紫蘇種子脂肪酸代謝及關鍵酶基因調控油脂合成規律的研究
2016-12-27 06:35:46王計平張玲慧王彥尊李潤植
中國糧油學報 2016年3期
關鍵詞:油脂
王計平 張玲慧 趙 靜 王彥尊 李潤植
(山西農業大學農學院,太谷 030801)
紫蘇種子脂肪酸代謝及關鍵酶基因調控油脂合成規律的研究
王計平 張玲慧 趙 靜 王彥尊 李潤植
(山西農業大學農學院,太谷 030801)
為了研究種子油脂及脂肪酸生物合成機制,特別是α-亞麻酸的高水平積累,采用氣相色譜法分析7個紫蘇品種的成熟種子及晉蘇1號種子不同發育時期脂肪酸組成、含量及其動態變化;實時熒光定量PCR檢測TAG生物合成關鍵酶基因在紫蘇不同組織及種子發育不同時期的表達特性,分析基因表達與脂肪酸及α-亞麻酸合成積累之間的關系。結果表明,紫蘇種子中不飽和脂肪酸(18∶1,18∶2,18∶3)含量占總脂肪酸90%以上,種子發育過程中,總脂肪酸含量隨著鮮重增加而不斷增加,種子形態也經歷由多水透明狀轉變為乳白色或深紫色最后成為褐色;隨著種子不斷發育,亞油酸含量逐漸降低,而α-亞麻酸含量不斷增加,到種子成熟時達總脂肪酸質量的60%以上。晉蘇1號作為高含α-亞麻酸的優質品種,α-亞麻酸含量達456.6 mg/g,亞麻酸與亞油酸之比為9∶1。Real-time PCR 分析結果顯示PfDGAT1和PfPDAT在紫蘇莖和根系中的表達量均很低,而在葉片和種子中高量表達,在種子發育不同時期,PfDGAT1表達量呈先升高后降低的趨勢,開花后30 d,表達量最高。PfDGAT1基因超量表達之后緊接著α-亞麻酸和總油脂快速積累,表明PfDGAT1在紫蘇α-亞麻酸和總油脂積累過程中起關鍵作用。
紫蘇 種子油脂 脂肪酸 二酰甘油酰基轉移酶基因(DGAT1) 磷脂二酰甘油酰基轉移酶(PDAT)
紫蘇(PerillafrutescensL.)是唇形科紫蘇屬一年生草本植物,有較強的適應性,可藥食兩用,廣泛分布于東亞各國,在我國已有2 000多年的栽培歷史。紫蘇籽出油率高達45%~55%,含有豐富的不飽和脂肪酸,占總脂肪酸含量的90%以上,其中α-亞麻酸質量分數達55%~65%,α-亞麻酸是人體組織細胞必需的脂肪酸,在體內參與磷脂的合成和代謝。更為重要的是,α-亞麻酸在人體細胞組織內可進一步合成人體必需的2種生命活性因子,即二十二碳六烯酸(22∶6 n-3,DHA)和二十碳五烯酸(20∶5 n-3, EPA)。因此,開發基于植物源的α-亞麻酸營養保健醫療產品,越來越引起國內外學術界和產業界的高度關注。作為一種營養保健食用油,紫蘇籽油具有降血壓、降低膽固醇、促進視網膜發育、提高記憶力、增強免疫力、延緩衰老、預防老年癡呆等功效[1-2]。目前,關于植物種子脂肪酸合成積累機制及代謝工程的研究已成為當今植物油脂化學與分子生物學,生物技術和油料作物遺傳育種研究的重點領域[3-7],但有關紫蘇α-亞麻酸生物合成積累及調控機制的研究相對較少。本試驗通過研究控制紫蘇種子α-亞麻酸合成及積累的關鍵酶基因,即選擇性轉移α-亞麻酸于三酰基甘油脂(TAG)分子中的二酰甘油酰基轉移酶(DGAT1)和磷脂二酰甘油酰基轉移酶(PDAT)的表達特性,結合紫蘇種子含油率及脂肪酸成分變化,揭示油脂合成關鍵酶基因表達對油脂合成的影響。
1 材料與方法
1.1 主要儀器設備
Agilent 7890A氣相色譜儀:美國安捷倫科技公司;Applied Biosystems 7500 Fast Real-Time PCR System(實時熒光定量PCR儀):美國應用生物系統公司。
1.2 供試材料
試驗所用材料由山西省農業科學院棉花研究所提供,紫蘇品種為中北2號、晉蘇1號、Y-早、Y-7、Y-4、Y-2、白蘇。
1.3 試驗方法
1.3.1 脂肪酸提取
選取紫蘇7個品種的成熟種子及于盛花期標記始開花朵,開花后10、20、30、40 d,分別取發育中的蒴果(四棱種)15粒于EP管中,立即放入液氮中速凍(-80 ℃保存),每次取材3次重復。樣品經真空冷凍干燥過夜,稱取凍干種子樣品(8~12 mg)置于干燥玻璃試管中。加入Tri17∶0(用于內對照),每mg干樣品加10 μg Tri17∶0(即每mg樣品加1 μL 10 mg/mL Tri17∶0溶液),在玻璃試管內小心研磨。加入1 mL氯仿∶甲醇(2∶1)溶液,用玻璃棒攪拌幾分鐘后靜置,加0.9% KCl 0.5 mL攪拌后再加入1 mL氯仿,混勻,離心,使其分層。用玻璃吸管將底部氯仿相吸出,并移至另一個新的玻璃管。將含有氯仿相的試管置于室溫下自然蒸發(留下10~20 μL即可)。向上述試管中加入0.5 mL甲醇鈉(含BHT),混勻后置于搖床上震蕩20~45 min。之后加入0.5 mL異辛烷,混勻后,離心,使溶液分層,再加入0.9% KCl 1 mL,混勻,再離心至分層。吸取上層(含有脂肪酸甲酯),放入GC小瓶保存待測。
1.3.2 脂肪酸成分測定
通過安捷倫7890A氣相色譜儀進行分析,條件為VF-23MS毛細管柱30 m×0.25 mm×0.25 μm。進樣口溫度為260 ℃,載氣為高純氮氣,分流比30∶1,進樣體積1 μL,恒壓22 psi。程序升溫:初溫為110 ℃,保持3 min,以4 ℃/min升至220 ℃,保持15 min。FID檢測器溫度為260 ℃,尾吹氣N245 mL/min,空氣450 mL/min,氫氣40 mL/min。用安捷倫軟件進行分析,各種脂肪酸的鑒定通過標樣保留時間確認,同樣條件下平行測定3次。
1.3.3 實時熒光定量PCR檢測
通過脂肪酸組分及含量測定優選品種晉蘇1號,取其根、莖、葉、花,并于盛花期標記始開花朵,10、20、30、40 d后取發育中的蒴果(四棱種),每次取材分3次重復。RNA提取及第一鏈cDNA的合成分別按照TIAN Gen Biotech公司TRIZOL及FastQuant cDNA第一鏈合成試劑盒說明書進行。
實時熒光定量PCR引物分別為:PfDGAT1F:5’-TGGTGGAATGCGAGGACA-3’/PfDGAT1R:5’-GACACAAGAAACGCGATCAA-3’,PfPDATF:5’-GGCATTCCAACTGAAAGAGC-3’/PfPDATR:5’-GTTTTTCCACGCCACCCT-3’,18S rRNAF:5’-CGGCTACCACATCCAAGGAA-3’/18S rRNAF:5’-GCTGGAATTACCGCGGCT-3’。反應體系為:SYBR Premix EX TaqⅡ(Tli RnaseH Plus)(2×) 5 μL,10 μmol/L引物對各0.4 μL,模板cDNA 1 μL,ROX Reference Dye(50×) 0.2 μL,無菌純水補足10 μL;反應條件為:95 ℃ 3 min,30個循環(95 ℃ 30 s;60 ℃ 30 s;72 ℃ 20 s),10 ℃保溫。所用引物均由生工生物工程(上海)有限公司合成。
2 結果與分析
2.1 不同品種紫蘇成熟種子中脂肪酸組分分析
通過氣相色譜儀對7個不同品種紫蘇種子脂肪酸組成進行分析,結果表明:紫蘇籽油中含有棕櫚酸(C16∶0)、棕櫚一烯酸(C16∶1)、硬脂酸(C18∶0)、油酸(C18∶1)、亞油酸(C18∶2)、亞麻酸(C18∶3)、花生酸(C20∶0)、花生一烯酸(C20∶1)(表1)。其中,棕櫚酸、硬脂酸、油酸、亞油酸、亞麻酸含量較高,而棕櫚一烯酸、花生酸、花生一烯酸為微量脂肪酸(質量分數,<1%)。飽和脂肪酸棕櫚酸和硬脂酸的含量明顯較低,在總脂肪酸中分別占6%~7%和2%~3%。7個品種中晉蘇1號亞麻酸含量最高,占總脂肪酸質量的60.7%,達到456.6 mg/g,亞麻酸與亞油酸的比值為9∶1,其中不飽和脂肪酸占總脂肪酸的91%,7個品種總不飽和脂肪酸均達到90%以上(圖1),上述結果與已有研究結果一致[8-10]。
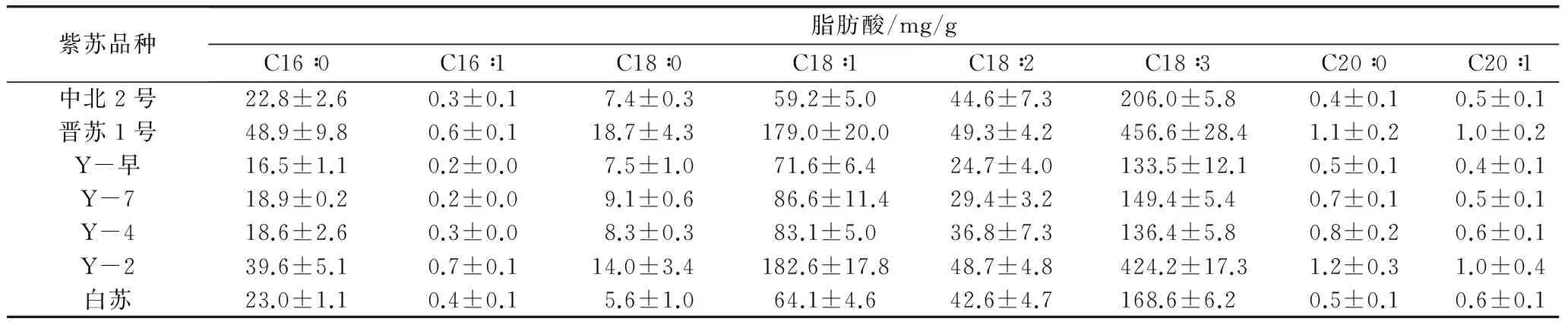
表1 不同品種紫蘇成熟種子脂肪酸組成及含量測定結果
2.2 紫蘇種子發育過程中脂肪酸組分分析
晉蘇1號果實發育初期4個果實晶瑩剔透,小如針尖;中期果實膨大,由透明狀轉向半透明或乳白色,通常同一基座上的4個果實發育大小不一;后期果實達到最大,為褐色,表皮有褐色網紋突出;末期果實不再增大,表皮的網紋變淡,果實全褐。在種子發育過程中定期取樣,通過氣相色譜進行脂肪酸組分分析(圖2)。結果表明,種子發育前期,隨著時間的推移,種子鮮重逐漸增大,到開花后30 d達到最大,然后呈現下降趨勢。而脂肪酸含量從開花后10 d開始緩慢增加,開花后30 d以后,種子鮮重降低的同時脂肪酸總量的積累迅速增大。
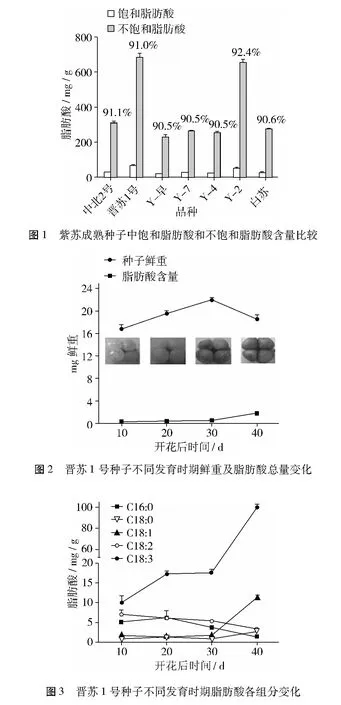
在種子發育過程中,脂肪酸積累量不斷增加,5種主要脂肪酸含量也在發生變化(圖3),每克種子中棕櫚酸(C16∶0)和亞油酸(C18∶2)的含量逐漸降低,硬脂酸(C18∶0)呈緩慢上升趨勢,油酸(C18∶1)開始增加緩慢,到了后期增加較快,變化趨勢與α-亞麻酸(C18∶3)類似。α-亞麻酸含量整體呈上升趨勢,開花后10 d種子中α-亞麻酸含量占總脂肪酸質量的40%,20 d的種子中 α-亞麻酸質量占53%,到開花后40 d時,平均每克鮮種子中α-亞麻酸質量達100.1 mg,占總脂肪酸質量的84%。
2.3 調控脂肪酸代謝關鍵酶時空表達分析
PfDGAT1在紫蘇不同組織的表達情況如圖4顯示,PfDGAT1在種子形成后期(開花后30 d)表達量最高,是葉片表達量的4.3倍,在花中表達量相對下調了1.1倍,該基因在紫蘇莖和根系中的表達量很低,分別是葉片表達量的0.32倍和0.53倍,說明種子是TAG合成的主要場所,PfDGAT1在紫蘇種子總油脂積累過程中行使重要功能。在種子發育過程中,PfDGAT1基因表達量呈現先升高后降低的趨勢。
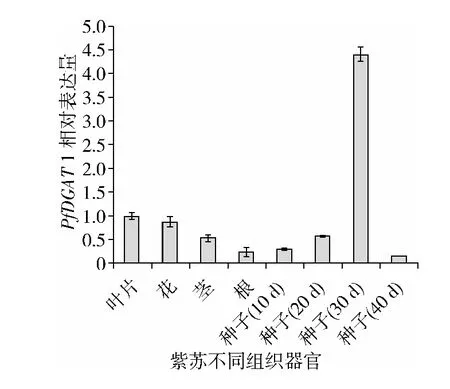
圖4 晉蘇1號不同組織器官PfDGAT1相對表達量的變化
根據圖5結果分析,紫蘇不同組織器官中PfPDAT基因表達量存在差異。在不同組織中表達量
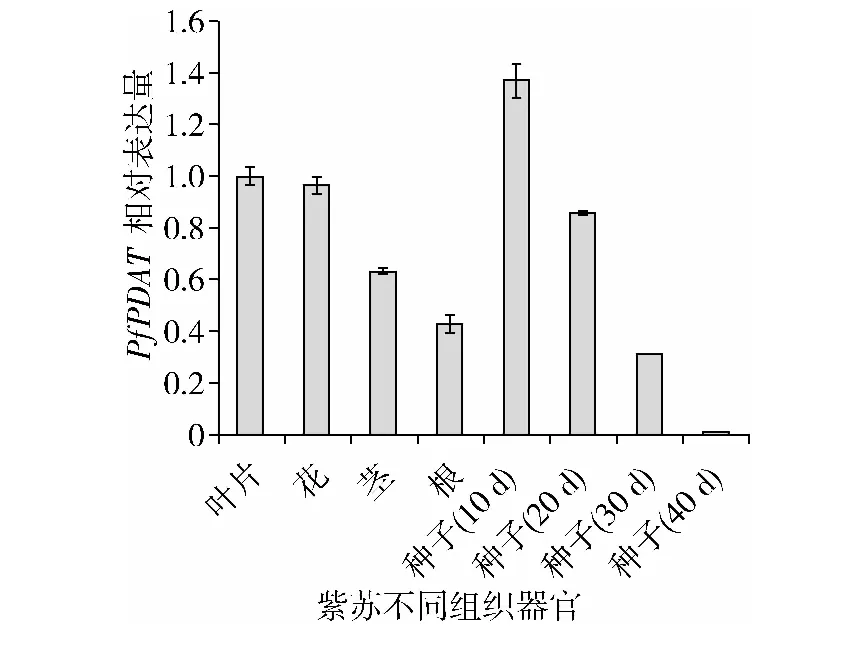
圖5 晉蘇1號不同組織器官PfPDAT相對表達量的變化
以葉片最高,莖和根系中表達量比較低,種子發育初期(開花后10 d)表達量最高,是葉片表達量的1.4倍,隨著種子逐漸成熟,PfPDAT基因呈下調表達,到開花后40 d,該基因表達量僅為種子發育初期的0.006倍。
3 討論與結論
通過氣相色譜分析得出紫蘇種子中主要含有棕櫚酸(C16∶0)、硬脂酸(C18∶0)、油酸(C18∶1)、亞油酸(C18∶2)、亞麻酸(C18∶3)5種脂肪酸,其中不飽和脂肪酸含量高,占總脂肪酸的90.5%~92.4%。晉蘇1號α-亞麻酸含量高,可達到456.6 mg/g,亞麻酸與亞油酸之比為9∶1,是高含α-亞麻酸的優質品種。紫蘇種子發育過程中各脂肪酸組分隨著種子發育呈現一定變化規律,飽和脂肪酸如棕櫚酸、硬脂酸含量較為穩定,不飽和脂肪酸如油酸含量隨著種子發育,前期積累量增長緩慢,后期顯著增加,而α-亞麻酸含量不斷增加直至種子成熟,亞油酸含量呈連續下降趨勢,這可能與種子發育過程中大量亞油酸向 α-亞麻酸快速轉化積累有關,而亞麻酸含量的增加抑制了亞油酸的合成,從而導致ALA與LA之間的比例差距加大[11]。從總體來看,脂肪酸累積過程較長,在進入開花期后需要30~40 d才能完成。本試驗研究發現隨著種子的發育,種子鮮重呈先升高后降低的趨勢,開花后30 d達到最大,種子形態也由多水透明狀轉變為乳白或深紫色最后變成褐色,同時種子逐漸變得硬實,脂肪酸的積累量迅速增加,到種子發育后期脂肪酸含量達到最大。
α-亞麻酸等脂肪酸合成后,經一系列酶促反應,最終轉入甘油-3-磷酸分子(glycerol-3 phosphate, G3P)而生成三酰基甘油(TAG),貯存于油體中。研究表明,PDAT 和DGAT1 是控制模式植物擬南芥種子油合成積累的關鍵酶[12-13]。王安可[14]在棉花中的研究發現在油脂儲存的主要器官種子中DGAT1表達量較高。Wang等[15]提取開花后10、20和30 d的大豆豆莢RNA,檢測了GmDGAT1基因的表達情況,發現隨著豆莢的發育表達量遞增,且在30 d時,基因的表達量達到最高。蓖麻種子中的DGAT1的表達量是從種子發育開始逐漸上升,在中期達到最大[16]。有研究報道PDAT 能選擇性積累羥化脂肪酸[17]。本研究對紫蘇不同器官及種子發育不同時期PfDGAT1及PfPDAT基因表達特性進行分析,結果表明,二者在種子中均大量表達,PfDGAT1表達量呈現先升高后降低的趨勢,在開花后30 d,表達量最高,緊接PfDGAT1基因高峰表達期的是α-亞麻酸和總油脂快速積累期。這表明PfDGAT1在紫蘇α-亞麻酸和總油脂積累過程中行使重要功能。
[1]Lee J E, SEong J E, Hong S H, et al. Inhibitory effect of perilla oil on hepatic lipid accumulation in the apoE knock-out mice fed high cholesterol diet[J]. Proceedings of the Nutrition Society, 2011, 70(4) : 142
[2]Chang H H, Chen C S, Lin J Y. Dietary perilla oil lowers serum lipids and ovalbumin-specific lgG1, but increases total lgE levels in ovalbumin-challenged mice[J]. Food and Chemical Toxicology, 2009, 47(4) : 848-850
[3]岳愛琴,李潤植.新型可再生工業用油脂的代謝工程[J].中國生物化學與分子生物學報, 2009, 25(6) : 501-509
[4]Li R Z, Yu K S, Hatanaka T, et al. Vernonia DGATs increase accumulation of epoxy fatty acids in oil[J]. Plant Biotechnology Journal, 2010, 8 : 184-195
[5]Clemente T E, Cahoon E B. Soybean oil: genetic approaches for modification of functionality and total content[J]. Plant Physiology, 2009, 151 : 1030-1040
[6]Durrentt T P, Benning C, Ohlrogge J. Plant triacylglycerols as feedstocks for the production of biofuels[J]. The Plant Journal, 2008, 54 : 293-601
[7]Dyer J M, Mullen R T. Engineering plant oils as high-value industrial feedstocks for biorefining: the need for underpinning cell biology research[J]. Physiologia Plantarum, 2008, 132 :11-22
[8]張志軍,孫子文,李會珍,等.不同品種紫蘇種子營養成分及脂肪酸組分分析[J].中國糧油學報, 2012, 27(8) :57-59
[9]張洪,黃建韶,趙東海.紫蘇營養成分的研究[J].湖南文理學院學報:自然科學版, 2006, 18(1) : 49-51
[10]Longvah T, Deosthale Y G, Uday Kumar P. Nutritional and short term toxicological evaluation of Perilla seed oil[J]. Food Chemistry, 2000, 70 : 13-16
[11]Ichihara K, Suda Y. Lipid biosynthesis in developing perilla seeds[J]. Phytochemistry, 2003, 63 : 139-143
[12]Zhang M, Fan J, Taylor D C, et al. DGAT1 and PDAT1 Acyltransferases have overlapping functions in Arabidopsis triacylglycerol biosynthesis and are essential for normal pollen and seed development[J]. Plant Cell, 2009, 21:3885-3901
[13]Shockey J M, Gidda S K, Chapital D C, et al. Tung tree DGAT1 and DGAT2 have nonredundant functions in triacylglycerol biosynthesis and are localized to different subdomains of the endoplasmic reticulum[J]. Plant Cell, 2006,18:2294-2313
[14]王安可.VgDGAT1基因在棉花中的遺傳轉化和表達研究[D].杭州: 浙江大學, 2013
[15]Wang H W, Zhang J S, Gai J Y, et al. Cloning and comparative analysis of the gene encoding diacylglycerol acyltransferase from wild type and cultivated soybean[J]. Theoretical and Applied Genetics, 2006, 112(6) : 1086-1097
[16]He X H, Turner C, Chen G Q, et al. Cloning and characterization of a cDNA encoding diacylglycerol acyltransferase from castor bean[J]. Lipids, 2004, 39(4) : 311-318
[17]van Erp H, Bate P G, Burgal J, et al. Castor phospholipid: diacylglycerol acyltransferase facilitates efficient metabolism of hydroxyl fatty acid in transgenic Arabidopsis[J]. Plant Physiology, 2011, 155(2) : 683-693.
Regulation of Controlling Oil Synthesis by Fatty Acid Metabolism of Perilla Seed and Key Enzyme Gene
Wang Jiping Zhang Linghui Zhao Jing Wang Yanzun Li Runzhi
(College of Agronomy, Shanxi Agricultural University, Taigu 030801)
In order to study the biosynthesis mechanism of grease and fatty acids in seeds, especially the high-level accumulation of α- linolenic acid, this paper adopted gas chromatography method to analyze composition, content and dynamic change of fatty acid in the mature seeds of seven perrila varieties and Jinsu No. 1 seed at different development stages; the developing seeds so as to investigate physiological-biochemical features of perilla seed oil and fatty acid biosynthesis. Expression pattern of key genes in fatty acid metabolism was analyzed. The results showed that unsaturated fatty acids (18∶1, 18∶2, 18∶3) made up 90% of total seed oil in perilla. Following seed developing, seed fresh weight and total fatty acid content were increased, and seed morphology was also changed from aqueous and transparent forms to milky white or dark purple and finally brown matured seeds. During seed development and maturation, linoleic (18∶2) content was gradually reduced while the level of α-linolenic acid (18∶3) was correspondingly increased up to more than 60% of the total fatty acids in the mature seeds. Of seven perilla varieties tested, variety Jinsu1 contained the highest level of α-linolenic acid (456.6 mg/g). The ratio of α-linolenic acid to linoleic acid is 9∶1. Real-time PCR analysis showed that small expression ofPfDGAT1 andPfPDATwas in root and stems while large expression of the genes was in leaves and seeds. Following seed development,PfDGAT1 expression increased slowly at the earlier stages, and then increased vastly at mid stages and reduced slowly at late stages, with the peak expression level in the seed at 30 days after flowering. ThePfDGAT1 highly expression stage was just ahead of the high accumulation stage of α-linolenic acid and total oil in seed development, indicating thatPfDGAT1 function importantly in perilla seed oil and α-linolenic acid accumulation.
PerillafrutescensL., seed oil, fatty acid, diacylglycerol acyltransferase enzyme (DGAT1), phospholipids diacylglycerol acyltransferase (PDAT)
S565.8
A
1003-0174(2016)03-0091-05
時間:2015-12-28 17:06:54
網絡出版地址:http://www.cnki.net/kcms/detail/11.2864.TS.20151228.1706.002.html
國家青年科學基金(31201266)
2014-12-26
王計平,女,1974年出生,博士,副教授,植物生理與分子生物學
李潤植,男,1959年出生,博士,教授,植物分子遺傳與基因工程
猜你喜歡
食品工業(2023年12期)2023-12-12 12:56:16
糧油食品科技(2022年6期)2022-11-25 12:18:34
糧油食品科技(2022年6期)2022-11-25 12:18:30
基層中醫藥(2020年8期)2020-11-16 00:55:20
環境衛生工程(2020年3期)2020-07-27 01:19:22
瘋狂英語·新讀寫(2020年3期)2020-06-06 09:06:14
塑料助劑(2019年3期)2019-07-24 08:51:22
中國糧油學報(2019年4期)2019-07-12 09:06:38
食品與機械(2018年11期)2019-01-03 06:15:34
中國糧油學報(2018年12期)2018-01-26 13:32:04
