標。本研究以重要農(nóng)業(yè)害蟲飛蝗()為材料,探討海藻糖酶基因的分子特性及生理功能,為飛蝗的有效治理提供理論依據(jù)。【方法】通過搜索飛蝗轉(zhuǎn)錄組和基因組數(shù)據(jù)庫,獲得4個海藻糖酶基因cDNA全長序列,運用blast、TMHMM和SignalP等相關(guān)軟件分析其序列特征;利用ClustalW進行海藻糖酶全長氨基酸序列比對,MEGA7構(gòu)建海藻糖酶的系統(tǒng)進化樹;運用reverse transcription-quantitative PCR(RT-qPCR)技術(shù)研究4個海藻糖酶基因在5齡若蟲不同組織及發(fā)育日齡的表達特性;體外合成4個基因的dsRNA后,分別注射5齡第2天若蟲,同時注射ds作為對照,收集注射dsRNA 48 h后的整蟲提取RNA,反轉(zhuǎn)錄成cDNA后,采用RT-qPCR技術(shù)檢測海藻糖酶基因以及幾丁質(zhì)合成關(guān)鍵基因UDP--乙酰葡糖胺焦磷酸化酶基因和幾丁質(zhì)合成酶1基因的表達量,并觀察記錄表型。【結(jié)果】飛蝗4個海藻糖酶均含有海藻糖酶的2個功能保守區(qū)以及1個甘氨酸富集區(qū),其中1個海藻糖酶具有跨膜結(jié)構(gòu)域,將其命名為LmTreM(GenBank登錄號:KX371563),1個海藻糖酶具有類跨膜結(jié)構(gòu),將其命名為LmTreM-like(GenBank登錄號:KX371565),其余2個均為可溶性海藻糖酶,分別命名為LmTreS1和LmTreS2(GenBank登錄號:KX371564和FJ795020);系統(tǒng)發(fā)育分析結(jié)果顯示,LmTreM和LmTreM-like與膜結(jié)合型海藻糖酶聚為一支,而LmTreS1和LmTreS2與可溶性海藻糖酶聚為一支;對4個基因在5齡若蟲不同組織部位和發(fā)育日齡表達量的分析表明、和具有組織特異表達特性,而在所有組織部位中均表達,且4個海藻糖酶基因呈現(xiàn)出不同的發(fā)育變化趨勢;RNAi結(jié)果表明,分別注射飛蝗4個海藻糖酶基因dsRNA至5齡若蟲后,與對照組相比,各基因表達量均顯著降低,且不存在基因間的交叉干擾,幾丁質(zhì)合成關(guān)鍵基因和的表達也無顯著變化,注射各海藻糖酶基因dsRNA的5齡若蟲均可成功蛻皮至成蟲。【結(jié)論】飛蝗存在1個膜結(jié)合型、1個類膜結(jié)合型和2個可溶性海藻糖酶基因,這4個基因具有不同的組織和發(fā)育表達特性,任一基因的表達沉默均不影響5齡飛蝗正常蛻皮至成蟲。
飛蝗;海藻糖酶基因;表達特性;RT-qPCR; RNA干擾
0 引言
【研究意義】飛蝗()是一種重要的農(nóng)業(yè)害蟲,蝗災(zāi)的有效防治亦是保證農(nóng)業(yè)安全生產(chǎn)的重要科學(xué)問題。目前,在飛蝗防治工作中仍以化學(xué)殺蟲劑為主要手段,因長期大量使用造成一系列問題,如污染農(nóng)業(yè)環(huán)境、對非靶標生物有負面影響以及導(dǎo)致害蟲產(chǎn)生抗藥性[1]等,研發(fā)用于飛蝗綜合治理的環(huán)境友好型殺蟲劑勢在必行。海藻糖酶(trehalase)是一類昆蟲能量代謝必不可少的酶,也是幾丁質(zhì)合成途徑的第一個酶[2-3],因此海藻糖酶可作為設(shè)計新型殺蟲劑的潛在靶標。【前人研究進展】海藻糖是昆蟲血淋巴中主要的糖類物質(zhì),海藻糖酶可將一分子海藻糖分解為兩分子葡萄糖,昆蟲海藻糖酶根據(jù)是否存在跨膜結(jié)構(gòu),可分為兩類:可溶性海藻糖酶(soluble trehalase,TreS或Tre1)與膜結(jié)合型海藻糖酶(membrane-bound trehalase,TreM或Tre2)[4]。可溶性海藻糖酶分解細胞內(nèi)的海藻糖,而膜結(jié)合型海藻糖酶分解胞外(主要為食物中)的海藻糖[5]。第一個可溶性海藻糖酶基因于1992年在黃粉蟲()中克隆獲得[6],直到2005年才在家蠶()中獲得第一個膜結(jié)合型海藻糖酶基因[7]。目前的研究表明,多數(shù)昆蟲體內(nèi)都存在1個膜結(jié)合型和1個可溶性海藻糖酶基因,僅少數(shù)昆蟲如異色瓢蟲()[8]褐飛虱()[9]中發(fā)現(xiàn)多個可溶性海藻糖酶基因。海藻糖為昆蟲體內(nèi)貯存能量的重要物質(zhì),已有研究表明,海藻糖酶能夠通過調(diào)節(jié)昆蟲體內(nèi)海藻糖的濃度抵抗低溫、干燥和農(nóng)藥等逆境脅迫。如在異色瓢蟲中,溫度逐漸降低時海藻糖酶的活性受到抑制,其中可溶性海藻糖酶4基因的mRNA表達顯著下調(diào)[8]。在干燥條件下嗜眠搖蚊()通過降低海藻糖酶的活性,使昆蟲體內(nèi)積累大量海藻糖,以保護昆蟲應(yīng)對干燥環(huán)境[10]。同時海藻糖酶也是幾丁質(zhì)合成通路中的第一個酶,在幾丁質(zhì)代謝過程中發(fā)揮著關(guān)鍵作用[3]。Chen等[11]利用RNAi技術(shù)分別將甜菜夜蛾()可溶性和膜結(jié)合型海藻糖酶基因表達沉默后,可抑制幾丁質(zhì)合成酶1和2基因的表達,從而導(dǎo)致表皮和中腸幾丁質(zhì)的合成減少最終引起昆蟲死亡;Zhao等[9]研究褐飛虱2個可溶性和1個膜結(jié)合型海藻糖酶基因的功能時,發(fā)現(xiàn)分別注射每個海藻糖酶基因的dsRNA后,昆蟲的死亡率達到14.16%—31.78%,且?guī)锥≠|(zhì)合成酶和幾丁質(zhì)酶等多個幾丁質(zhì)代謝通路的基因表達相應(yīng)下調(diào)。【本研究切入點】關(guān)于昆蟲海藻糖酶基因的已有研究大都集中在完全變態(tài)昆蟲,而飛蝗是典型的漸變態(tài)昆蟲,目前尚未在飛蝗中開展海藻糖酶基因的分子特性及功能研究。【擬解決的關(guān)鍵問題】搜索飛蝗轉(zhuǎn)錄組和基因組數(shù)據(jù)庫,獲得海藻糖酶基因cDNA序列,采用RT-qPCR技術(shù)研究在飛蝗不同組織部位和發(fā)育時期的mRNA表達特性,運用RNA干擾技術(shù)分析在飛蝗發(fā)育過程中的作用,豐富昆蟲海藻糖酶基因的研究內(nèi)容,為基于RNAi的害蟲防治提供基礎(chǔ)數(shù)據(jù)。
1 材料與方法
試驗于2012—2014年在山西大學(xué)應(yīng)用生物學(xué)研究所完成。
1.1 材料
供試昆蟲:飛蝗蟲卵購買自山西晉源北邵飛蝗養(yǎng)殖公司,置于溫度為(30±2)℃,相對濕度40%±10%,光照14 h、黑暗10 h的人工氣候箱內(nèi)進行孵化,孵化的飛蝗若蟲給予新鮮小麥幼苗和麥麩飼喂。飼喂至5齡若蟲后,選擇2 h內(nèi)的飛蝗進行試驗。
主要試劑:RNAisoTMPlus、RNase-free DNase I、M-MLV Reverse Transcriptase購于TaKaRa試劑公司;SYBR Premix Ex Taq購于TOYOBO試劑公司;2×Taq PCR MasterMix購于TIANGEN試劑公司;T7 RiboMAXTMExpress RNAi System購于Promega試劑公司。
1.2 飛蝗海藻糖酶基因及其編碼蛋白質(zhì)特性分析
1.2.1 飛蝗海藻糖酶基因cDNA序列搜索及特性分析 搜索筆者課題組構(gòu)建的飛蝗轉(zhuǎn)錄組數(shù)據(jù)庫獲得3個海藻糖酶基因cDNA全長序列和1個cDNA片段,將部分cDNA片段輸入飛蝗基因組數(shù)據(jù)庫中獲得其全長序列。使用ExPaSy網(wǎng)站上的相關(guān)軟件計算分子量及等電點。運用TMHMM和SignalP軟件預(yù)測4個海藻糖酶是否具有跨膜域和信號肽。
1.2.2 飛蝗海藻糖酶系統(tǒng)發(fā)育分析 使用GENEDOC軟件對飛蝗4個海藻糖基因氨基酸序列進行多位點比對,并標示出海藻糖酶標簽序列PGGRFrEyYyWDtY和qWDyPNaWpP及甘氨酸富集區(qū)GGGGEY;利用ClustalW軟件對NCBI已登錄的昆蟲以及部分細菌海藻糖酶基因全長氨基酸序列全長進行比對(表1)。使用MEGA7軟件中的NJ法構(gòu)建系統(tǒng)進化樹。
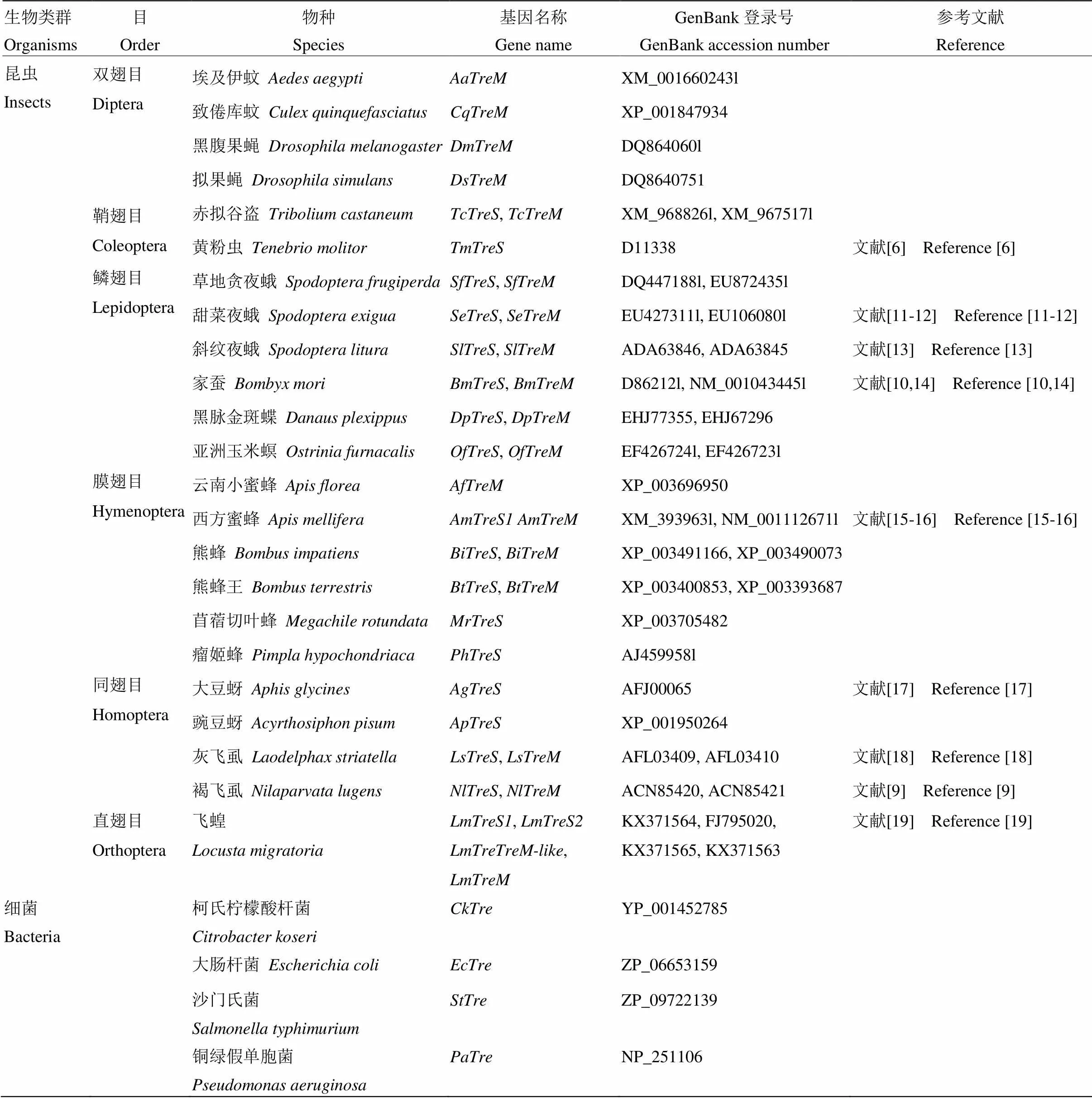
表1 用于構(gòu)建海藻糖酶系統(tǒng)進化樹的生物類群及海藻糖酶基因的GenBank登錄號
1.3 5齡飛蝗若蟲不同組織及發(fā)育日齡海藻糖酶基因的mRNA表達
1.3.1 樣品的收集 解剖5齡第2天飛蝗若蟲的體壁、前腸、中腸、后腸、胃盲囊、馬氏管、脂肪體、肌肉和氣管,進行不同組織部位基因的表達分析;選擇5齡第1天至第7天若蟲的肌肉、馬氏管、中后腸、體壁以及整蟲,進行不同發(fā)育日齡的基因表達分析。以上所有樣品均取3個生物學(xué)重復(fù),每個生物學(xué)重復(fù)至少3頭若蟲。所有樣品在體視顯微鏡下解剖后速凍于液氮中,-80℃保存?zhèn)溆谩?/p>
1.3.2 RT-qPCR表達引物的設(shè)計 根據(jù)飛蝗4個海藻糖酶基因和幾丁質(zhì)合成關(guān)鍵基因(GenBank accession number JX484802)和(GenBank accession number GU067730)的核苷酸序列,以為內(nèi)參基因,運用Primer 3.0軟件設(shè)計特異性的表達引物,引物由上海英濰捷基公司合成,具體的引物信息見表2。
1.3.3 總RNA提取、cDNA合成及RT-qPCR RNAisoTMPlus提取上述樣品總RNA,RNase free-DNAase I去除所提取總RNA中的DNA,1.5%的瓊脂糖凝膠電泳檢測RNA的質(zhì)量,用酶標儀進行定量,按照M-MLV Reverse Transcriptase說明書將1.5 μg的總RNA合成cDNA。在Applied Biosystems 7300 Real-Time PCR system(Applied Biosystems,USA)上進行RT-qPCR,PCR反應(yīng)體系按照SYBR Premix Ex Taq說明書進行配置,每個樣品3次技術(shù)重復(fù),并進行熔解曲線分析以保證擴增條帶的特異性。
1.3.4 數(shù)據(jù)處理與分析 分別將不同組織部位或發(fā)育日齡中最低表達量的數(shù)值視為1,其他組織部位或發(fā)育日齡的表達量以相對于最低表達量的倍數(shù)進行分析。試驗結(jié)果以平均數(shù)±標準差表示。采用SPSS中的Tukey’s HSD對試驗結(jié)果進行差異顯著性分析,柱形圖上不同字母代表基因表達差異顯著(<0.05)。
1.4 飛蝗海藻糖酶基因生物學(xué)功能分析
1.4.1 飛蝗海藻糖酶基因dsRNA引物的設(shè)計及體外合成 在E-RNAi網(wǎng)站設(shè)計飛蝗4個海藻糖酶基因和的dsRNA引物,引物信息見表2。分別以含有、、和的質(zhì)粒DNA為模板,通過PCR反應(yīng)獲得用于體外轉(zhuǎn)錄dsRNA的模板。PCR反應(yīng)體系及反應(yīng)程序按照2×Taq PCR MasterMix說明書進行。1.5%瓊脂糖凝膠電泳檢測PCR產(chǎn)物的單一性,按照Gel Extraction Kit說明書對DNA產(chǎn)物進行回收純化。以上述純化產(chǎn)物為模板,參照T7 RiboMAXTMExpress RNAi System試劑盒說明書進行dsRNA合成,酶標儀 上進行dsRNA定量至終濃度為2 μg·μL-1,保存至-80℃?zhèn)溆谩?/p>
1.4.2 dsRNA的注射 分別選取5齡第2天飛蝗若蟲進行、、的dsRNA注射,同時注射ds作為對照組。采用微量注射器(寧波三愛儀器廠)分別將上述dsRNA從飛蝗第2和3腹節(jié)連接處注入體腔,注射量為每頭10 μg,每個基因設(shè)置3個生物學(xué)重復(fù),每個重復(fù)30頭若蟲。注射完畢后,將所有試蟲放置于人工氣候箱中飼養(yǎng)。
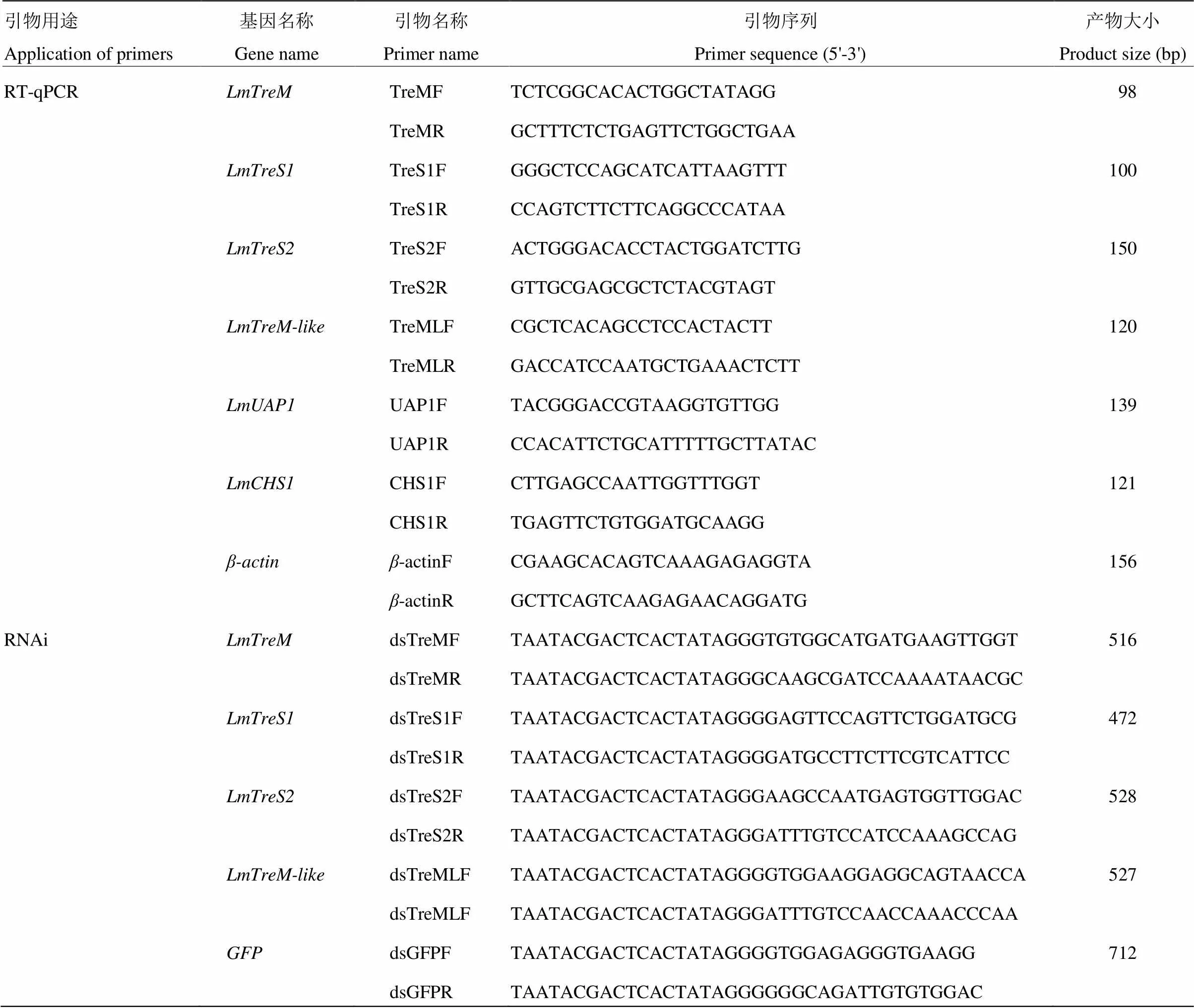
表2 所用RT-qPCR和RNAi的引物信息
1.4.3 沉默效率檢測及表型觀察 分別收集注射上述基因dsRNA 48 h后的整蟲樣本,提取RNA,反轉(zhuǎn)錄為cDNA,采用RT-qPCR方法對海藻糖酶基因的沉默效果和UDP--乙酰葡糖胺焦磷酸化酶1基因()、幾丁質(zhì)合成酶1基因()的表達進行檢測,3個生物學(xué)重復(fù),每組用蟲3頭,具體操作與1.3.4的檢測方法相同,其余試蟲用來觀察表型。
2 結(jié)果
2.1 飛蝗海藻糖酶基因及其編碼蛋白質(zhì)特性分析
將搜索獲得的4個海藻糖酶基因cDNA全長序列進行blast分析,結(jié)果顯示4個海藻糖酶均具有海藻糖酶特有的2個標簽序列PGGRFrEyYyWDtY和qWDyPNaWpP以及1個甘氨酸富集區(qū)GGGGEY(圖1)。使用ExPaSy網(wǎng)站上的相關(guān)軟件計算分子量及等電點,表明飛蝗4個海藻糖酶的相對分子量在62.8—69.9 kD,其pI均為酸性(表3)。TMHMM分析表明1個海藻糖酶具有典型的跨膜結(jié)構(gòu),命名為膜結(jié)合型海藻糖酶LmTreM,2個海藻糖酶沒有跨膜結(jié)構(gòu),分別將其命名為可溶性海藻糖酶LmTreS1和LmTreS2,1個海藻糖酶存在類跨膜結(jié)構(gòu),軟件分析結(jié)果顯示其概率為0.59(概率>0.6即可預(yù)測為跨膜結(jié)構(gòu)),故暫將其命名為類膜結(jié)合型海藻糖酶LmTreM-like。SignalP預(yù)測發(fā)現(xiàn)只有LmTreM-like具有信號肽。GENEDOC分析表明4個海藻糖酶氨基酸的一致度在47%—65%,其中LmTreM和LmTreM-like氨基酸的一致度最高達65%(表4)。

單下劃線標示為海藻糖酶的兩個標簽序列;雙下劃線標示為海藻糖酶的甘氨酸富集區(qū);箭頭所示為LmTre2的信號肽;方框所示為LmTreM的跨膜結(jié)構(gòu)域Trehalase two signature regions and the glycine-rich region were indicated by the underline and double underline. The arrow represented the signal peptide in LmTre2. Putative transmembrane region was boxed in LmTreM

表3 飛蝗海藻糖酶基因編碼蛋白的特性
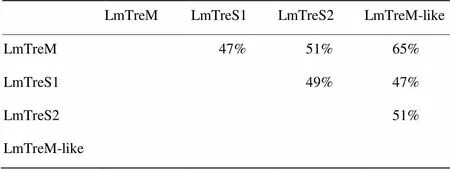
表4 飛蝗4個海藻糖酶基因氨基酸一致度分析
2.2 海藻糖酶系統(tǒng)進化樹分析
根據(jù)已知昆蟲和部分細菌海藻糖酶基因氨基酸全長序列構(gòu)建系統(tǒng)進化樹(圖2),結(jié)果顯示LmTreM和LmTreM-like與其他昆蟲膜結(jié)合型海藻糖酶優(yōu)先聚在一支;LmTreS1和LmTreS2與大豆蚜、豌豆蚜、褐飛虱、灰飛虱等不完全變態(tài)昆蟲可溶性海藻糖酶聚為一支,表明其親緣關(guān)系較近,而與全變態(tài)昆蟲可溶型海藻糖酶相距較遠。
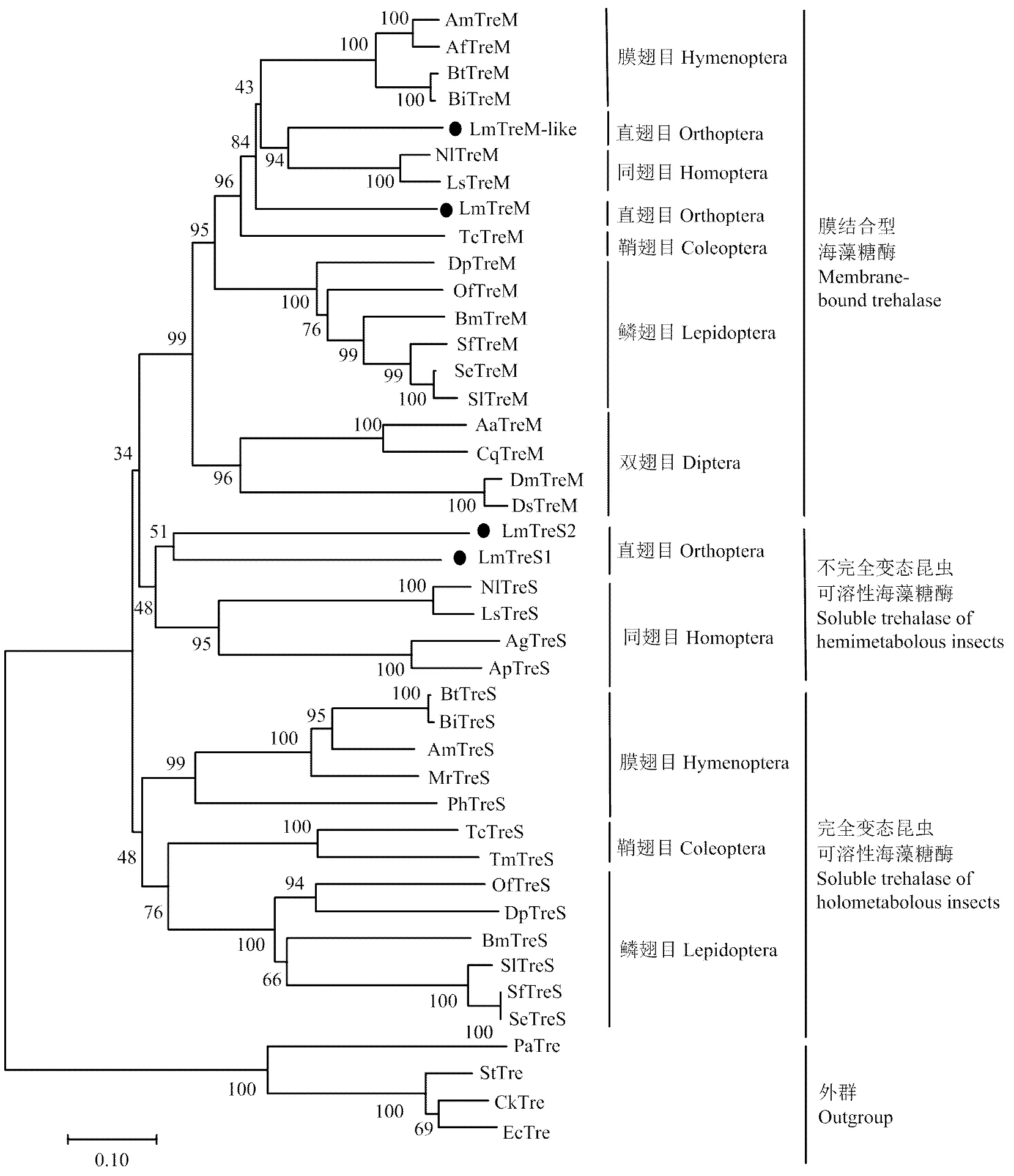
圖2 根據(jù)已知昆蟲和部分細菌海藻糖酶氨基酸序列構(gòu)建的系統(tǒng)發(fā)育樹
2.3 飛蝗4個海藻糖酶基因的表達
2.3.1 在不同組織部位的表達 飛蝗在肌肉中特異性表達,而在其他組織中的表達較低;在馬氏管表達最高,其次是中腸和后腸,在其他組織部位表達較低;主要在體壁中表達,其次是脂肪體、肌肉和氣管;在不同組織部位中的表達相對均衡(圖3)。
2.3.2 在不同發(fā)育時期表達 由不同組織表達結(jié)果可知和分別在肌肉、馬氏管和中后腸、體壁()中高表達,而在各組織的表達相對均衡,因此解剖5齡若蟲每一天的上述組織或整蟲提取RNA進行RT-qPCR分析。結(jié)果表明在5齡1—2 d表達較低,3—6 d表達比較高,到第7天表達又有所下降;在在馬氏管和中后腸的表達量隨日齡增加逐漸上升,第3—7天保持穩(wěn)定高表達;在1—3 d表達量比較高,之后表達量逐漸降低;在5齡發(fā)育過程中穩(wěn)定表達,沒有顯著差異(圖4)。
2.4 飛蝗海藻糖酶基因生物學(xué)功能
分別將、、與的dsRNA注射至5齡第2天的飛蝗若蟲,48 h后收集整蟲樣品提取RNA,利用RT-qPCR技術(shù)檢測4個海藻糖酶基因的沉默效率和幾丁質(zhì)合成關(guān)鍵基因和的表達,結(jié)果表明各基因的表達均被有效沉默,且無交叉干擾現(xiàn)象,和的表達也無顯著變化,注射各基因的dsRNA后5齡若蟲均可順利蛻皮至成蟲(圖5、圖6)。
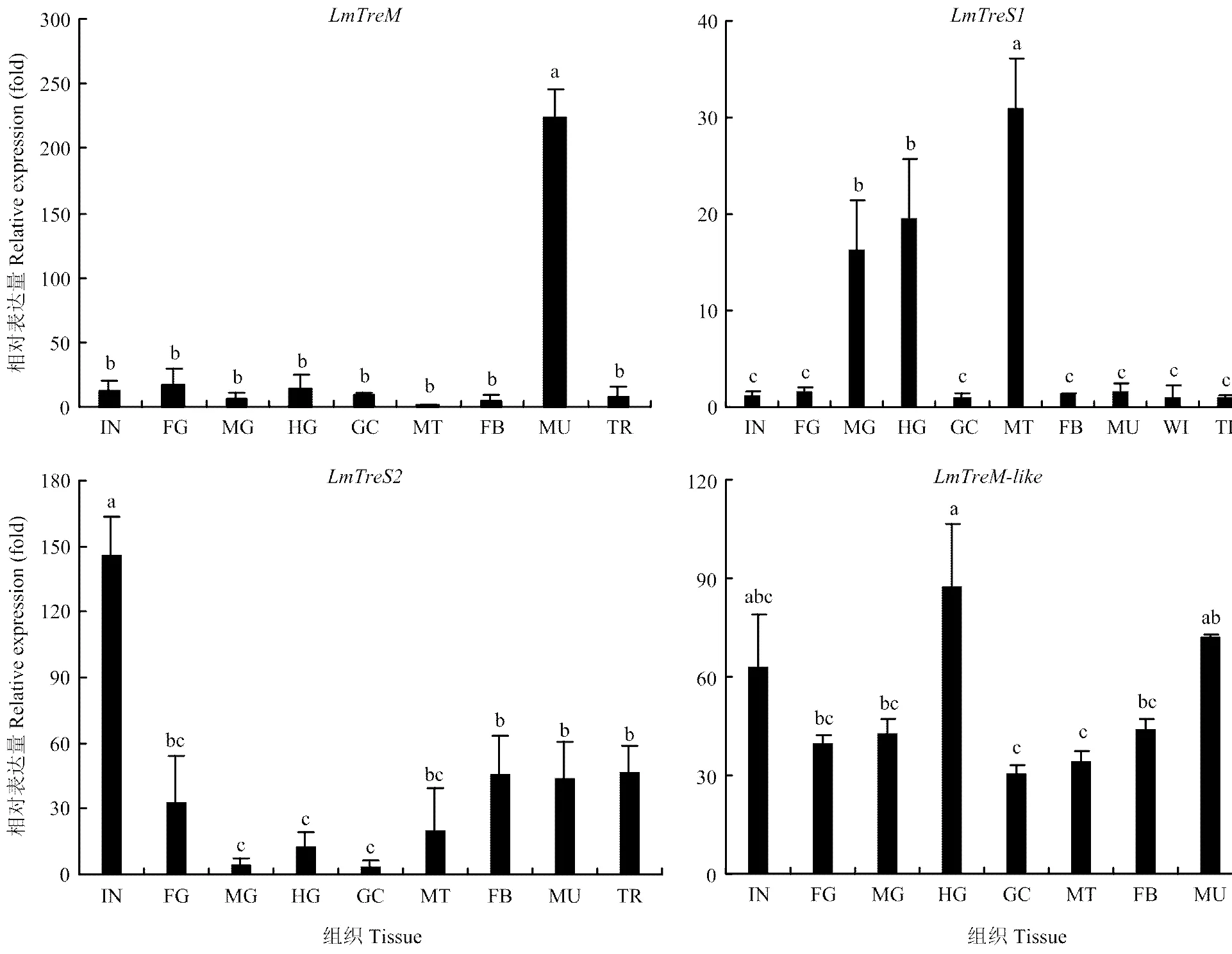
IN:體壁 Integument;FG:前腸 Foregut;MG:中腸 Midgut;HG:后腸 Hindgut;GC:胃盲囊 Gastric caeca;MT:馬氏管 Malpighian tubules;FB:脂肪體 Fat body;MU:肌肉 Muscle;TR:氣管 Trachea。圖中數(shù)值為平均數(shù)±標準差,柱形圖上不同字母代表基因表達差異顯著(P<0.05, Tukey’s HSD檢驗)。下同Each value represents mean±SD, different letters indicated significant differences (P<0.05, Tukey’s HSD test). The same as below
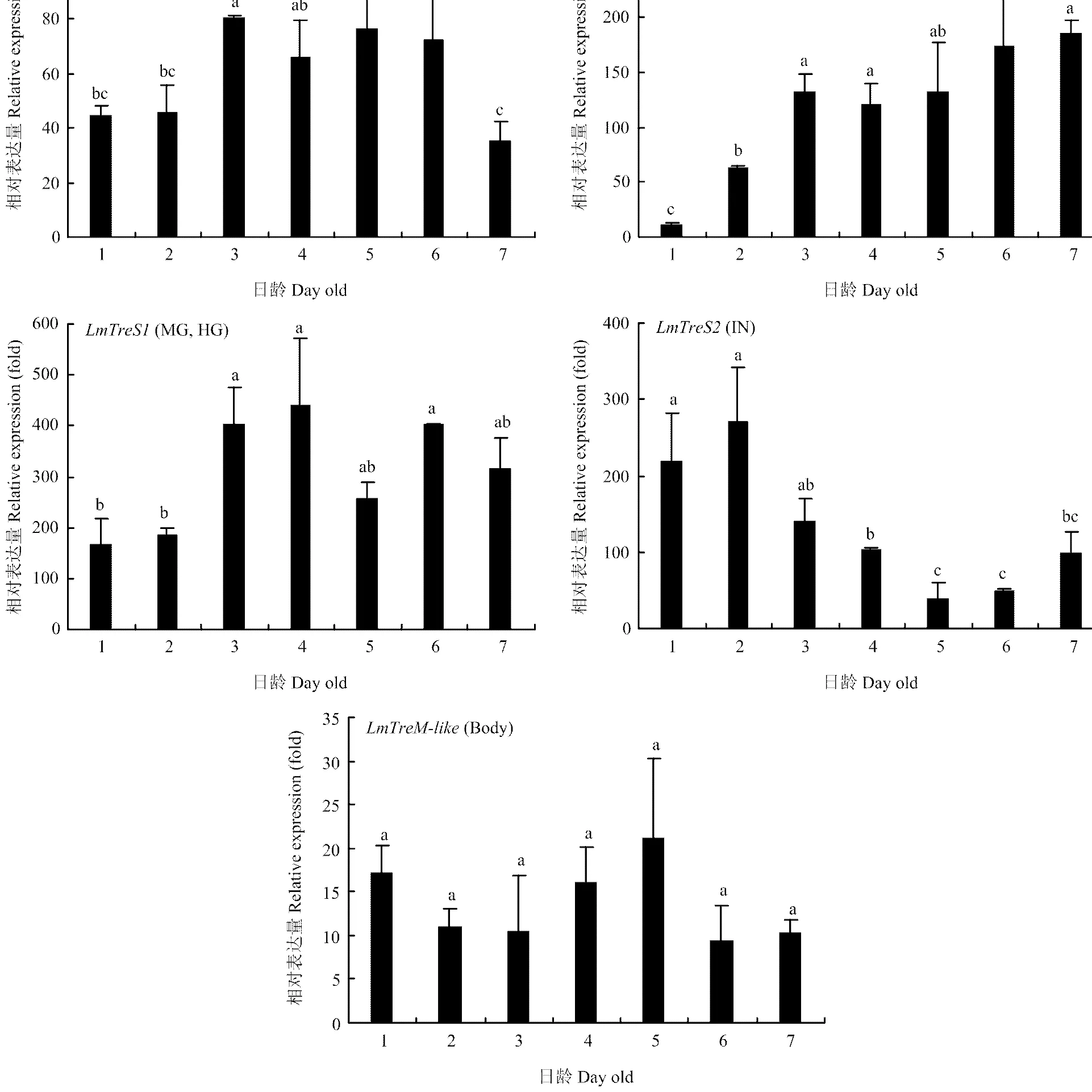
MU:肌肉 Muscle;MT:馬氏管 Malpighian tubules;MG:中腸 Midgut;HG:后腸 Hindgut;IN:體壁 Integument;Body:整蟲
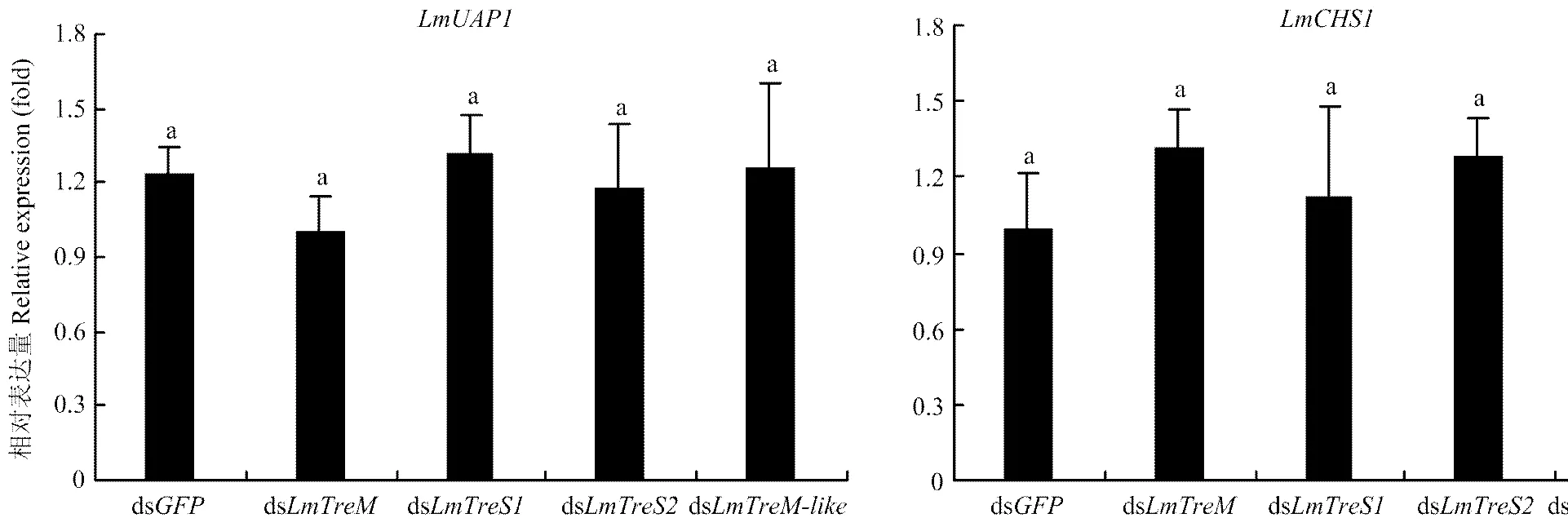
圖6 注射海藻糖酶基因dsRNA后對幾丁質(zhì)合成關(guān)鍵基因LmUAP1和LmCHS1表達量的影響
3 討論
根據(jù)是否具有跨膜結(jié)構(gòu)可將昆蟲海藻糖酶分為膜結(jié)合型和可溶性海藻糖酶兩類[20]。筆者課題組之前已報道飛蝗1個可溶性海藻糖酶基因的分子特性[19],本研究發(fā)現(xiàn)飛蝗還存在另外3個海藻糖酶基因,分別命名為LmTreM、LmTreS1、LmTreS2和LmTreM-like,LmTreS2即為之前報道的海藻糖酶。LmTreM-like與LmTreM氨基酸的一致度最高達65%,而與LmTreS1和LmTreS2的氨基酸一致度僅為47%和51%。然而運用TMHMM軟件分析表明該酶僅具有類跨膜域結(jié)構(gòu),概率為0.59(概率>0.6即可表明有典型的跨膜結(jié)構(gòu)),推測LmTreM-like末端存在的類跨膜結(jié)構(gòu)使其與膜結(jié)合型海藻糖酶具有較近的親緣關(guān)系。
本研究采用RT-qPCR技術(shù)對4個海藻糖酶基因在5齡飛蝗不同組織和發(fā)育日齡的表達進行了分析,結(jié)果表明4個海藻糖酶基因在不同組織部位的表達差異顯著。在肌肉中特異性表達,并且在5齡蛻皮前后表達較低而齡期中間表達較高,可能與飛蝗在蛻皮前后運動減少而生長發(fā)育階段運動較活躍的生理現(xiàn)象相關(guān),Wegener等[21]發(fā)現(xiàn)飛蝗肌肉中膜結(jié)合型海藻糖活性占比達95%,注射海藻糖酶抑制劑后能夠影響飛蝗的活動能力并減少取食,這與本文中主要在肌肉表達相符,推測膜結(jié)合型海藻糖酶主要為昆蟲的飛行運動提供能量。主要在馬氏管和中后腸表達,馬氏管是昆蟲排泄和滲透調(diào)節(jié)的主要器官,而中后腸為主要的消化器官,推測該基因主要參與消化代謝及廢物排泄;主要在體壁中表達,且在蛻皮后的1—3 d內(nèi)表達量相對較高,之后表達量逐漸降低[19],這與飛蝗幾丁質(zhì)合成關(guān)鍵基因UDP--乙酰氨基葡萄糖焦磷酸化酶1基因()[22]和幾丁質(zhì)合成酶1基因()的變化規(guī)律相似[23]。筆者課題組已利用RNAi技術(shù)證實和均參與了飛蝗表皮幾丁質(zhì)合成,因此推測可能與飛蝗新表皮幾丁質(zhì)合成需要大量海藻糖酶有關(guān)。這與在甜菜夜蛾中發(fā)現(xiàn)可溶性海藻糖酶基因主要在體壁和馬氏管中高表達結(jié)果相一致,利用RNAi技術(shù)發(fā)現(xiàn)該基因表達沉默后,主要影響昆蟲表皮中幾丁質(zhì)的合成[11-12]。而飛蝗在各個組織部位和發(fā)育時期中的分布則比較均衡。
本研究采用RNAi技術(shù)研究飛蝗4個海藻糖酶基因的生物學(xué)功能,分別將4個海藻糖酶基因dsRNA注射至5齡若蟲后,發(fā)現(xiàn)沉默效率均在60%以上,4個基因之間無交叉干擾,但任一基因的表達沉默后并未影響幾丁質(zhì)合成關(guān)鍵基因和的表達,5齡飛蝗可正常蛻皮至成蟲。海藻糖酶是昆蟲能量代謝必不可少的一個酶,受蛻皮激素20E的調(diào)控,如在綠盲蝽()中用幾丁質(zhì)合成抑制劑氟鈴脲處理昆蟲或注射ds后可溶性海藻糖酶的表達及酶活性顯著降低,造成昆蟲發(fā)育遲緩及體重降低等表型[24-25]。海藻糖酶也是幾丁質(zhì)合成過程中第一個酶, Zhao等利用RNAi技術(shù)闡明了甜菜夜蛾和褐飛虱中海藻糖酶基因在幾丁質(zhì)合成過程中的功能,結(jié)果表明分別抑制各個海藻糖酶基因的表達后,幾丁質(zhì)合成酶等基因也相應(yīng)下調(diào),最終導(dǎo)致昆蟲死亡[9,11]。本研究結(jié)果與甜菜夜蛾及褐飛虱等昆蟲的結(jié)果有區(qū)別,表明不同昆蟲之間存在差異。飛蝗體內(nèi)有4個海藻糖酶基因,推測沉默其中任一個基因后,其他海藻糖酶基因能夠彌補這個海藻糖酶基因的缺失,因此單獨干擾某一基因并不影響5齡飛蝗正常蛻皮至成蟲。今后將同時干擾4個基因的表達,觀察飛蝗生長發(fā)育情況,以深入揭示飛蝗海藻糖酶基因的分子功能。
4 結(jié)論
飛蝗具有1個膜結(jié)合型、1個類膜結(jié)合型和2個可溶性海藻糖酶基因,這4個海藻糖酶基因具有不同的組織和發(fā)育表達特性,任一基因的表達下調(diào)并不影響5齡飛蝗蛻皮至成蟲。
References
[1] Yang M L, Zhang J Z, Zhu K Y, Xuan T, Liu X J, Guo Y P, Ma E B. Mechanisms of organophosphate resistance in a field population of oriental migratory locust,(Meyen)., 2009, 71(1): 3-15.
[2] Elbein A D, Pan Y T, Pastuszak I, Carroll D. New insights on trehalose: a multifunctional molecule., 2003, 13(4): 17-27.
[3] Merzendorfer H, Zimoch L. Chitin metabolism in insects: structure, function and regulation of chitin synthases and chitinases., 2003, 206(24): 4393-4412.
[4] 唐斌, 魏蘋, 陳潔, 王世貴, 張文慶. 昆蟲海藻糖酶的基因特性及功能研究進展. 昆蟲學(xué)報, 2012, 55(11): 1315-1321.
TANG B, WEI P, CHEN J, WANG S G, ZHANG W Q. Progress in gene features and functions of insect trehalases., 2012, 55(11): 1315-1321. (in Chinese)
[5] Shukla E, Thorat L J, Nath B B, Gaikwad S M. Insect trehalase: physiological significance and potential applications., 2015, 25(4): 357-367.
[6] Takiguchi M, Niimi T, Su Z H, Yaginuma T. Trehalase from male accessory gland of an insect,. cDNA sequencing and developmental profile of the gene expression., 1992, 288(1): 19-22.
[7] Mitsumasu K, Azuma M, Niimi T, Yamashita O, Yaginuma T. Membrane-penetrating trehalase from silkworm. Molecular cloning and localization in larval midgut., 2005, 14(5): 501-508.
[8] Shi Z K, Liu X J, Xu Q Y, Qi Z, Wang S, Zhang F, Wang S G, Tang B. Two novel soluble trehalase genes cloned fromand regulation of the enzyme in a rapid changing temperature.:, 2016, 198: 10-18.
[9] Zhao L, Yang M, Shen Q, Liu X, Shi Z, Wang S, Tang B. Functional characterization of three trehalase genes regulating the chitin metabolism pathway in rice brown planthopper using RNA interference., 2016, 6: Article number 27841.
[10] Mitsumasu K, Kanamori Y, Fujita M, Iwata K , Tanaka D, Kikuta S, Watanabe M Cornette R, Okuda T, Kikawada T. Enzymatic control of anhydrobiosis-related accumulation of trehalase in the sleeping chironomid,.,2010, 277(20): 4215-4228.
[11] Chen J, Tang B, Chen H, Yao Q, Huang X, Chen J, Zhang D, Zhang W. Different functions of the insect soluble and membrane-bound trehalase genes in chitin biosynthesis revealed by RNA interference., 2010, 5(4): e10133.
[12] Tang B, Chen X, Liu Y, Tian H, Liu J, HU J, XU W, ZHANG W. Characterization and expression patterns of a membrane-bound trehalase from., 2008, 9: 51.
[13] Zou Q, Wei P, Xu Q, Zheng H Z, Tang B, Wang S G. cDNA cloning and characterization of two trehalases from(Lepidoptera; Noctuidade)., 2013, 12(2): 901-915.
[14] Su Z H, Sato Y, Yamashita O. Purification, cDNA cloning and northern blot analysis of trehalase of pupal midgut of the silkworm,, 1993, 1173(2): 217-224.
[15] Lee J H, Saito S, Mori H, Nishimoto M, Okuyama M, Kim D, Wongchawalit J, Kimura A, Chiba S. Molecular cloning of cDNA for trehalase from the European honeybee,L, and its heterologous expression in., 2007, 71(9): 2256-2265.
[16] Mori H, Lee J H, Okuyama M, Nishimoto M, Ohguchi M, Kim D, Kimura A, Chiba S.Catalytic reaction mechanism based on alpha-secondary deuterium isotope effects in hydrolysis of trehalose by European honeybee trehalase., 2009, 73(11): 2466-2473.
[17] Bansal R, Mian M A, Mittapalli O, Michel A P. Molecular characterization and expression analysis of soluble trehalase gene in, a migratory pest of soybean., 2013, 103(3): 286-295.
[18] 張倩, 魯鼎浩, 蒲建, 吳敏, 韓召軍. 灰飛虱海藻糖酶基因的克隆及RNA干擾效應(yīng). 昆蟲學(xué)報, 2012, 55(8): 911-920.
ZHANG Q, LU D H, PU J, WU M, HAN Z J. Cloning and RNA interference effects of trehalase genes in, 2012, 55(8): 911-920. (in Chinese)
[19] 劉曉健, 張歡歡, 李大琪, 崔淼, 馬恩波, 張建珍. 飛蝗可溶型海藻糖酶基因的序列分析及mRNA表達特性. 昆蟲學(xué)報, 2012, 55(11): 1264-1271.
LIU X J, ZHANG H H, LI D Q, CUI M, MA E B, ZHANG J Z. Sequence characterization and mRNA expression profiling of a soluble trehalase gene in(Orthoptera: Acrididae)., 2012, 55(11): 1264-1271. (in Chinese)
[20] Becker A, Schl?der P, Steele J E, Wegener G. The regulation of trehalose metabolism in insects., 1996, 52: 433-439.
[21] Wegener G, Macho C, Schl?der P, Kamp G, Ando O. Long-term effects of the trehalase inhibitor trehazolin on trehalase activity in locust flight muscle.,2010, 213(22): 3852-3857.
[22] Liu X J, Li F, Li D Q, Ma E B, Zhang W Q, Zhu K Y, Zhang J Z. Molecular and functional analysis of UDP--acetylglucosamine pyrophosphorylases from the migratory locust,., 2013, 8(8): e71970.
[23] Zhang J Z, Liu X J, Zhang J Q, Li D Q, Sun Y, Guo Y P, Ma E B, Zhu K Y. Silencing of two alternative splicing-derived mRNA variants of chitin synthase 1 gene by RNAi is lethal to the oriental migratory locus(Meyen)., 2010, 40(11): 824-833.
[24] Tan Y A, Xiao L B, Sun Y, Zhao J, Bai L X. Sublethal effects of the chitin synthesis inhibitor, hexaflumuron, in the cotton mirid bug,(Meyer-Dür).,2014, 111: 43-50.
[25] Tan Y A, Xiao L B, Zhao J, Xiao Y F, Sun Y, Bai L X. Ecdysone receptor isoform-B mediates soluble trehalase expression to regulate growth and development in the mirid bug,(Meyer-Dür)., 2015, 24(6): 611-623.
(責(zé)任編輯 岳梅)
Molecular Characteristics and Functional analysis ofGenes in
LIU Xiao-jian1, SUN Ya-wen1,2, CUI Miao1, MA En-bo1, ZHANG Jian-zhen1
(1Research Institute of Applied Biology, Shanxi University, Taiyuan 030006;2College of Life Science, Shanxi University, Taiyuan 030006)
【Objective】 Trehalase, the only enzyme that hydrolyzes one trehalose molecule into two glucose molecules, plays key roles in insect energy metabolism and chitin synthesis. So trehalase could be served as a potential target for insect pest control. In this paper, the molecular characteristics and functions ofgenes were explored in an important agricultural pest. The results will provide a reasonable basis for the effective management of locusts. 【Method】By searching the transcriptome and genome of, four full-length cDNA ofgenes were obtained. Sequence characteristics of these four trehalases were analyzed by using blast, TMHMM and SignalP softwares. Multiple amino acid sequence alignment of trehalases was made using ClustalW. The phylogenetic tree was constructed by MEGA 7. The expression patterns ofgenes in different tissues and developmental days were studied in the 5th-instar nymphs by RT-qPCR. The dsRNAs of fourgenes were synthesized, and then injected into the 5th-instar nymphs on day 2, respectively. Control nymphs were injected with dsalone. The whole body after the dsRNA injection for 48 h was used for total RNA extraction and cDNA synthesis. RT-qPCR were performed to determine the transcript levels of fourgenesand genes involved in chitin synthesis such as UDP--acetylglucosamine pyrophosphorylase geneandchitin synthase 1 gene. The abnormal nymphs displayed phenotypes were carefully observed. 【Result】Blast analysis showed that all these four trehalases contained two conservative regions and a glycine enrichment region. A membrane-bound trehalase with a typical transmembrane domain was named LmTreM (GenBank accession number KX371563). A transmembrane like domain was predicted in another trehalase, named LmTreM-like (GenBank accession number KX371565). The two remains were soluble trehalases, named LmTreS1 and LmTreS2, respectively (GenBank accession number KX371564 and FJ795020). Phylogenetic analysis showed that LmTreM and LmTreM-like, LmTreS1 and LmTreS2 were clustered with membrane-bound and soluble trehalases, respectively. RT-qPCR was carried out to analyze the expression patterns of fourgenes in different tissues and developmental days in the 5th-instar nymphs of. The results indicated,andwere mainly expressed in specific tissues, andwas consistently expressed in all selected tissues. mRNA transcripts of fourgenes were different during the development of 5th-instar nymphs. RNAi results suggested that expressions of fourgenes were significantly reduced compared to the control, and the injection of,,anddsRNA only repressed the expression of the targetedwithout reducing that of other, the expressions ofandinvolved in chitin synthesis were not affected. However, nymphs could successfully molt to adults after injected with fourdsRNAs, respectively.【Conclusion】One membrane-bound, one membrane-bound like and two soluble trehalase were found in. The fourgenes showed different tissue and developmental expression characteristics, silencing of each of these fourgenes in the 5th-instar nymphs could not affect the normal molting to adults.
; trehalase gene; expression characteristics; RT-qPCR; RNA interference
2016-07-01;接受日期:2016-09-21
國家自然科學(xué)基金(31402020,31272380)、山西省高校科技創(chuàng)新(2016113)、山西省青年基金(201601D021120)、山西省科技基礎(chǔ)條件平臺建設(shè)(2015091010)
劉曉健,E-mail:lxiaoljian@163.com。通信作者張建珍,Tel:0351-7018871;E-mail:zjz@sxu.edu.cn
