旱麥草EtAP2基因的克隆及其對干旱脅迫的響應(yīng)表達(dá)
2017-01-03 01:39:12王夢瑤周茜萍呂新華
西北植物學(xué)報 2016年11期
關(guān)鍵詞:植物
王夢瑤,周茜萍,呂新華,馮 麗,孫 黎
(石河子大學(xué) 生命科學(xué)學(xué)院,農(nóng)業(yè)生物技術(shù)重點實驗室,新疆 石河子 832003)
旱麥草EtAP2基因的克隆及其對干旱脅迫的響應(yīng)表達(dá)
王夢瑤,周茜萍,呂新華,馮 麗,孫 黎*
(石河子大學(xué) 生命科學(xué)學(xué)院,農(nóng)業(yè)生物技術(shù)重點實驗室,新疆 石河子 832003)
以旱麥草(Eremopyrumtriticeum)為實驗材料,利用RT-PCR技術(shù)從旱麥草葉片中克隆了1個AP2/ERF家族基因,命名為EtAP2(GenBank登錄號KX622583)。EtAP2基因含有1 128 bp開放閱讀框,編碼375個氨基酸,相對分子質(zhì)量40.87 kD,等電點為5.36。多序列比對和進(jìn)化樹分析表明,該基因編碼蛋白具有2個AP2保守結(jié)構(gòu)域,與小麥AP2/ERF家族蛋白具有較近的親緣關(guān)系。實時熒光定量PCR分析表明,15% PEG 6000模擬干旱脅迫可誘導(dǎo)EtAP2基因在根和葉中表達(dá),且在根中對干旱脅迫的響應(yīng)大于葉片。研究表明,EtAP2可能參與旱麥草對干旱逆境脅迫應(yīng)答的調(diào)節(jié)。
旱麥草;EtAP2;克隆;基因表達(dá)
AP2/ERF (APETALA2/Ethylene-Responsive Factor,乙烯響應(yīng)因子)是植物最大的基因家族之一,編碼植物特有的轉(zhuǎn)錄因子。AP2/ERF超基因家族以AP2/ERF結(jié)構(gòu)域為特征,該結(jié)構(gòu)域由60~70個氨基酸組成,與DNA結(jié)合有關(guān)。AP2/ERF超基因家族包括AP2、RAV、DREB和ERF亞家族[1]。AP2亞家族又分為AP2和ANT組,ANT組又可細(xì)分為ANT、AIL以及PLT組[2]。AP2亞家族成員蛋白通常包含多個AP2/ERF結(jié)構(gòu)域,RAV亞家族轉(zhuǎn)錄因子包含一個AP2/ERF結(jié)構(gòu)域和B3結(jié)構(gòu)域,DREB和ERF亞家族蛋白僅含有一個AP2/ERF結(jié)構(gòu)域[3]。AP2/ERF基因家族成員在調(diào)節(jié)植物發(fā)育、抵御生物和非生物脅迫中起重要作用[4]。隨著第二代測序技術(shù)在植物研究中的應(yīng)用,AP2/ERF轉(zhuǎn)錄因子已在多種植物中得到鑒定,如擬南芥[5]、小麥[6]、水稻[7]和玉米[8]等。但目前為止,有關(guān)旱麥草(Eremopyrumtriticeum)AP2/ERF家族基因的鑒定和特征研究尚未見報道。
旱麥草是禾本科小麥族旱麥草屬早春短生育期植物,小麥的野生近緣種,主要分布在準(zhǔn)噶爾盆地沙區(qū)荒漠中,具有生育期短、特早熟、抗旱、耐鹽、分蘗力強(qiáng)、抗白粉病以及光和效率高等優(yōu)良性狀。本研究以旱麥草為研究對象,根據(jù)前期構(gòu)建的干旱脅迫下的轉(zhuǎn)錄組數(shù)據(jù)庫,對旱麥草AP2/ERF轉(zhuǎn)錄因子家族成員EtAP2基因進(jìn)行了克隆、序列分析和干旱脅迫下的表達(dá)特征研究,以期為后續(xù)研究該基因的功能奠定基礎(chǔ)。
1 材料和方法
1.1 材 料
旱麥草種子由本實驗室成員于2015年6月采自新疆準(zhǔn)噶爾盆地荒漠。
1.2 方 法
1.2.1 材料的處理 2016年6月將采集的旱麥草種子種植于石河子大學(xué)生命科學(xué)學(xué)院溫室。將生長2周的旱麥草幼苗用15% PEG 6000[9]分別處理0.5、1、3、6、12、24和48 h,以未處理的旱麥草幼苗為對照,取其根系和葉片樣品液氮速凍后儲存于-80 ℃冰箱。
1.2.2 旱麥草EtAP2基因的克隆 采用RNA prep Pure Plant Kit試劑盒(Tiangen,北京)提取旱麥草幼苗葉片總RNA,用PrimeScriptTMReverse Transcriptase(TaKaRa,大連)將總RNA反轉(zhuǎn)錄成cDNA。根據(jù)課題組通過旱麥草轉(zhuǎn)錄組測序獲得的 EST 序列(GenBank登錄號為KX622583),利用Primer Premier 5.0軟件設(shè)計引物EtAP2-F和EtAP2-R(表1),PCR擴(kuò)增該序列的開放閱讀框(ORF)。PCR反應(yīng)條件為:94 ℃預(yù)變性3 min,94 ℃變性30 s,55 ℃退火30 s,72 ℃延伸30 s,30個循環(huán),72 ℃延伸8 min。產(chǎn)物回收純化后與 pMD19-T載體連接,轉(zhuǎn)化大腸桿菌Trans5α,經(jīng)菌落PCR驗證后送生工生物工程(上海)股份有限公司測序。
1.2.3EtAP2基因的生物信息學(xué)分析 序列比對和保守結(jié)構(gòu)預(yù)測使用DNAMAN 5.0和Blast(http://blast.ncbi.nlm.nih.gov/Blast.cgi)進(jìn)行。蛋白質(zhì)基本性質(zhì)分析使用EXPASY(http://web.expasy.org/protparam/)相關(guān)在線工具完成。系統(tǒng)進(jìn)化樹構(gòu)建使用MEGA 4.0 軟件進(jìn)行[10]。蛋白質(zhì)三級結(jié)構(gòu)使用Swiss-Model (http:/ /swissmode.l expasy.org /) 軟件預(yù)測。
1.2.4 旱麥草EtAP2基因表達(dá)性分析 采用實時熒光定量PCR(qRT-PCR)技術(shù)檢測EtAP2基因在干旱脅迫下的轉(zhuǎn)錄表達(dá)水平。基因特異引物為EtAP2-RT-F和EtAP2-RT-R,以旱麥草18S rRNA基因為內(nèi)參,引物為Et18S-F和Et18S-R(表1)。按照SYBR Premix ExTaqTM試劑盒(TAKARA)說明書,在羅氏LightCycler480 PCR儀進(jìn)行qRT-PCR,分析EtAP2基因相對表達(dá)量。擴(kuò)增程序為:95 ℃預(yù)變性10 min,95 ℃變性10 s,56 ℃退火30 min,72 ℃延伸32 s,40個循環(huán)。每個樣品做3次重復(fù),以未處理旱麥草為對照。數(shù)據(jù)采用2-ΔΔCt法計算。
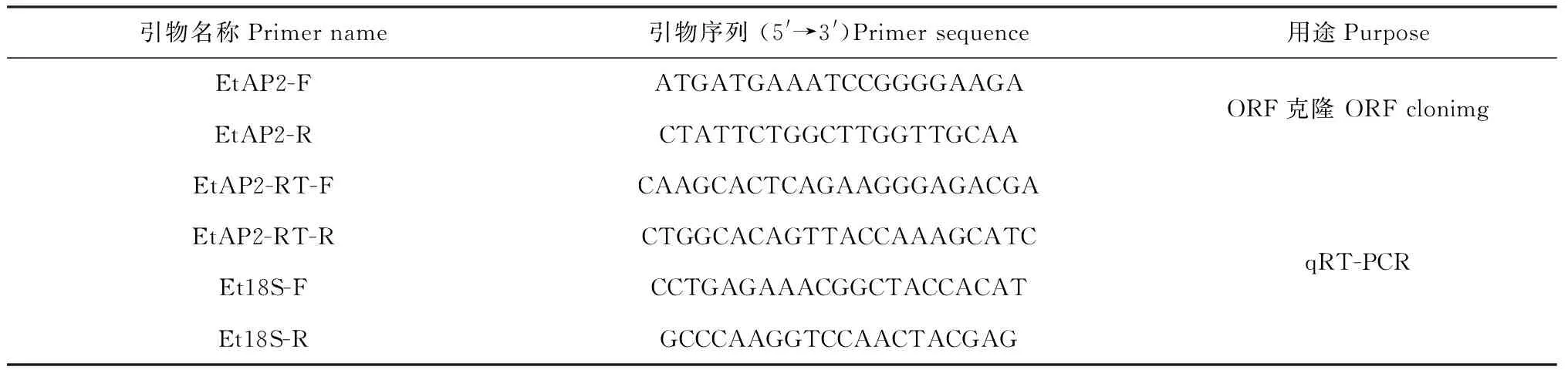
表1 本實驗用到的引物
2 結(jié)果與分析
2.1 EtAP2基因克隆及序列分析
根據(jù)旱麥草轉(zhuǎn)錄組測序結(jié)果設(shè)計引物EtAP2-F和EtAP2-R,通過RT-PCR擴(kuò)增出一個約1.1 kb片段,與預(yù)期大小相同(圖1)。進(jìn)一步測序分析表明,該片段含有1 128 bp開放閱讀框,編碼375個氨基酸(圖2),Blast比對結(jié)果表明,該ORF編碼的氨基酸序列與小麥TaAP2氨基酸序列相似性最高為96%,因此將該基因命名為EtAP2(GenBank登錄號為KX622583)。
為了解EtAP2轉(zhuǎn)錄因子的生物活性及潛在功能,利用EXPASY在線分析工具ProtParam對翻譯得到的EtAP2氨基酸組成和理化性質(zhì)進(jìn)行分析和預(yù)測。結(jié)果顯示:EtAP2相對分子質(zhì)量為40.87 kD,等電點為5.36,酸性氨基酸為16個,堿性氨基酸15個,脂肪族氨基酸含量占89%,而芳香族氨基酸只占6.2%。保守結(jié)構(gòu)域分析發(fā)現(xiàn), 該蛋白在191~262與292~354位氨基酸之間含有2個典型的AP2結(jié)構(gòu)域(圖3),與其他植物AP2/ERF家族轉(zhuǎn)錄因子的AP2保守結(jié)構(gòu)域相似度較高,表明EtAP2屬于AP2亞家族。AP2結(jié)合域由大約60個氨基酸組成,是非常保守的DNA結(jié)合區(qū),在其N-端有一個堿性親水區(qū),具有3個反平行的β-折疊,這3個β-折疊在識別順式作用元件中起到重要的作用[11]。利用protscale工具對EtAP2進(jìn)行了親水性和疏水性分析,結(jié)果表明,EtAP2大部分氨基酸為親水性氨基酸,屬于親水性蛋白。
利用BlastP檢索EtAP2同源蛋白,獲得幾個相似度較高的AP2/ERF氨基酸序列,包括小麥(Triticumaestivum,AK334560)、大麥(Hordeumvulgare,AK252982)、二穗短柄草(Brachypodiumdistachyon,XM_014901079.1)、玉米(Zeamays,NM_001138224.1)、谷子(Setariaitalica,XM_004953799.2)和短花藥野生稻(Oryzabrachyantha,XM_015833886)。DNAMAN5.0比對結(jié)果(圖3)表明,EtAP2轉(zhuǎn)錄因子與小麥AP2/ERF相似性最高(96%),與玉米的相似性較低(83%)。
2.2 EtAP2系統(tǒng)進(jìn)化分析
為研究旱麥草EtAP2與其他植物同源蛋白之間的進(jìn)化關(guān)系,通過 MEGA 4.0構(gòu)建了EtAP2與小麥、水稻等物種AP2/ERF家族轉(zhuǎn)錄因子的系統(tǒng)進(jìn)化樹(圖4)。結(jié)果表明,EtAP2轉(zhuǎn)錄因子在進(jìn)化關(guān)系上屬于AP2 /ERF家族轉(zhuǎn)錄因子AP2亞家族中的AIL組,與小麥、山羊草等禾本科植物的AP2蛋白聚為一類,其中與小麥AP2蛋白(AK334560)的親緣關(guān)系最近,屬于同一進(jìn)化分支。
2.3 EtAP2轉(zhuǎn)錄因子的三級結(jié)構(gòu)預(yù)測與分析
利用Swiss model程序?qū)tAP2進(jìn)行蛋白質(zhì)三級結(jié)構(gòu)同源建模分析(圖5),結(jié)果表明該蛋白與擬南芥AP2/ERF家族AtERF1蛋白的三級結(jié)構(gòu)相似性為 44.26%。該結(jié)構(gòu)中存在一個由色氨酸、絲氨酸、谷氨酸、組氨酸組成的保守序列。2個AP2結(jié)構(gòu)域位于DNA的大溝中,都存在一個接頭區(qū),大概由30個氨基酸組成[12]。在與目的基因結(jié)合過程中,前一個AP2結(jié)構(gòu)域與5′-C端結(jié)合,后一個與3′-N端結(jié)合[13]。
2.4 EtAP2基因在模擬干旱脅迫下的表達(dá)分析
為了研究EtAP2基因?qū)Ω珊得{迫的響應(yīng),利用qRT-PCR方法檢測了其在旱麥草根和葉片中的表達(dá)量(圖6)。在15% PEG6000模擬干旱脅迫處理下,EtAP2在葉中的表達(dá)變化相對平緩,在0.5 h有輕微下降,1 h時輕微上升,3 h表達(dá)量最低,之后在6 h達(dá)到最高水平,為對照的1.7倍,在12~48 h呈現(xiàn)降-升-降的趨勢。在干旱脅迫處理48 h之內(nèi),EtAP2在根中的表達(dá)量都高于對照,3 h達(dá)到最大,是對照的21倍,之后在6 h又急劇下降,直到48 h表達(dá)水平最低。這些結(jié)果表明,在干旱處理條件下,EtAP2的表達(dá)水平被顯著誘導(dǎo),且在根系中對干旱的響應(yīng)大于葉片。
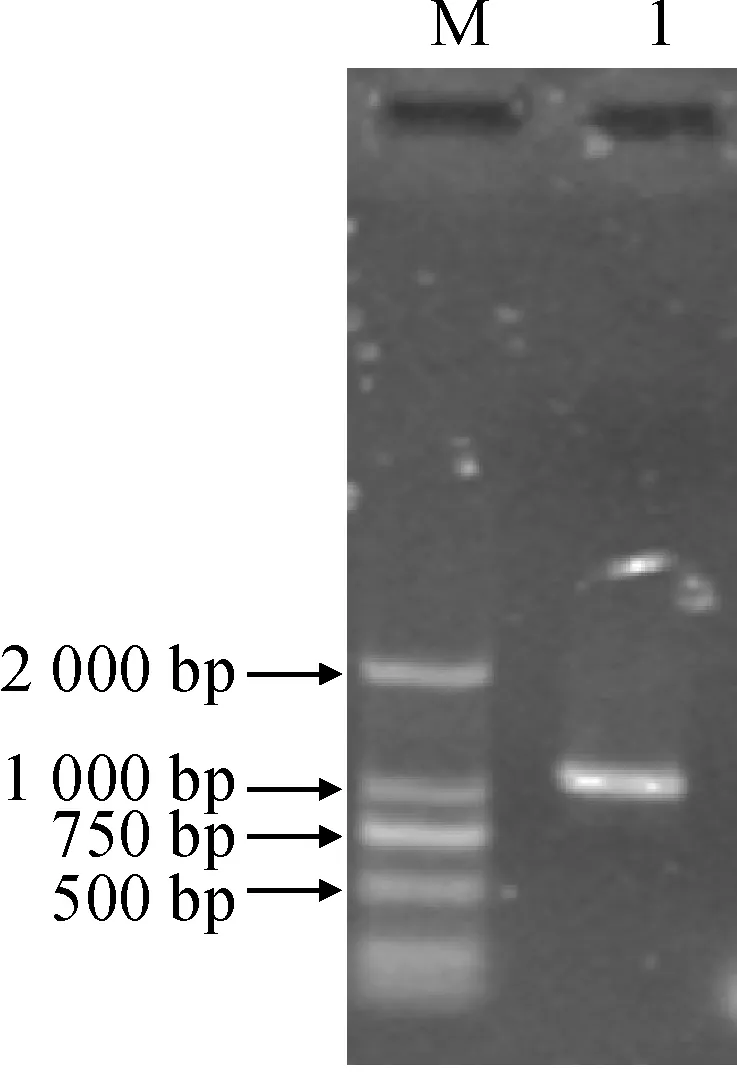
M. DL2000; 1. PCR擴(kuò)增產(chǎn)物圖1 EtAP2基因的RT-PCR擴(kuò)增M. DL2000; 1. Product of PCRFig.1 RT-PCR application of EtAP2
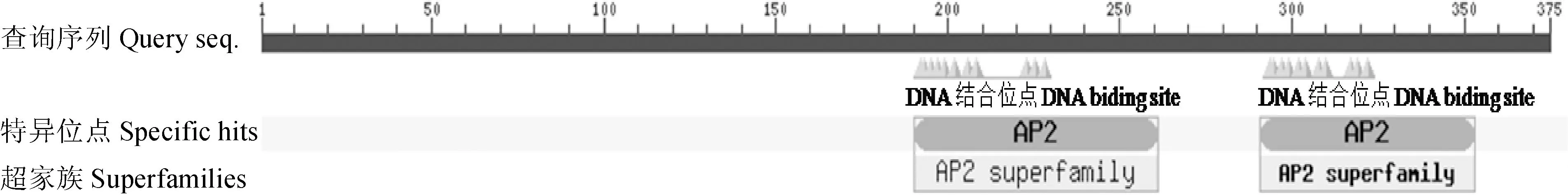
圖2 EtAP2蛋白結(jié)構(gòu)域分析Fig.2 Protein domain analysis of EtAP2

AK334560. 小麥;AK252982. 大麥;XM_014901079.1. 二穗短柄草;XM_015833886. 短花藥野生稻;XM_004953799.2. 谷子;NM_001138224.1. 玉米圖3 旱麥草EtAP2蛋白序列的多重比對AK334560. Triticum aestivum;AK252982. Hordeum vulgare;XM_014901079.1. Brachypodium distachyon;XM_015833886. Oryza brachyantha;XM_004953799.2. Setaria italica;NM_001138224.1. Zea maysFig.3 Multi-alignment of EtAP2 protein with AP2 proteins in plants

圖中各節(jié)處數(shù)字表示重復(fù)1 000次Bootstrap值,括號內(nèi)為氨基酸登陸號圖4 旱麥草EtAP2與其他AP2蛋白的系統(tǒng)進(jìn)化樹The numbers at note represent the bootstrap values with 1 000 replicates in this figure, and the accession number of each sequence is given in the bracketFig.4 Phylogenetic analysis of EtAP2 and other plant AP2 proteins
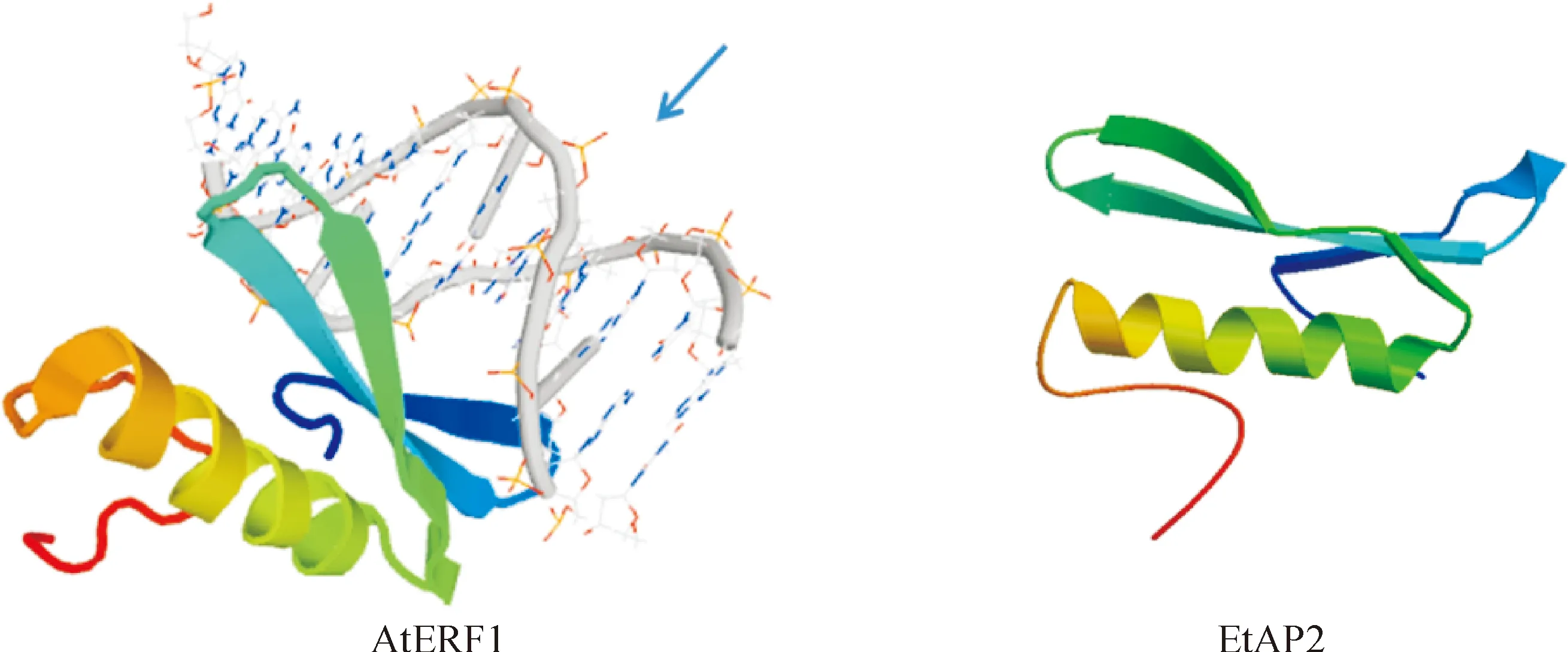
箭頭所指方向為DNA分子圖5 AtERF1和 EtAP2轉(zhuǎn)錄因子DNA結(jié)合域三維結(jié)構(gòu)建模The arrow head is DNAFig.5 Three-dimensional structure modeling of DNA-binding domain of EtAP2 and AtERF1 protein
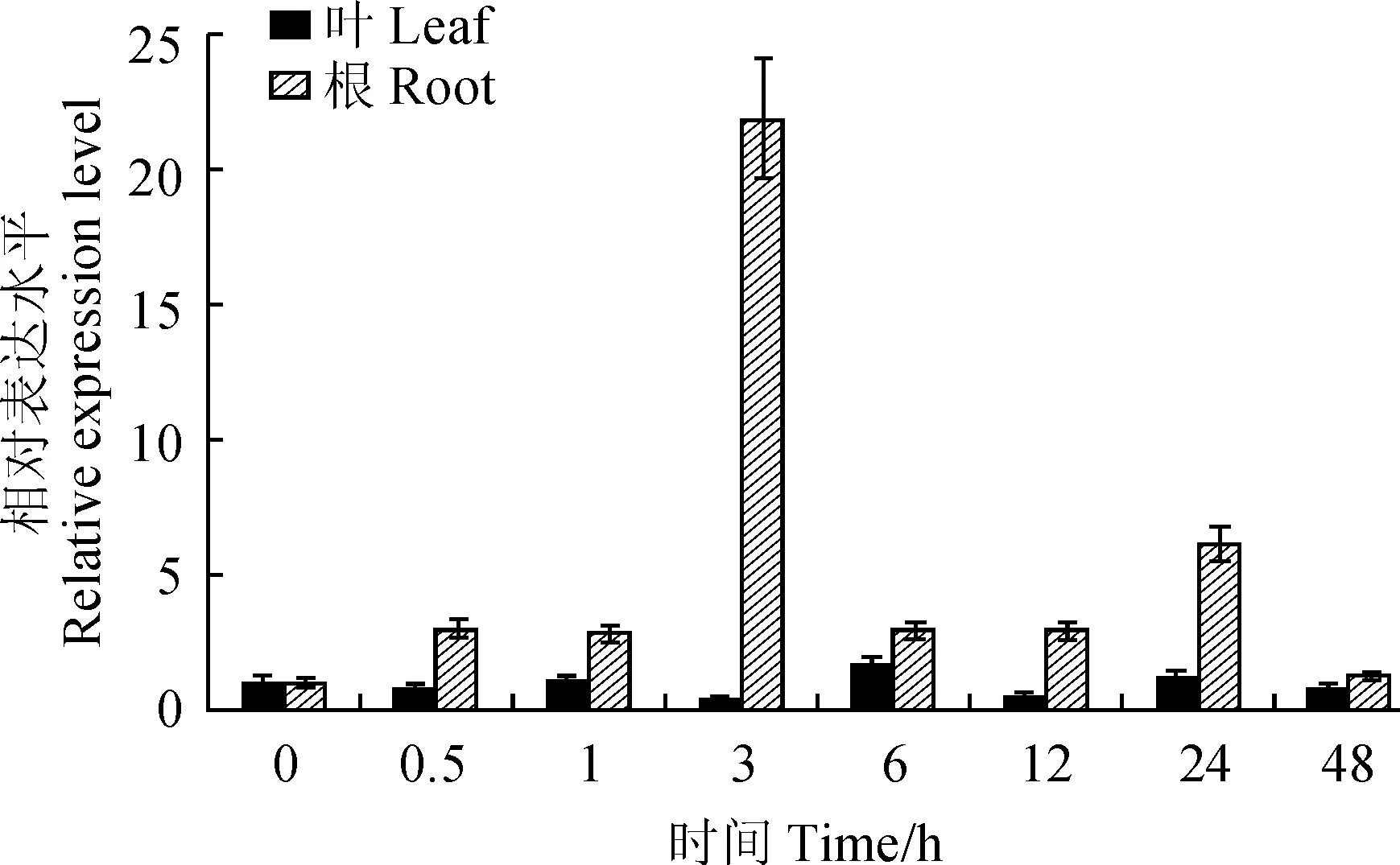
圖6 EtAP2基因在模擬干旱脅迫下在旱麥草幼苗根和葉中的表達(dá)Fig.6 Expression of EtAP2 in roots and leaves of E. triticeum seedlings under simulated drought stress
3 討 論
植物在干旱等逆境脅迫下會在生理和基因水平上進(jìn)行自身調(diào)整,以適應(yīng)外界環(huán)境。AP2/ERF作為重要的轉(zhuǎn)錄因子,在植物自身調(diào)節(jié)中起到非常關(guān)鍵的作用,能夠快速調(diào)節(jié)植物體內(nèi)相關(guān)基因的表達(dá)來提高植物對逆境脅迫的耐受性[14]。旱麥草分布在干旱、瘠薄荒漠地區(qū),生長發(fā)育過程會受到各種非生物因素的影響,研究其抗逆相關(guān)基因并進(jìn)行功能分析,有助于從分子機(jī)制上進(jìn)一步了解其適應(yīng)逆境的機(jī)理,同時為小麥等農(nóng)作物的抗逆性狀改良提供候選基因。
AP2/ERF家族是植物普遍存在的一類轉(zhuǎn)錄因子,廣泛參與植物逆境脅迫誘導(dǎo)[15]。許多研究表明,過量表達(dá)某些AP2/ERF轉(zhuǎn)錄因子基因會增強(qiáng)植物的抗旱性。Dossa等[16]對芝麻進(jìn)行轉(zhuǎn)錄組分析后,發(fā)現(xiàn)一個AP2si16基因,可以作為增強(qiáng)芝麻耐旱性的候選基因之一。Zhang等[17]從大豆中克隆一個GmERF3基因,并將其在煙草中過表達(dá),在干旱條件下,轉(zhuǎn)基因煙草中游離脯氨酸和可溶性碳水化合物的含量明顯高于非轉(zhuǎn)基因煙草,顯著增加了煙草對干旱和高鹽的耐受性。Zeng等[18]對AP2/ERF家族蛋白進(jìn)行了系統(tǒng)分析,發(fā)現(xiàn)該家族蛋白主要參與植物的生長發(fā)育和對各種脅迫的應(yīng)激反應(yīng),包括光形態(tài)建成,花的發(fā)育,干旱和寒冷的應(yīng)激反應(yīng)以及對生長素和脫落酸的敏感性等。
本研究以旱麥草為研究對象,從干旱脅迫的幼苗中克隆了EtAP2基因的cDNA,該基因編碼的氨基酸序列具有AP2/ERF轉(zhuǎn)錄因子家族的基本特征,都含有AP2結(jié)合域。系統(tǒng)進(jìn)化樹分析結(jié)果表明,EtAP2轉(zhuǎn)錄因子與小麥親緣關(guān)系最近,都屬于AP2亞族。本研究中qRT-PCR分析表明,EtAP2的表達(dá)量受干旱誘導(dǎo),并且在根中的表達(dá)對干旱脅迫的響應(yīng)程度大于葉片,說明EtAP2基因在干旱脅迫條件下可能參與基因表達(dá)調(diào)控,可以作為小麥等農(nóng)作物抗性育種重要的候選基因。這些研究將有助于了解AP2/ERF轉(zhuǎn)錄因子EtAP2在旱麥草逆境調(diào)控中的作用,為進(jìn)一步研究和解析AP2/ERF家族其他成員的功能提供參考。
[1] WESSLER S R. Homing into the origin of the AP2 DNA binding domain [J].TrendsinPlantScience, 2005,10: 54-56.
[2] TANG M F, LI G S, CHEN M S. The phylogeny and expression pattern of APETALA2-like genes in rice [J].JournalofGeneticsandGenomics, 2007,34(10): 930-938.
[3] SAKUMA Y, LIU Q, DUBOUZET J G,etal. DNA-binding specificity of the ERF/AP2 domain of Arabidopsis DREBs, transcription factors involved in dehydration- and cold-inducible gene expression[J].BiochemicalandBiophysicalResearchCommunications, 2002,290: 998-1 009.
[4] LIU S, WANG X, WANG H,etal. Genome-wide analysis ofZmDREBgenes and their association with natural variation in drought tolerance at seedling stage ofZeamaysL [J].PlosGenetics, 2013,9(9):119-129.
[5] OHTO M A, FLOYD S K, FISCHER R L,etal. Effects of APETALA2 on embryo, endosperm, and seed coat development determine seed size inArabidopsis[J].SexualPlantReproduction, 2009,22: 277-289.
[6] ZHU X L, QI L, LIU X,etal. The wheat ethylene response factor transcription factor pathogen-induced ERF1 mediates host responses to both the necrotrophic pathogenRhizoctoniacerealisand freezing stresses[J].PlantPhysiology, 2014,164(3): 1 499-1 514.
[7] NAKANO T, SUZUKI K, FUJIMURA T,etal. Genome-wide analysis of the ERF gene family inArabidopsisand rice [J].PlantPhysiology, 2006,140: 411-432.
[8] DU H, HUANG M, ZHANG Z,etal. Genome-wide analysis of the AP2/ERF gene family in maize waterlogging stress response [J].Euphytica, 2014,198(1): 115-126.
[9] HIROSHI M, SHINJIRO Y, ATSUSHI H,etal. Dwarf and delayed-flowering 1, a novelArabidopsismutant deficient in gibberellin biosynthesis because of overexpression of a putative AP2 transcription factor [J].ThePlantJournal, 2004,37(5): 720-729.
[10] TAMURA K, DUDLEY J, NEI M,etal. MEGA4: molecular evolutionary genetics analysis (MEGA) software version 4.0 [J].MolecularBiologyEvolution, 2007,24(8): 1 596-1 599.
[11] 張計育, 王慶菊, 郭忠仁. 植物AP2/ERF 類轉(zhuǎn)錄因子研究進(jìn)展[J]. 遺傳, 2012,34(7): 835-847.
ZHANG J Y, WANG Q J, GUO Z R. Progresses on plant AP2/ERF transcription factors[J].Hereditas, 2012,34(7): 835-847.
[12] RIECHMANN J L, MEYEROWITZ E M. The AP2/EREBP family of plant transcription factors [J].JournalofBiologicalChemistry, 1998, 379(6): 633-646.
[13] NOLE-WILSON S, KRIZEK B A. DNA binding properties of theArabidopsisfloral development protein AINTEGUMENTA [J].NucleicAcidsResearch, 2000,28(21): 4 076-4 082.
[14] HUIJUAN Z, YONGBO H, LEI H,etal. Arabidopsis AtERF014 acts as a dual regulator that differentially modulates immunity againstPseudomonassyringaepv.tomato andBotrytiscinerea[J].ScientificReports, 2016,6:30 251.
[15] GOLLDACK D, LI C, MOHAN H, PROBST N. Tolerance to drought and salt stress in plants: unraveling the signaling networks [J].FrontiersinPlantScience, 2014,5:151.
[16] DOSSA K, WEI X, LI D,etal. Insight into the AP2/ERF transcription factor superfamily in sesame and expression profiling of DREB subfamily under drought stress[J].BMCPlantBiology, 2016,16(1): 171.
[17] ZHANG G Y, CHEN M, LI L C,etal. Overexpression of the soybeanGmERF3 gene, an AP2/ERF type transcription factor for increased tolerances to salt, drought, and diseases in transgenic tobacco [J].JournalofExperimentalBotany, 2009,60(13): 3 781-3 796.
[18] ZENG L, YIN Y, YOU C,etal. Evolution and protein interactions of AP2 proteins in Brassicaceae: evidence linking development and environmental responses [J].JournalofIntegrativePlantBiology, 2016,58(6): 549-563.
(編輯:宋亞珍)
Cloning and Expression Response to Simulated Drought Stress ofEtAP2 fromEremopyrumtriticeum
WANG Mengyao, ZHOU Xiping, Lü Xinhua, FENG Li, SUN Li*
(Key Laboratory of Agricultural Biotechnology, College of Life Sciences, Shihezi University, Shihezi, Xinjiang 832003, China)
A AP2/ERF gene was isolated fromEremopyrumtriticeumby RT-PCR method, and designated asEtAP2 (GenBank accession no. KX622583).EtAP2 contained an open reading frame (ORF) of 1 128 bp encoding 375 amino acids. The molecular mass of EtAP2 was 40.87 kDa with isoelectric point (PI) of 5.36. Multiple sequences alignment and phylogenetic analysis indicated that EtAP2 protein contained two conserved AP2 domains, which had close genetic relationship with AP2/ERF transcription factor inTriticumaestivum. Quantitative real-time PCR analysis revealed that simulated drought stress (15% PEG6000) treatment significantly induced the expression ofEtAP2, and root is more sensitive to drought stress than leaf. The results indicated thanEtAP2 might involve in response to drought stress regulation inE.triticeum.
Eremopyrumtriticeum;EtAP2; cloning; gene expression
1000-4025(2016)11-2167-06
10.7606/j.issn.1000-4025.2016.11.2167
2016-09-16;修改稿收到日期:2016-11-01
石河子大學(xué)高層次人才科研啟動資金(RCZX201139)
王夢瑤(1993-),男,在讀碩士生,主要從事植物分子生物學(xué)研究。E-mail:1561855244@qq.com
*通信作者:孫 黎,博士,副教授,主要從事植物分子生物學(xué)研究。E-mail: sunlishz@126.com
Q785;Q786
A
猜你喜歡
少兒科學(xué)周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學(xué)閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數(shù)英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數(shù)英綜合(2020年11期)2020-12-16 02:57:22
學(xué)苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
