游仆蟲(原生動物,纖毛門)皮層銀線系的形態發生模式?
2016-12-24 01:58:33謝冬梅范鑫鵬顧福康
中國海洋大學學報(自然科學版) 2016年12期
謝冬梅, 范鑫鵬, 倪 兵, 顧福康
(華東師范大學生命科學學院,上海 200241)
?
游仆蟲(原生動物,纖毛門)皮層銀線系的形態發生模式?
謝冬梅, 范鑫鵬??, 倪 兵, 顧福康
(華東師范大學生命科學學院,上海 200241)
有關研究證實,游仆類的銀線系在超微結構水平上對應了由微管束構成的網絡骨架結構,關于其在纖毛蟲細胞分裂過程中的發生模式鮮有報道。本工作應用銀浸染色法和掃描電鏡術,以扇形游仆蟲(Euplotesvannus)、鐮游仆蟲(Euplotesharpa)和包囊游仆蟲(Euplotesencysticus)為代表,研究了游仆類纖毛蟲3種類型皮層銀線系的形態發生過程。研究表明,游仆蟲3種類型銀線系的形態發生過程呈現基本相同的模式:該3種纖毛蟲新銀線網在棘毛原基基體周圍發生,據此推測口圍帶小膜、棘毛基體復合單元和背觸毛基體單元不僅對相應纖毛器結構行使微管組織中心作用,也可能對相應皮層區銀線網的發生具有定位和組織中心的作用;圍繞在基體左側的銀線網先于右側開始發育,據此推測銀線系的分化可能也具有方向性;腹面銀線系發生中離新結構發生區近的老結構先瓦解,由此使新結構不斷延伸發展,據此推測老銀線網對新結構的形成可能起到誘導及定向作用;背面銀線系發生中在每一背觸毛列的中部范圍相鄰2列新背觸毛間的老銀線網消失,背觸毛左、右兩側的新銀線網會合形成新銀線網,并且前仔蟲前半部分和后仔蟲后半部分的老銀線網保留下來共同組成新細胞的背面銀線網,由此表明,其老銀線網參與了新結構的組成并對新結構的形成具有物質貢獻。
游仆蟲;銀線系;形態發生;銀浸染色法;掃描電鏡術
應用銀浸染色法(Silver impregnation technique),能使膜口類、前口類、腎形類和游仆類纖毛蟲的細胞皮層顯示一種網狀或條紋狀結構,這種結構被稱為銀線系(Silver-line system)[1]。其中,游仆蟲屬于纖毛蟲中最高等的類群,研究其皮層銀線系的形態特征及其分化對揭示纖毛蟲皮層結構模式具有重要意義。自60多年前Tuffrau發現游仆蟲細胞表面具有銀線系以來,對游仆蟲銀線系在細胞非分裂時期形態的觀察中已經積累了較豐富的資料[2-8]。Curds[9]根據游仆蟲屬不同種類細胞背皮層銀線系網格的組成特征,將游仆蟲屬的背銀線系分為5種基本類型,即:相鄰2列背觸毛間含有1列銀線網的單扇形型銀線系(Single-vannussilver-line system);相鄰2列背觸毛間含有2列大小近似相等的銀線網的雙闊口型銀線系(Double-eurystomussilver-line system);相鄰2列背觸毛間含有2列大小不等的銀線網的雙盤狀型銀線系(Double-patellatype);相鄰2列背觸毛間含有多列排列規則的銀線網的多線型銀線系(Multiple silver-line system);相鄰2列背觸毛間含有多列排列不規則的銀線網的復雜型銀線系(Complex silver-line system)。根據相鄰2列背觸毛之間銀線網的列數,這5種銀線系可概括為3種主要類型:單線型、雙線型和多線型[10]。近年來,所在實驗室通過用生化抽提結合掃描電鏡術,以及應用熒光紫杉醇標記法顯示,游仆蟲背、腹面網狀纖維骨架與銀浸法獲得的銀線系是同一種結構,并且也是一類重要的皮層微管骨架[11-13]。但是,目前對游仆蟲這類細胞表面骨架僅限于基本形態的描述,尚未深入到其形態發生及其結構形成機理的研究。因此,作者應用銀浸染色法及掃描電鏡技術,以代表單線型銀線系的扇形游仆蟲(Euplotesvannus)、代表雙線型銀線系的鐮游仆蟲(E.harpa)和代表多線型銀線系的包囊游仆蟲(E.encysticus)3種游仆蟲為材料,對游仆蟲皮層銀線系的形態及形態發生進行了比較觀察,以期為揭示該類纖毛蟲生命活動中細胞結構的分化及其調控機理提供新的資料。
1 材料和方法
1.1 材料
鐮游仆蟲采自上海市崇明島北湖,扇形游仆蟲和包囊游仆蟲來自中國海洋大學原生動物學研究室。將3種游仆蟲在室溫下用表面皿建立純系培養,扇形游仆蟲以煮沸后鹽度為30的海水為培養液,其余2種以煮沸過濾后的湖水(鹽度0)為培養液,每日喂食3次淡水草履唇滴蟲(Chilomonasparamecium),不同鹽度的游仆蟲均能夠以淡水草履唇滴蟲為食。一般在每次喂食后數小時內能獲得大量的分裂細胞。
1.2 方法
顯示細胞表面銀線系及纖毛器采用Chatton-Lwoff銀浸染色技術[14]并稍作變更:固定液用3%重鉻酸鉀、1%三氧化鉻、2%鋨酸按7∶7∶5(體積比)相混合,現配現用。固定10 min后轉入Da Fano液中再固定。
掃描電鏡樣品制備主要步驟如下:將收集的蟲體在6份飽和升汞溶液和1份1%的鋨酸相混合的固定液中固定10 min;用超純水清洗,除去固定液;梯度酒精脫水;CO2臨界點干燥;將干燥后的樣品轉移至樣品銅臺上;噴金;用掃描電子顯微鏡(Hitachi S-4800)觀察和拍照[15]。
2 結果
2.1 細胞分裂間期3種游仆蟲皮層銀線系的形態
2.1.1 細胞分裂間期扇形游仆蟲皮層銀線系的形態 扇形游仆蟲的細胞腹皮層纖毛器包括位于左前側的口圍帶、波動膜,以及按10-5-2-2或10-5-3-2模式分布的額腹棘毛(FC)、橫棘毛(TC)、尾棘毛(CC)和左緣棘毛(LMC)。在腹皮層表面布滿四邊形的或近似四邊形的銀線網。其中,在額腹棘毛與橫棘毛之間的區域含7~8列銀線網,其網格排列較整齊,從前到后每列銀線網約由9格緊密排列的四邊形網格構成;在口圍帶基部與波動膜基部交叉處的銀線網以及位于最右側橫棘毛右后方的伸縮泡孔周圍的銀線網較小,且網格呈緊密排列(見圖1A)。在細胞背面,背皮層纖毛器包括9~10列背觸毛(DK),整個背皮層表面布滿四邊形銀線網。在相鄰2列背觸毛間的銀線網均由1列組成,其中在中部范圍于每2列背觸毛間的銀線網約為14格,在其兩側每2列背觸毛間的銀線網漸少(見圖1B)。
2.1.2 細胞分裂間期鐮游仆蟲皮層銀線系的形態 鐮游仆蟲的細胞腹皮層纖毛器包括位于左前側的口圍帶、波動膜,以及按10-5-2-2模式分布的額腹棘毛、橫棘毛、尾棘毛和左緣棘毛。在腹皮層表面布滿多邊形銀線網。其中,位于口腔前方,口圍帶與波動膜之間的皮層表面布滿多邊形至六邊形的銀線網;在口圍帶基部與波動膜基部交叉處的銀線網以及位于最右側橫棘毛右后方的伸縮泡孔周圍的銀線網較小,且網格呈緊密排列(見圖1C)。在細胞背面,背皮層纖毛器包括13列背觸毛,整個背皮層表面布滿四邊形銀線網。在相鄰兩列背觸毛間的銀線網均由2列組成,其中在中部范圍于每2列背觸毛間的銀線網約為48格,在其兩側每2列背觸毛間的銀線網漸少(見圖1D)。
2.1.3 細胞分裂間期包囊游仆蟲皮層銀線系的形態 包囊游仆蟲的細胞腹皮層纖毛器包括位于左前側的口圍帶、波動膜,以及按9-5-2-2模式分布的額腹棘毛、橫棘毛、尾棘毛和左緣棘毛。在腹皮層表面布滿多邊形銀線網。其中,位于口腔前方,口圍帶與波動膜之間的皮層表面布滿多邊形至六邊形的銀線網;在口圍帶基部與波動膜基部交叉處的銀線網以及位于最右側橫棘毛右后方的伸縮泡孔周圍的銀線網較小,且網格呈緊密排列(見圖1E、G)。在細胞背面,背皮層纖毛器包括7列背觸毛,整個背皮層表面布滿多邊形銀線網。在相鄰2列背觸毛間的銀線網排列不規則,其中在中部范圍于每2列背觸毛間的銀線網約為71格,在其兩側每2列背觸毛間的銀線網漸少(見圖1F、H)。
2.2 分裂期3種游仆蟲皮層銀線系的形態發生
2.2.1 分裂期扇形游仆蟲皮層銀線系的形態發生 在扇形游仆蟲中,首先于口圍帶基部后端左側皮層產生成斜向“一”字形排列小群毛基體(見圖2A),毛基體快速增殖,形成后仔蟲口圍帶原基區。此時,在口原基區右前方的龕腔壁上出現緊密排列的新銀線網(見圖2B)。當口原基區自前端開始逐步組裝成半弧形排列的小膜時,新銀線網在其右側不斷擴展。伴隨著龕腔變大,口圍帶小膜充分發育變寬,其一側的新銀線網也充分擴展,成為處于后仔蟲口腔前方,在口圍帶和波動膜之間皮層表面的新銀線網(見圖2C)。
新口原基形成時,在額腹橫棘毛皮層區形成2組各含5列原基的前、后額腹橫棘毛原基區(見圖2D),其每列原基不斷變粗,并按由后至前、由左至右順序分化成3、3、3、3、2段棘毛原基(見圖2E、F)。與此同時,在每段棘毛原基的左側和右側按自后至前的順序發生細小的銀線網,其銀線網不斷發育伸展,于每列原基的左右兩側各形成1列新銀線網(見圖2G、H),最終在新的額腹橫棘毛原基區形成7~8列四邊形或近似四邊形的銀線網。此外,伴同老口側膜前方唇和新口圍帶原基右側皮層區的前、后2個新棘毛原基的分化,其周圍也發生細小的銀線網并不斷發育伸展,形成四邊形或近似四邊形的銀線網(見圖2E)。隨著額腹橫棘毛原基進一步發育形成新棘毛、蟲體演化成前、后仔蟲時,腹面殘留的老棘毛及老銀線網消失,新棘毛及新銀線網相繼定位,成為前、后仔蟲的額腹橫棘毛和腹面銀線網。
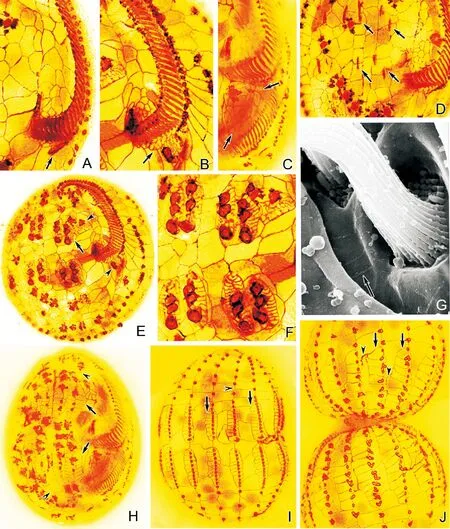
(A~F:銀浸染色照片;G,H:掃描電鏡照片;標尺:20 μm。A.扇形游仆蟲腹面含有排列較整齊的四邊形或近四邊形銀線網;無尾箭頭示伸縮泡孔;B.扇形游仆蟲背面單扇形型銀線系;C.鐮游仆蟲腹面含有排列不規則的多邊形至六邊形銀線網;無尾箭頭示伸縮泡孔;D.鐮游仆蟲背面雙闊口型銀線系;E、G.包囊游仆蟲腹面含有大小不一且排列不規則的多邊形至六邊形銀線網;有尾箭頭示波動膜;無尾箭頭示伸縮泡孔;F、H.包囊游仆蟲背面復雜型銀線系。CC:尾棘毛;DK:背觸毛;FC:額腹棘毛;LMC:左緣棘毛;MC:口棘毛;TC:橫棘毛。A~F: Silver nitrate impregnation micrographs; G,H: Scanning electron micrographs; Scale bars: 20 μm. A. The ventral side ofE.vannuscontains neatly arranged quadrilateral silver-line system;arrowhead depicts the contractile vacuole pore; B. Dorsal view ofE.vannus, showing the single-vannussilver-line system; C. The ventral silver-line system ofE.harpacontains irregular polygons to hexagons;arrowhead refers to the contractile vacuole pore; D. Dorsal view ofE.harpa, showing the double-eurystomussilver-line system; E, G. The ventral silver-line system ofE.encysticuscontains irregular meshwork of different sized polygons to hexagons;arrow depicts the paroral membrane; arrowhead marks the contractile vacuole pore; F, H. Dorsal view ofE.encysticus, showing the complex silver-line system. CC: Caudal cirri; DK: Dorsal kinety; FC: Frontoventral cirri; LMC: Left marginal cirri; MC: Migratory cirrus; TC: Transverse cirri.)
圖1 3種游仆蟲細胞分裂間期皮層銀線系形態
Fig.1 The silver-line system of threeEuplotesspecies in non-dividing stage
在細胞背面,新背觸毛及其周圍銀線系的發生與腹皮層纖毛器和銀線系的分化相伴進行。首先在每個老背觸毛列的中部范圍各形成1列新背觸毛原基,于每列原基中基體間分開成一定距離時,在各個毛基體的周圍產生細小的銀線網,其中在基體左側的新銀線網先于右側開始發育,接著在每個背觸毛基體兩側各形成1列新銀線網(見圖2I)。細胞形成分裂溝時,每列新背觸毛在其中部分開成前、后兩部分,相鄰兩列新背觸毛間的老銀線網消失,背觸毛左、右兩側的新銀線網會合成1列。隨著細胞的演化,新形成的背觸毛和新銀線網與同一列中的其他老背觸毛及老銀線網,最終成為新仔蟲的背觸毛列及其背皮層銀線網(見圖2J)。
2.2.2 分裂期鐮游仆蟲銀線系的形態發生 鐮游仆蟲腹面纖毛器和銀線系的發生與扇形游仆蟲相似,但額腹橫棘毛原基及其周圍銀線系的發生卻略有不同。在新口原基形成時(見圖3A),于額腹橫棘毛皮層區形成2組各含5列原基的前、后額腹橫棘毛原基區,其每列原基不斷變粗,并按由后至前、由左至右順序分化成3、3、3、3、2段棘毛原基(見圖3B、C)。與此同時,在每段棘毛原基的左側和右側按自后至前的順序發生細小的新銀線網,其銀線網不斷發育伸展,于每列原基的周圍各形成許多排列不規則的新銀線網,最終在新的額腹橫棘毛原基區形成多邊形銀線網(見圖3D~F、H)。此外,伴同老口側膜前方唇和新口圍帶原基右側皮層區的前、后2個新棘毛原基的分化,其周圍也發生細小的銀線網并不斷發育伸展,形成多邊形銀線網(見圖3B、D)。隨著額腹橫棘毛原基進一步發育形成新棘毛、蟲體演化成前、后仔蟲時,腹面殘留的老棘毛及老銀線網消失,新棘毛及新銀線網相繼定位,成為前、后仔蟲的額腹橫棘毛和腹面銀線網(見圖3G)。
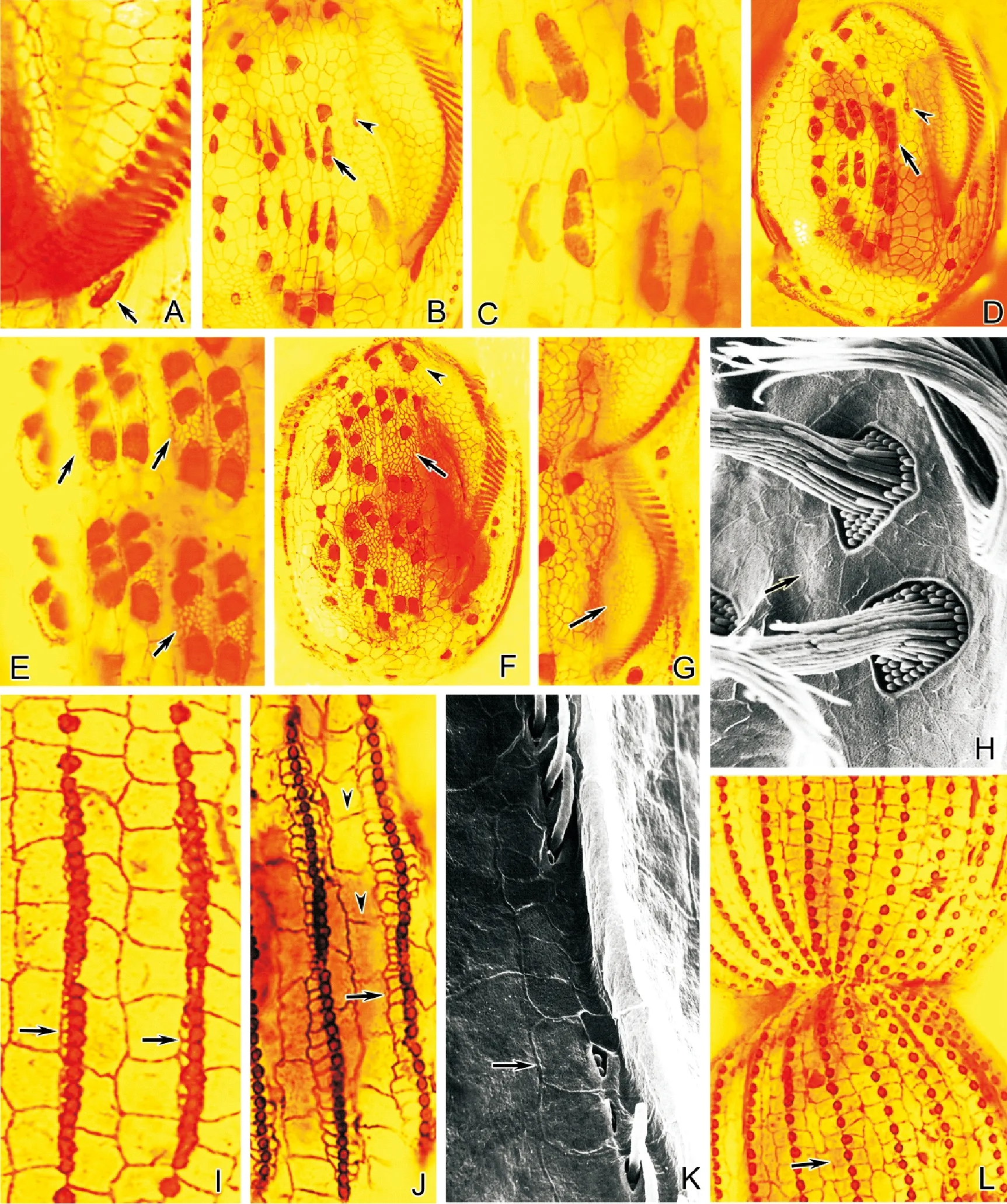
(A~F和H~J:銀浸染色照片;G:掃描電鏡照片。A.口原基分化出小群毛基體(箭頭所示);B.口原基周圍銀線網(箭頭所示);C.后仔蟲口腔前方銀線網(箭頭所示);D.額腹橫棘毛原基出現(箭頭所示);E.示分成段的額腹橫棘毛原基和其周圍銀線網(有尾箭頭),以及前、后仔蟲口棘毛原基(無尾箭頭);F、G.分成段的額腹橫原基及其周圍銀線網的細節圖;H.腹面新銀線網(有尾箭頭)和老銀線網(無尾箭頭);I、J.背觸毛原基發生及其周圍新銀線網。有尾箭頭示背面新銀線網;無尾箭頭示老銀線網。A~F&H~J: Silver nitrate impregnation micrographs; G:Scanning electron micrograph. A.Asmall number of basal bodies are differentiating from the oral primordium (arrow);B. Silver-line system around the oral primordium (arrow);C. Silver-line system in front of the buccal cavity of proter (arrows);D. The appearance of frontoventral-transverse anlagen (arrows);E.Fragmented frontoventral-transverse anlagen and the silver-line system around them (arrow); arrowheads mark themigratory cirrus anlagen of proter and opisthe;F, G. Detail of the fragmented frontoventral-transverse anlagen and the silver-line system around them; H.Ventral view of the new (arrows) and the old (arrowheads)silver-line system; I, J. Dorsal bristle basal bodies appear and the new silver-line system around them. Showing the dorsal view of the new (arrows) and the old (arrowheads) silver-line system.)
圖2 扇形游仆蟲皮層銀線系的形態發生
Fig.2 Morphogenesis ofE.vannus
在細胞背面,新背觸毛及其周圍銀線系的發生與扇形游仆蟲的基本相似,只是在細胞形成分裂溝時,每列新背觸毛在其中部分開成前、后兩部分,相鄰2列新背觸毛間的老銀線網消失,背觸毛左、右兩側的新銀線網匯合成2列(見圖3I~L)。隨著細胞的演化,新形成的背觸毛和新銀線網與同一列中的其他老背觸毛及老銀線網,最終成為新仔蟲的背觸毛列及其背皮層新銀線網。
(A~G、I、J、L:銀浸染色照片;H,K:掃描電鏡照片。A.口原基分化出小群毛基體(箭頭所示);B.示開始分段的額腹橫棘毛原基(有尾箭頭)和前仔蟲口棘毛原基(無尾箭頭);C.開始分段的額腹橫棘毛原基及其左側新銀線網的細節圖;D.示分成段的額腹橫棘毛原基和其周圍新銀線網(箭頭),以及前仔蟲口棘毛原基(無尾箭頭);E.示分成段的額腹橫棘毛原基及其周圍銀線網的細節圖(箭頭);F、H.示腹面新銀線網(有尾箭頭)和老銀線網(無尾箭頭);G.后仔蟲口腔前方銀線網(箭頭所示);I. 背觸毛原基發生及其周圍新銀線網;J、K. 示背觸毛原基周圍新銀線網發展擴大(有尾箭頭),及老銀線網(無尾箭頭);L.背面新銀線網(箭頭所示)。A~G, I, J, L:Silver nitrate impregnation micrographs; H, K: Scanning electron micrographs. A. A small number of basal bodies are differentiating from the oral primordium (arrow);B. The frontoventral-transverse anlagen (arrow)start to fragment;migratory cirrus anlagen of proter (arrowhead); C. Detail of the frontoventral-transverse anlagen start to fragment and the new silver-line system on the left; D. Fragmented frontoventral-transverse anlagen and the new silver-line system around them(arrow); migratory cirrus anlagen of proter (arrowhead); E. Detail of the fragmented frontoventral-transverse anlagen and the silver-line system around them (arrows); F, H.Ventral view of the new (arrows)and the old (arrowhead)silver-line system; G. Silver-line system in front of the buccal cavity of proter (arrow); I. Dorsal bristle basal bodies appear and the new silver-line system around them;J, K. New silver-line system around the dorsal bristle basal bodiesis extending further (arrows);arrowheads depict the old silver-line system;L.Dorsal view of the new (arrow) silver-line system.)
圖3 鐮游仆蟲皮層銀線系的形態發生
Fig.3 Morphogenesis ofE.harpa
2.2.3 分裂期包囊游仆蟲銀線系的形態發生 包囊游仆蟲腹面纖毛器和銀線系的發生與扇形游仆蟲相似, 但額腹橫棘毛原基及其周圍銀線系的發生卻略有不同。在新口原基形成時(見圖4A),于額腹橫棘毛皮層區形成2組各含5列原基的前、后額腹橫棘毛原基區,其每列原基不斷變粗,并按由后至前、由左至右順序分化成3、3、3、2、2段棘毛原基(見圖4B、C)。與此同時,在每段棘毛原基的左側和右側按自后至前的順序發生細小的新銀線網,其銀線網不斷發育伸展,于每列原基的周圍各形成許多排列不規則的新銀線網,最終在新的額腹橫棘毛原基區形成多邊形銀線網(見圖4E、F)。隨著額腹橫棘毛原基進一步發育形成新棘毛、蟲體演化成前、后仔蟲時,腹面殘留的老棘毛及老銀線網消失,新棘毛及新銀線網相繼定位,成為前、后仔蟲的額腹橫棘毛和腹面銀線網。

(A、B、D~J:銀浸染色照片;C:掃描電鏡照片。A.口原基分化出小群毛基體(箭頭所示);B.額腹橫棘毛原基(箭頭所示);C、E.分段的額腹橫棘毛原基及其周圍銀線網(箭頭所示);D.后仔蟲口圍帶原基小膜(箭頭所示);F.腹面新銀線網(箭頭所示);G.新背觸毛基體發生(箭頭所示);H. 背觸毛原基及其周圍新銀線網(有尾箭頭);老銀線網(無尾箭頭);I.背觸毛原基周圍新銀線網(有尾箭頭)發展擴大;老銀線網(無尾箭頭);J.背面新銀線網(有尾箭頭)和老銀線網(無尾箭頭)。A, B, D~J: Silver nitrate impregnation micrographs; C:Scanning electron micrographs.A. A small number of basal bodies are differentiating from the oral primordium;B. Frontoventral-transverse anlagen (arrow);C, E.Fragmented frontoventral-transverse anlagen and the silver-line system around them (arrows); D. The new formed membranelles of opisthe (arrow);F. Ventral view of the new silver-line system (arrow); G.New dorsal bristle basal bodies appear (arrow); H.Dorsal bristle basal bodies and the new silver-line system around them (arrow); the old silver-line system (arrowhead);I.New silver-line system around the dorsal bristle basal bodies is extending further (arrow);the old silver-line system (arrowhead); J. Dorsal view of the new (arrow)and the old (arrowhead) silver-line system.)
圖4 包囊游仆蟲皮層銀線系的形態發生
Fig.4 Morphogenesis ofE.encysticus
在細胞背面,新背觸毛及其周圍銀線系的發生與腹皮層纖毛器和銀線系的分化相伴進行。首先在每個老背觸毛列的中部范圍各形成1列新背觸毛原基(見圖4G),于每列原基中基體間分開成一定距離時,在各個毛基體的周圍產生細小的銀線網,其中在基體左側的新銀線網先于右側開始發育,接著在每個背觸毛基體兩側各形成多列新銀線網。細胞形成分裂溝時,每列新背觸毛在其中部分開成前、后兩部分,相鄰2列新背觸毛間的老銀線網消失,背觸毛左、右兩側的新銀線網會合成多列。隨著細胞的演化,新形成的背觸毛和新銀線網與同一列中的其他老背觸毛及老銀線網,最終成為新仔蟲的背觸毛列及其背皮層銀線網(見圖4H~J)。
3 討論
3.1 游仆蟲腹面銀線系形態
游仆蟲背面銀線系因具有種類的特異性,一直以來被應用于游仆蟲類的物種鑒定與分類,而關于腹面銀線系的特點未見進一步探討[2,9]。本實驗結果顯示扇形游仆蟲腹面銀線網排列整齊且稀疏,而鐮游仆蟲和包囊游仆蟲腹面銀線網形狀不規則且排列緊密。這種不同的排列特征分別與其背面銀線網的排列規律相類似,尤其體現在銀線網的緊密程度上;而背腹面銀線網的緊密程度又與背觸毛列中的基體數目密切相關。這種對應關系亦出現在其他已有銀線系報道的游仆蟲種類中[16-23],即在所有銀線系類型中,背觸毛基體數目越多,背面銀線網越密集,腹面網格也相應密集。據此推測,游仆蟲腹面銀線系雖不能如背面銀線系以具體類型劃分,但其形態特征與背面銀線系具有一定程度的一致性,這顯示細胞在背腹面銀線系結構形成過程中的整體性調控。
3.2 游仆蟲銀線系的形態發生
游仆蟲皮層纖毛器、纖毛器附屬微管及皮層銀線網等是一類重要的的皮層細胞骨架[12],目前對該類纖毛蟲皮層纖毛器及其纖毛器附屬微管的形態發生及其細胞調控的研究已經取得較豐富的資料[13,24-35],但對其銀線系的形態發生尚未見報道。本文應用銀浸染色法及掃描電鏡術顯示,扇形游仆蟲、鐮游仆蟲和包囊游仆蟲3種纖毛蟲皮層銀線網的形態發生與細胞皮層纖毛器的發生是相伴進行的,其腹皮層銀線網在口圍帶原基區分化形成的小膜原基和額腹橫棘毛原基區原基列分段形成的棘毛原基周圍發生,背皮層銀線網在每列背觸毛原基的各個背觸毛基體周圍發生,由于此時的口圍帶小膜原基、棘毛原基和背觸毛基體已經形成纖毛基體復合單元和基體單元[24,36-40],也即其銀線網的形態發生是在纖毛器原基形成口圍帶小膜原基、棘毛原基基體復合單元和背纖毛基體單元后啟動的。根據Shao et al.劃分的游仆蟲屬額腹橫棘毛原基的分段模式,扇形游仆蟲和鐮游仆蟲屬于charon-type,II至V額腹橫棘毛原基分段為3∶3∶3∶3∶2,包囊游仆蟲則屬于affinis-type,分段為3∶3∶3∶2∶2[41],即前2種游仆蟲與包囊游仆蟲相比,其第5列額腹橫棘毛原基多斷裂生成一個棘毛3/V的基體。本文觀察到在扇形游仆蟲和鐮游仆蟲中,新銀線網于棘毛3/V基體周圍產生并隨該基體的遷移而持續分化;而對于該部位無此棘毛基體的包囊游仆蟲,新銀線網產生僅限于2/V和1/V 2個棘毛原基附近。這進一步驗證了在非纖毛區無新銀線網獨立發生,新銀線網的發生與分化與纖毛基體密切相關。據以上資料推測,所述的口圍帶小膜、棘毛基體復合單元和背觸毛基體單元不僅對相應纖毛器結構行使微管組織中心作用,也可能對相應皮層區銀線網的發生具有定位作用和組織中心的作用。結果還顯示,3種游仆蟲圍繞在基體左側的銀線網先于右側開始發育,與游仆蟲新棘毛原基發生和分化過程相一致[25],據此推測銀線系的分化可能也具有一致的方向性。因此,以上所得結果可為揭示纖毛蟲生命活動中細胞皮層結構的分化及其調控機理提供新的資料。
3.3 老結構在游仆蟲新銀線系形成過程中的作用
關于游仆蟲皮層纖毛器形態發生過程中新、老結構的演化及其作用關系已有較多研究[25,42],但對銀線系發生中新、老結構的更替及其作用機理等方面尚未取得相關資料。本文觀察到,3種游仆蟲腹面銀線系發生中老纖毛結構及其周圍銀線網按序先后瓦解,其中離新結構發生區近的老結構先瓦解,由此使新結構不斷延伸發展,據此推測老銀線網對新結構的形成可能起到了誘導定向作用;細胞出現分裂溝時,在每一背觸毛列的中部范圍相鄰2列新背觸毛間的老銀線網消失,背觸毛左、右兩側的新銀線網會合形成新銀線網,并且前仔蟲前半部分和后仔蟲后半部分的老銀線網保留下來共同組成新細胞的背銀線網,結果表明,游仆蟲的老銀線網不僅參與了新結構的組成,并對新結構的形成具有物質貢獻。
4 結語
本文揭示了游仆蟲銀線系形態發生過程中的結構特征,為研究這類微管類細胞骨架提供了新的資料。但本研究并未揭示新銀線網在發生時,皮層纖毛器及其附屬微管和銀線系在皮層下的變化情況,因此,在本研究的基礎上,可以通過透射電鏡等方法進一步研究探討銀線系、皮層纖毛器及其附屬微管之間的關系,構建其在形態發生時的三維模型,為游仆蟲皮層模式形成提供更多資料。
[1] Tuffrau M. Les caractères specifiques dans le genreEuplotes(Note Préliminaire) [J]. Bulletin de la Societe Zoologique de France, 1954, 79: 463-465.
[2] Tuffrau M. Revision of genusEuploteson the bases of structural comparison [J]. Hydrobiologia, 1960, 15(1): 1-77.
[3] Tuffrau M, Fryd-Versavel G, Tuffrau H, et al. Description ofEuplotesversatilisn. sp., a marine tropical ciliate exhibiting an unusually extensive phenotypic plasticity [J]. European Journal of Protistology, 2000, 36(4): 355-366.
[4] Chen X, Zhao Y, Al-Farraj S A,et al. Taxonomic descriptions of two marine ciliates,Euplotesdammamensisn. sp. andEuplotesbalteatus(Dujardin, 1841) Kahl, 1932 (Ciliophora, Spirotrichea, Euplotida), collected from the Arabian Gulf, Saudi Arabia [J]. Acta Protozoologica, 2013, 52(2): 73-89.
[5] Dai R, Xu K, He Y. Morphological, physiological, and molecular evidences suggest thatEuplotesparawoodruffiis a junior synonym ofEuploteswoodruffi(Ciliophora, Euplotida) [J]. The Journal of Eukaryotic Microbiology, 2013, 60(1): 70-78.
[6] Chen X, Ma H, Al-Rasheid K A S. Taxonomic description of a new marine ciliate,Euplotesqingdaoensisn. sp. (Ciliophora: Euplotida) [J]. Chinese Journal of Oceanology and Limnology, 2014, 32(2): 426-432.
[7] Giuseppe G D, Erra F, Frontini F P, et al. Improved description of the bipolar ciliate,Euplotespetzi, and definition of its basal position in theEuplotesphylogenetic tree [J]. European Journal of Protistology, 2014, 50(4): 402-411.
[8] Giuseppe G D, Dini F, Vallesi A, et al. Genetic relationships in bipolar species of the protist ciliate,Euplotes[J]. Hydrobiologia, 2015, 761(1): 71-83.
[9] Curds C R. A guide to the species of the genusEuplotes(Hypotrichida, Ciliatea) [J]. Bulletin of the British Museum (Natural History) Zoology, 1975, 28: 5-9.
[10] Gates M A, Curds C R. The argyrome of the genusEuplotes[J]. Bulletin of the British Museum (Natural History) Zoology, 1979, 35: 127-200.
[11] 朱慧, 鄒士法, 李藝松, 等. 用非離子去垢劑抽提獲得的小游仆蟲皮層細胞骨架的構形[J]. 動物學研究, 2004, 25(5): 422-428. Zhu H, Zou S, Li Y, et al. Cortical cytoskeleton ofEuplotesgracilis(Protozoa, Ciliophora) by non-ionic detergent extraction [J]. Zoological Research, 2004, 25(5): 422-428.
[12] 李藝松, 柳偉君, 顧福康. 伍氏游仆蟲皮層微管類細胞骨架的熒光標記[J]. 復旦學報 (自然科學版), 2008, 47(3): 364-369. Li Y, Liu W, Gu F. Cortical microtubular cytoskeleton inEuploteswoodruffirevealed by fluorescent labeling [J]. Journal of Fudan University (Natural Science), 2008, 47(3): 364-369.
[13] 林欽, 范鑫鵬, 孫磊, 等. 小腔游仆蟲 (Euplotesaediculatus) 皮層微管胞器的熒光標記[J]. 華東師范大學學報 (自然科學版), 2013(5): 53-60. Lin Q, Fan X, Sun L, et al. Microtubular organelles inEuplotesaediculatus (Ciliophora: Hypotrichida) revealed by fluorescent labeling [J]. Journal of East China Normal University (Natural Science), 2013(5): 53-60.
[14] 史新柏. 銀浸法在纖毛蟲研究中的應用[J]. 哈爾濱師范學院學報 (自然科學版), 1963, 1: 79-83. Shi X. Application of silver-impregnation methods in the study of ciliates [J]. Journal of Harbin Normal University (Natural Science), 1963, 1: 79-83.
[15] 顧福康, 倪兵. 原生動物掃描電鏡樣品制備方法的探討[J]. 電子顯微學報, 1993(6): 525-529. Gu F, Ni B. The exploration of preparing protozoan specimens for scanning electron microscopy [J]. Journal of Chinese Electron Microscopy Society, 1993(6): 525-529.
[16] Borror A C.EuplotesminutaYocom (Ciliophora, Hypotrichida) [J]. Journal of Protozoology, 1962, 9(3): 271-273.
[17] Pan Y, Li L, Shao C, et al. Morphology and ontogenesis of a marine ciliate,Euplotesbalteatus(Dujardin, 1841) Kahl, 1932 (Ciliophora, Euplotida) and definition ofEuploteswilbertinov. spec [J].Acta Protozoologica, 2012, 51(1): 29-38.
[18] Jong O, Mann K. Redescription of newly recorded ciliate,Euplotesmuscorum(Ciliophora: Polyhymenophora: Hypotrichida) and comparison with related species from Korea [J]. Korean Journal of Systematic Zoology, 2003, 19(2): 227-235.
[19] Song W, Warren A, Hill B F. Description of a new freshwater ciliate,Euplotesshanghaiensisnov. spec. from China (Ciliophora, Euplotidae) [J]. European Journal of Protistology, 1998, 34 (2): 104-110.
[20] Liu M, Fan Y, Miao M, et al. Morphological and morphogenetic redescriptions and SSU rRNA gene-based phylogeny of the poorly-known speciesEuplotesamietiDragesco, 1970 (Ciliophora, Euplotida) [J]. Acta Protozoologica, 2015, 54: 171-182.
[21] Fan X, Huang J, Lin X, et al. Morphological and molecular characterization ofEuplotesencysticus(Protozoa: Ciliphora: Euplotida) [J]. Journal of the Marine Biological Association of the United Kingdom, 2010, 90(7): 1411-1416.
[22] Jiang J, Zhang Q, Warren A, et al. Morphology and SSU rRNA gene-based phylogeny of two marineEuplotesspecies,E.orientalisspec. nov. andE.raikoviAgamaliev, 1966 (Ciliophora, Euplotida) [J]. European Journal of Protistology, 2010, 46(2): 121-132.
[23] Jiang J, Zhang Q, Hu X, et al. Two new marine ciliates,Euplotessinicussp. Nov. andEuplotesparabalteatussp. nov., and a new small subunit rRNA gene sequence ofEuplotesrariseta(Ciliophora, Spirotrichea, Euplotida) [J]. International Journal of Systematic and Evolutionary Microbiology, 2010, 60: 1241-1251.
[24] 顧福康, 龐延斌, 張作人. 一種游仆蟲無性生殖的研究II.無性分裂過程中皮層結構的形態發生[J]. 動物學報, 1987, 33(4): 362-369. Gu F, Pang Y, Zhang Z. On the asexual reproduction of the genusEuplotesII. Morphogenesis of cortical structures during the asexual division [J]. Acta Zoologica Sinica, 1987, 33(4): 362-369.
[25] 顧福康, 張作人. 一種游仆蟲棘毛基部纖維的形態及其在形態發生過程中的演化[J]. 動物學研究, 1989, 10(2): 89-96. Gu F, Zhang Z. The morphology of cirral-base-associated fibers inEuplotessp. and their morphogenesis [J]. Zoological Research, 1989, 10(2): 89-96.
[26] 牛延寧, 吳月華, 倪兵, 等. 包囊游仆蟲纖毛器微管在不同生理狀態下的分化[J]. 華東師范大學學報 (自然科學版), 2007(6): 106-111. Niu Y, Wu Y, Ni B, et al. Ciliature microtubule differentiation ofEuplotesencysticusin different physiological conditions [J]. Journal of East China Normal University (Natural Science), 2007(6): 106-111.
[27] 余齊耀, 張萌, 陳季武, 等. 顯示纖毛蟲細胞微管骨架的兩種熒光標記方法[J]. 生物學雜志, 2012, 29(1): 92-94. Yu Q, Zhang M, Chen J, et al. Two fluorescent labeling methods for revealing microtubular cytoskeleton in ciliates [J]. Journal of Biology, 2012, 29(1): 92-94.
[28] 吳娜, 周慧琳, 范鑫鵬, 等. 澳洲管膜蟲纖毛器微管的熒光標記及其激光掃描共聚焦顯微觀察[J]. 生物學雜志, 2015, 32(1): 30-33. Wu N, Zhou H, Fan X, et al. Observation on ciliature microtubules ofCyrtohymenaaustralisusing fluorescent labeling and laser scanning confocal microscopy [J]. Journal of Biology, 2015, 32(1): 30-33.
[29] Jiang J, Shao C, Xu H, et al. Morphogenetic observations on the marine ciliateEuplotesvannusduring cell division (Protozoa: Ciliophora) [J]. Journal of the Marine Biological Association of the United Kingdom, 2010, 90(4): 683-689.
[30] Rashmi F, Thorsten S, Sabine F, et al. Description of the HalophileEuplotesqatarensisnov. spec. (Ciliophora, Spirotrichea, Euplotida) isolated from the Hypersaline Khor Al-Adaid Lagoon in Qatar [J]. Journal of Eukaryotic Microbiology, 2016. DOI:10.111/jeu.12305.
[31] Fleury A. Dynamics of the cytoskeleton during morphogenesis in the ciliateEuplotesI. Basal bodies related microtubular system [J]. European Journal of Protistology, 1991, 27: 99-114.
[32] Fleury A. Dynamics of the cytoskeleton during morphogenesis in the ciliateEuplotesII. Cortex and continuous microtubular systems [J]. European Journal of Protistology, 1991, 27(3): 220-237.
[33] 畢紅衛, 龐延斌. 卡龍游仆蟲Euplotescharon的形態和形態發生的研究[J]. 華東師范大學學報 (自然科學版), 2001(2): 71-77. Bi H, Pang Y. The studies on the morphology and morphogenesis ofEuplotescharon[J]. Journal of East China Normal University (Natural Science), 2001(2): 71-77.
[34] 王梅, 宋微波. 卡龍游仆蟲無性生殖期間的形態發生學研究[J]. 動物學研究, 1995, 16(3): 233-238. Wang M, Song W. Morphogenetical studies on the marine ciliateEuplotescharon[J]. Zoological Research, 1995, 16(3): 233-238.
[35] 龐延斌, 魏紅兵. 小腔游仆蟲Euplotesaediculatus形態和形態發生的研究[J]. 華東師范大學學報 (自然科學版), 1999(1): 103-109. Pang Y, Wei H. Studies on the morphology and morphogenesis inEuplotesaediculatus[J]. Journal of East China Normal University (Natural Science), 1999(1): 103-109.
[36] Jerka-Dziadosz M, Frankel J. An analysis of the formation of ciliary primordia in the hypotrich ciliateUrostylaweissei[J].Journal of Protozoology, 1969, 16(4): 512-537.
[37] Jerka-Dziadosz M. Ultrastructural study on development of the hypotrich ciliateParaurostylaweisseiI. Formation and morphogenetic movements of ventral ciliary primordial [J]. Protistologica, 1980, 16: 571-589.
[38] Jerka-Dziadosz M. Ultrastructural study on development of hypotrich ciliateParaurostylaweisseiII. Formation of the adoral zone of membranelles and its bearing of problems of ciliate morphogenesis [J]. Protistologica, 1981, 17: 67-81.
[39] Jerka-Dziadosz M. Ultrastructural study on development of hypotrich ciliateParaurostylaweisseiIII. Formation of paroral membranelles and essary on comparative morphogenesis [J]. Protistologica, 1981, 17: 83-97.
[40] Jerka-Dziadosz M. Ultrastructural study on development of the hypotrich ciliateParaurostylaweisseiIV. Morphogenesis of dorsal bristles and caudal cirri [J]. Protistologica, 1982, 18: 237-251.
[41] Shao C, Ma H, Gao S, et al. Reevaluation of cortical developmental patterns inEuplotes(s.1.), including a morphogenetic redescription ofE.charon(Protozoa, Ciliophora, Euplotida) [J]. Chinese Journal of Oceanology and Limnology, 2010, 28(3): 593-602.
[42] 顧福康, 張作人. 腹毛目纖毛蟲無性生殖周期中形態發生的研究[J]. 華東師范大學學報 (自然科學版), 1990, 2: 85-93. Gu F, Zhang Z. Study on morphogenesis of hypotrich ciliates during asexual reproduction [J]. Journal of East China Normal University (Natural Science), 1990, 2: 85-93.
責任編輯 朱寶象
更正聲明
《中國海洋大學學報(自然科學版)》第11期中“DNA條形碼技術在鯔科魚類鑒定中的應用”論文刊登的作者信息、單位信息的中英文有誤,在此進行更正。正確的作者信息、單位信息的中英文如下:
劉 璐1, 孫典榮2, 李純厚2, 韓志強3, 高天翔3, 宋 娜1??
(1.中國海洋大學水產學院,山東 青島 266003;2.中國水產科學研究院南海水產研究所,廣東 廣州 510300;3.浙江海洋大學水產學院,浙江 舟山 316022)
LIU Lu1, SUN Dian-Rong2, LI Chun-Hou2, HAN Zhi-Qiang3, GAO Tian-Xiang3, SONG Na1
(1.College of Fisheries, Ocean University of China, Qingdao 266003, China;2.South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Guangzhou 510300, China;3.Fishery College, Zhejiang Ocean University, Zhoushan 316022, China)
特此聲明!
中國海洋大學學報(自然科學版)編輯部
Morphogenesis of the Cortical Silver-Line Systems in the Ciliate Genus Euplotes (Protozoa, Ciliophora)
XIE Dong-Mei, FAN Xin-Peng, NI Bing, GU Fu-Kang
(School of Life Sciences, East China Normal University, Shanghai 200241, China)
The silver-line system ofEuplotescorresponds to the cytoskeleton network, which is composed of microtubules at the ultrastructure level, and its morphogenesis pattern during cell division is rarely reported. Based on the application of silver nitrate impregnation and scanning electron microscopy, and usingE.vannus,E.harpaandE.encysticusas the representatives, the morphogenesis of three types of cortical silver-line systems in genusEuploteswas examined in this study.The result showed that the morphogenesis of three types of cortical silver-line systems in the genusEuplotesshared the following features: the ventral silver-line system occurred around the new membranelles and the fragmented frontoventral-transverse anlagen; and the dorsal silver-line system occurred around the basal bodies of the dorsal bristle primordia. Thus, it was speculated that the composite element of basal bodies was not only microtubule organizing center, but also the center of the silver-line system. The new meshwork on the left of the basal bodies developed ahead of the meshwork on the right, thus, it was speculated that the silver-line system oriented similarly with ciliary organelles during their morphogenesis. In addition, the old meshwork near new silver-line first disintegrated, which led to the new silver-line system to develop constantly. It was assumed that the old silver-line system played a role of induction and orientation in the formation of new structure. The old silver-line system locating in the first half of the proter and in the latter part of the opisthe was reserved to form the dorsal silver-line system in new cells. It was supposed that the old silver-line system participated in the formation of new structure and also had a material contribution to it.
Euplotes; silver-line system; morphogenesis; silver nitrate impregnation; scanning electron microscopy
國家自然科學基金項目(31172042;31572223)資助 Supported by National Natural Science Foundation of China (31172042;31572223)
2015-11-06;
2016-03-16
謝冬梅(1989-),女,碩士生。E-mail: zynx9194@sina.com
?? 通訊作者:E-mail: xpfan@bio.ecnu.edu.cn
Q952;Q954
A
1672-5174(2016)12-041-10
10.16441/j.cnki.hdxb.20150385
謝冬梅, 范鑫鵬, 倪兵, 等. 游仆蟲(原生動物,纖毛門)皮層銀線系的形態發生模式[J]. 中國海洋大學學報(自然科學版), 2016, 46(12): 41-50.
XIE Dong-Mei, FAN Xin-Peng, NI Bing, et al. Morphogenesis of the cortical silver-line systems in the ciliate genusEuplotes(Protozoa, Ciliophora)[J]. Periodical of Ocean University of China, 2016, 46(12): 41-50.
