TLR2/4在MTB Hsp16.3刺激小鼠巨噬細胞過程中表達及作用的初步探討①
2017-02-15 09:43:36李姍姍劉芊伊張繼東馮繼紅李龍梅泮紅飛羅軍敏
中國免疫學雜志 2017年1期
李姍姍 秦 歡 劉芊伊 徐 林 張繼東 馮繼紅 李龍梅 泮紅飛 羅軍敏
(遵義醫(yī)學院免疫學教研室,貴州省免疫分子應用研究工程中心,遵義563099)
TLR2/4在MTB Hsp16.3刺激小鼠巨噬細胞過程中表達及作用的初步探討①
李姍姍 秦 歡 劉芊伊 徐 林 張繼東 馮繼紅 李龍梅 泮紅飛 羅軍敏
(遵義醫(yī)學院免疫學教研室,貴州省免疫分子應用研究工程中心,遵義563099)
目的:在體外細胞水平,初步探討TLR2/4在MTB Hsp16.3刺激小鼠骨髓來源巨噬細胞的表達及作用。方法:從BALB/c小鼠的脛腓骨中取骨髓細胞,與GM-CSF共培養(yǎng)獲得骨髓來源的M0型巨噬細胞,流式檢測F4/80和CD11b的表達并觀察其形態(tài);100 ng/ml MTB Hsp16.3刺激M0巨噬細胞,分別培養(yǎng)0、12、24、36、48、72 h,Real-time PCR檢測不同時間點TLR2/4的表達水平;利用RNAi干擾技術沉默巨噬細胞表面的TLR2/4受體,MTB Hsp16.3刺激沉默TLR2/4的M0巨噬細胞,分別培養(yǎng)0、12、24、36、48、72 h,Real-time PCR檢測不同時間點TLR2/4及Ym-1、Fizz1、IL-10 、TNF-α、iNOS、TGF-β細胞因子的表達水平。結(jié)果:M0型巨噬細胞在MTB Hsp16.3刺激后出現(xiàn)了較短的偽足,沉默TLR2/4 的M0型巨噬細胞在MTB Hsp16.3刺激后出現(xiàn)了較長偽足;M0巨噬細胞在MTB Hsp16.3刺激后高表達TLR2/4。MTB Hsp16.3刺激沉默TLR2/4 的M0型巨噬細胞,高表達M1型巨噬細胞相關的細胞因子IL-6、TNF-α、iNOS,低表達M2型巨噬細胞相關的細胞因子IL-10、TGF-β、Ym-1、Fizz1。結(jié)論:MTB Hsp16.3 通過TLR2/4促進M0型巨噬細胞向M2型轉(zhuǎn)化,抑制M1型巨噬細胞,這可能參與了MTB躲避巨噬細胞的吞噬過程。
結(jié)核分枝桿菌;Toll樣受體;MTB Hsp16.3;替代性活化巨噬細胞
結(jié)核病(Tuberculosis,TB)是由結(jié)核分枝桿菌(Mycobacterium tuberculosis,MTB)引起的慢性傳染性疾病。MTB是典型的胞內(nèi)菌,主要寄居于巨噬細胞內(nèi)與宿主免疫系統(tǒng)相互斗爭[1]。當機體感染MTB后,巨噬細胞可發(fā)揮免疫調(diào)節(jié)作用,抑制或殺滅MTB[2]。同時MTB可以通過多種機制來逃逸巨噬細胞的殺傷,如抑制巨噬細胞的凋亡,逃避巨噬細胞的吞噬及避免反應氧和反應氮產(chǎn)物的毒性等[3]。在多種機制的研究中,MTB逃避巨噬細胞的吞噬受到了較為廣泛的關注。Lopes等[4]研究發(fā)現(xiàn),Dnak蛋白可以誘導巨噬細胞向低吞噬功能的M2型轉(zhuǎn)化,使得MTB可以逃避巨噬細胞的吞噬。本實驗室前期研究發(fā)現(xiàn),MTB Hsp16.3可以刺激巨噬細胞向M2型轉(zhuǎn)化[5],然而其機制是尚不清楚,為此探討MTB Hsp16.3刺激巨噬細胞向M2型轉(zhuǎn)化的機制將有助于后續(xù)研究MTB逃避巨噬細胞吞噬的機制。Toll樣受體(Toll-like receptor,TLR)是介導天然免疫和獲得性免疫的主要受體[6],可以通過MTB的一些成分介導細胞活化,啟動天然免疫系統(tǒng)的吞噬和殺菌的效應機制。在TLR家族中,TLR2/4是巨噬細胞識別和殺傷結(jié)核分枝桿菌的重要受體[7]。在前期研究的基礎上,本實驗擬觀察巨噬細胞在MTB Hsp16.3誘導后TLR2/4的動態(tài)變化水平,觀察MTB Hsp16.3刺激沉默TLR2/4的M0型巨噬細胞相關細胞因子的動態(tài)表達情況,從TLR2/4信號通路探討MTB Hsp16.3刺激巨噬細胞向M2型轉(zhuǎn)化的作用機制,為深入探討MTB逃避巨噬細胞的吞噬作用的機制奠定基礎。
1 材料與方法
1.1 材料 MTB Hsp16.3蛋白由本實驗室制備保存。BALB/c小鼠購自重慶醫(yī)科大學實驗動物中心;PrimeScript RT reagent Kit Perfect Real-time、SYBR Premix Ex Taq 、Lipofectamine 2000購自TaKaRa公司;Real-time PCR引物合成由Invitrogen公司完成;siRNA有廣州銳博公司設計合成。
1.2 方法
1.2.1 收集和鑒定小鼠骨髓來源的巨噬細胞 將6~8周雌性BALB/c小鼠脫臼處死,無菌取小鼠脛骨和股骨骨髓細胞,經(jīng)紅細胞裂解液裂解,10% FBS的DMEM培養(yǎng)液洗滌后,1 000 r/min,10 min離心后加入10% FBS的DMEM培養(yǎng)液調(diào)整細胞密度至1×106ml-1,培養(yǎng)第7天后去除非貼壁細胞,再用10% FBS的DMEM培養(yǎng)液培養(yǎng);第8天收集貼壁細胞即為骨髓來源巨噬細胞,并在光鏡下觀察巨噬細胞的形態(tài),收集細胞。
1.2.2 siRNA轉(zhuǎn)染 由廣州銳博公司設計合成3對siRNA序列。待骨髓來源的巨噬細胞比率達到或大于80%,可進行脂質(zhì)體轉(zhuǎn)染。每孔加入2 ml含10% FBS的DMEM培養(yǎng)液,1 μl Lipofectamine 2000 與 20 pmol siRNA轉(zhuǎn)染混合物,37℃、5%CO2條件下轉(zhuǎn)染48 h,設為轉(zhuǎn)染組。
1.2.3 MTB Hsp16.3蛋白刺激巨噬細胞 100 ng/ml MTB Hsp16.3[7]分別刺激M0巨噬細胞和轉(zhuǎn)染組的巨噬細胞,每組3個復孔,并設空白對照組。置于5%CO2的37℃恒溫培養(yǎng)箱中培養(yǎng)0、12、24、36、48、72 h,培養(yǎng)后分別收集細胞及上清。
1.2.4 細胞總RNA抽提和逆轉(zhuǎn)錄 取上述3組培養(yǎng)后的細胞,TRIzol法抽提總RNA(按試劑盒說明書進行),采用紫外分光光度儀測定所提RNA的濃度和純度。以總RNA為模板,Oligo(dt)為引物,mRNA反轉(zhuǎn)錄為cDNA進行后續(xù)檢測。
1.2.5 qRT-PCR檢測相關因子mRNA的相對表達量 以反轉(zhuǎn)錄的cDNA為模板進行qRT-PCR(按照SYBR Green qPCR Master Mix 試劑盒說明書進行),反應體系:ddH2O 8 μl,SYBR 10 μl,cDNA 1 μl,上、下游引物(見表1)各0.5 μl,總體積20 μl。反應參數(shù):95℃預變性30 s;95℃變性5 s;60℃ 30 s,45個循環(huán)。Real-time PCR實驗數(shù)據(jù)用2-ΔΔCt法(Ct為熒光達到閾值時所需PCR的循環(huán)數(shù))分析樣本TLR2、TLR4、Ym-1、Fizz1、IL-10、TGF-β、iNOS、IL-6、TNF-α mRNA的相對表達量。
表1 引物序列
Tab.1 Primer sequence

GeneUpstream(5′?3′)Downstream(5′?3′)GAPDHGAGCCAAACGGGTCATCATCTGAGGGGCCATCCACAGTCTTTLR2TGGTGTCTGGAGTCTGCTGTGCGCTCCGTACGAAGTTCTCAGTLR4CCTGACACCAGGAAGCTTGAACTTCAAGGGGTTGAAGCTCAGYm?1GCAGAAGCTCTCCAGAAGCAATCCTGATTGGCCTGTCCTTAGCCCAACTGFizz1GCTGATGGTCCCAGTGAATACCCAGTAGCAGTCATCCCAGCIL?10TACAGCCGGGAAGACAATAAAGGAGTCGGTTAGCAGTATGTGF?βGGCGGTGCTCGCTTTGTATCCCGAATGTCTGACGTATTGAiNOSCTGCAGCACTTGGATCAGGAACCTGGGAGTAGCCTGTGTGCACCTGGAAIL?6GAGGATACCACTCCCAACAGACCAAGTGCATCATCGTTGTTCATACATNF?αCAGGGGCCACCACGCTCTTCTTTGTGAGTGTGAGGGTCTGG
1.2.6 ELISA檢測轉(zhuǎn)染后巨噬細胞相關細胞因子表達水平 取上述培養(yǎng)后收集好的細胞培養(yǎng)上清,用ELISA檢測TNF-α、IL-10、TGF-β細胞因子的分泌。按照試劑盒說明加樣,置于37℃溫育60 min,加洗滌液洗板5次,加顯色劑A、B,37℃避光顯色5~15 min后加入終止液,終止反應。450nm波長檢測吸光度(OD)值并測各細胞因子濃度。
2 結(jié)果
2.1 MTB Hsp16.3刺激巨噬細胞后TLR2/4的變化
2.1.1 巨噬細胞的形態(tài) 光鏡下觀察M0型巨噬細胞呈圓形(圖1A),MTB Hsp16.3刺激后巨噬細胞出現(xiàn)了較短的偽足(圖1B),MTB Hsp16.3刺激沉默TLR2/4的巨噬細胞出現(xiàn)了較長的偽足(圖1C)。
2.1.2 MTB Hsp16.3刺激巨噬細胞后TLR2/4 mRNA表達水平 在體外培養(yǎng)條件下,100 ng/ml MTB Hsp16.3刺激M0型巨噬細胞,取貼壁細胞進行Real-time PCR,檢測不同時間點TLR2/4 mRNA的相對表達水平。所有擴增曲線均無非特異性熒光,溶解曲線呈單峰,產(chǎn)物特異性好。結(jié)果顯示,MTB Hsp16.3刺激后的巨噬細胞TLR2/4 mRNA (圖2) 的相對表達量較對照組顯著增加(P<0.05)。
2.2 MTB Hsp16.3刺激沉默TLR2/4的巨噬細胞后的變化
2.2.1 TLR2/4 siRNA的沉默效率 轉(zhuǎn)染24 h后,取各組貼壁細胞進行Real-time PCR,檢測TLR2/4 mRNA表達水平,結(jié)果顯示:與陰性對照組比較,siRNA各組的TLR2/4 mRNA表達水平均明顯降低(圖3),在后續(xù)的實驗中分別用了TLR2的siRNA-3,TLR4的siRNA-1序列進行實驗。
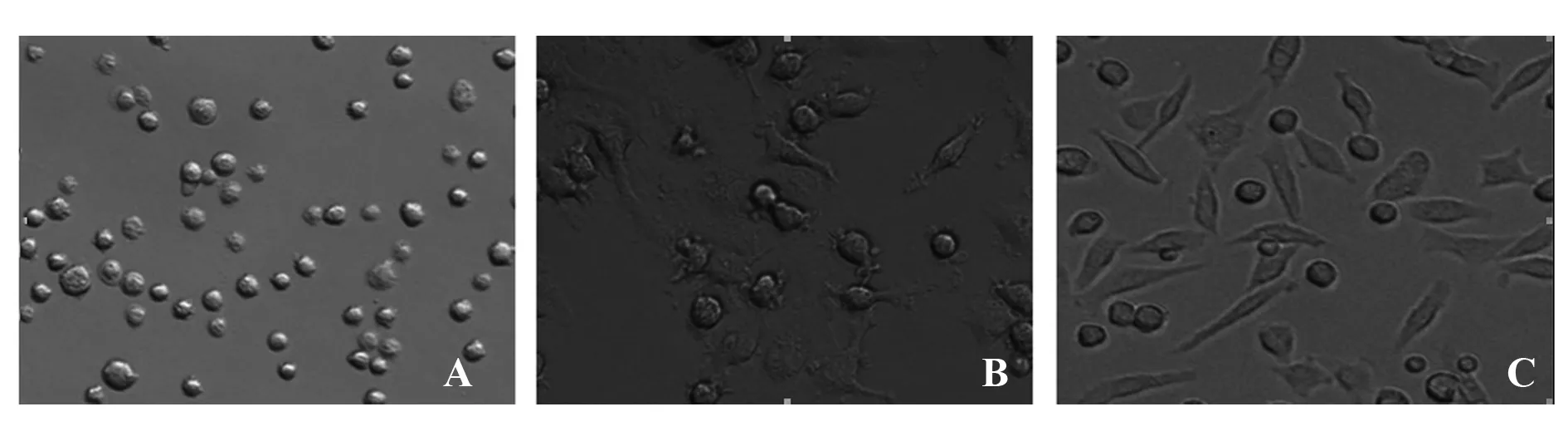
圖1 電鏡下巨噬細胞形態(tài)(×20)Fig.1 Morphology of macrophages by light microsco-pe(×20)Note: A.M0 macrophages;B.Macrophages after MTB Hsp16.3 stimulated;C.si-TLR2/4 macrophages effected with MTB Hsp16.3.
2.2.2 MTB Hsp16.3刺激沉默TLR2/4的巨噬細胞后相關細胞因子mRNA表達水平 在體外培養(yǎng)條件下,100 ng/ml MTB Hsp16.3刺激沉默TLR2/4后的巨噬細胞,分別刺激0、12、24、36、48、72 h后取貼壁細胞進行Real-time PCR,檢測不同時間點細胞因子mRNA的相對表達水平。所有擴增曲線均無非特異性熒光,溶解曲線呈單峰,產(chǎn)物特異性好。結(jié)果顯示:MTB Hsp16.3刺激沉默TLR2的巨噬細胞后Ym-1、Fizz和IL-10 mRNA(圖4A)的相對表達量較MTB Hsp16.3刺激組顯著下降(P<0.05),iNOS、IL-6和TNF-α mRNA的相對表達量較MTB Hsp16.3 刺激組顯著上調(diào)(P<0.05); MTB Hsp16.3刺激沉默TLR4的巨噬細胞后Fizz1、TGF-β和IL-10 mRNA(圖4B)的相對表達量較MTB Hsp16.3刺激組顯著下降(P<0.05),iNOS、IL-6和TNF-α mRNA的相對表達量較MTB Hsp16.3刺激組顯著上調(diào)(P<0.05)。
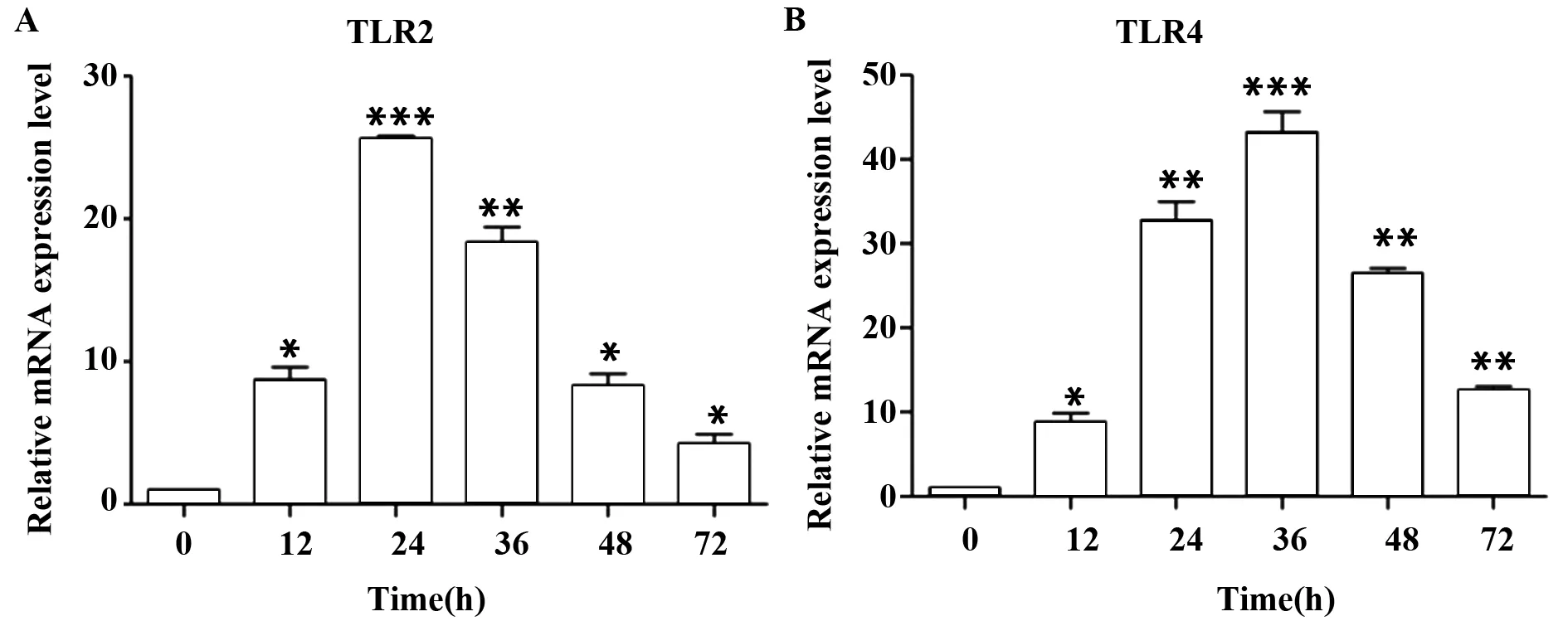
圖2 MTB Hsp16.3刺激巨噬細胞后TLR2/4 mRNA表達水平Fig.2 Expression level of TLR2/4 mRNA after MTB Hsp16.3 stimulated macrophagesNote: A.TLR2;B.TLR4;*.P<0.05;**.P<0.01;***.P<0.001.
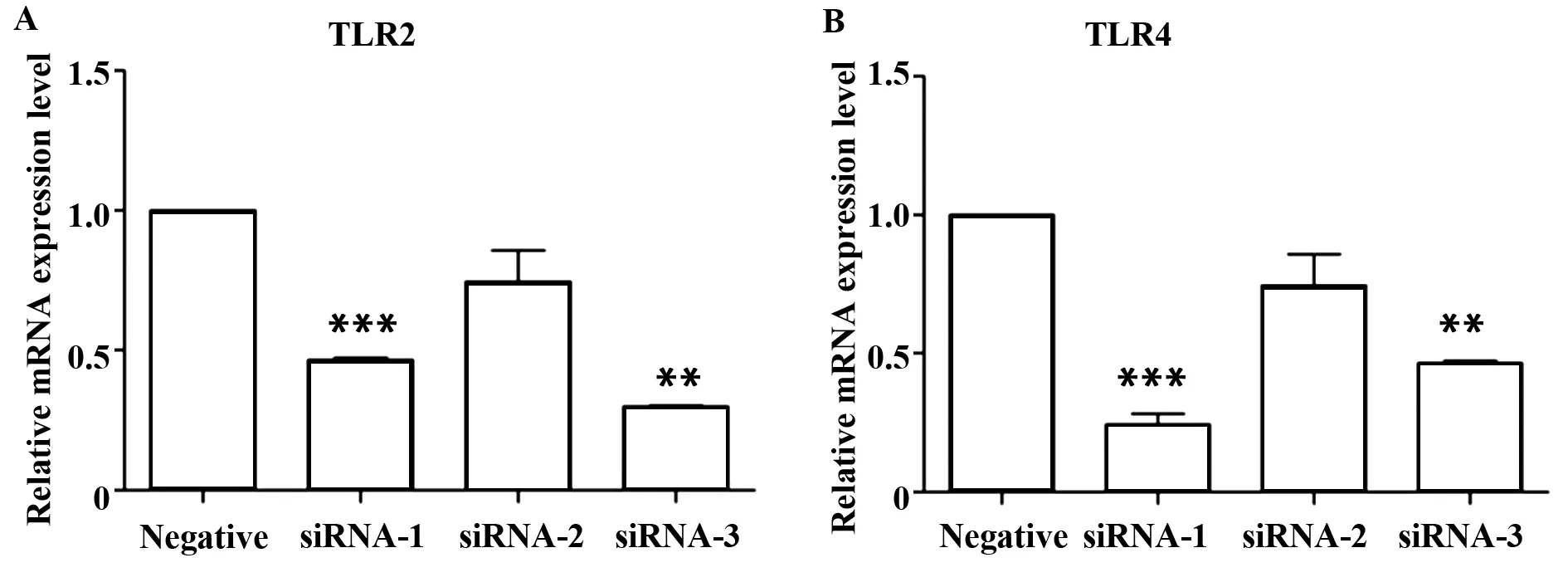
圖3 TLR2/4 siRNA的沉默效率Fig.3 Silencing efficiency of TLR2/4 siRNANote: A.TLR2;B.TLR4;**.P<0.01;***.P<0.001.
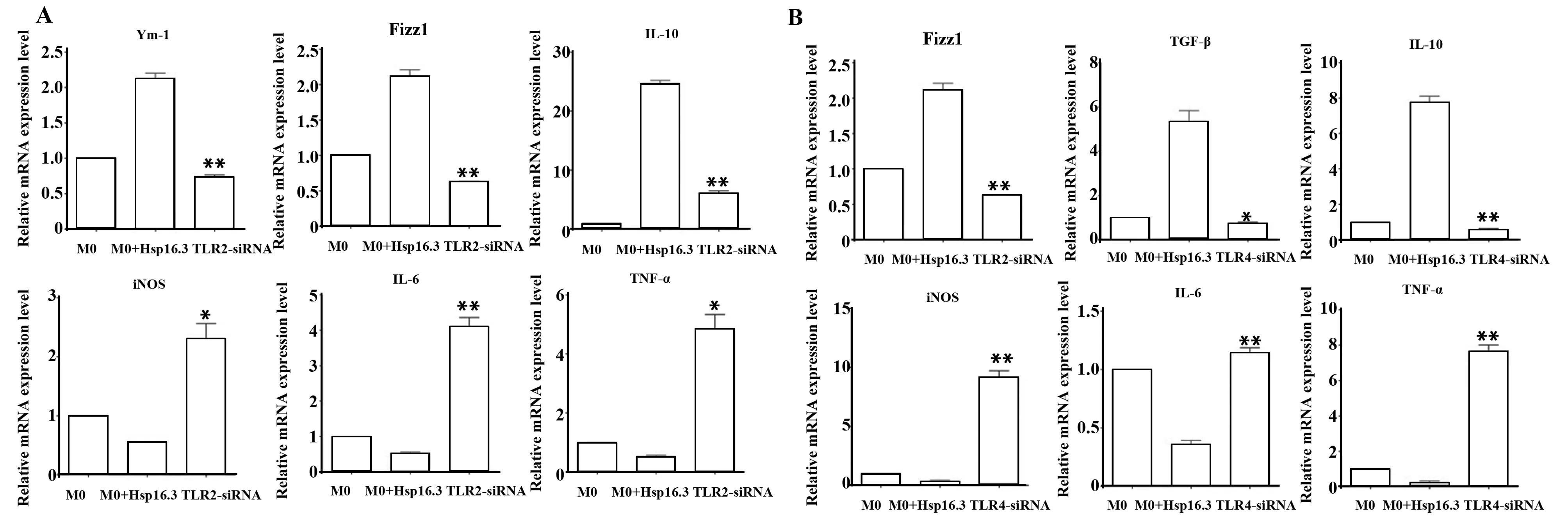
圖4 MTB Hsp16.3刺激沉默TLR2/4的巨噬細胞相關細胞因子的表達水平Fig.4 Expression levels of cytokine mRNA in TLR2/4 silencing macrophages after MTB Hsp16.3 stimulatedNote: A.TLR2;B.TLR4;*.P<0.05;**.P<0.01.
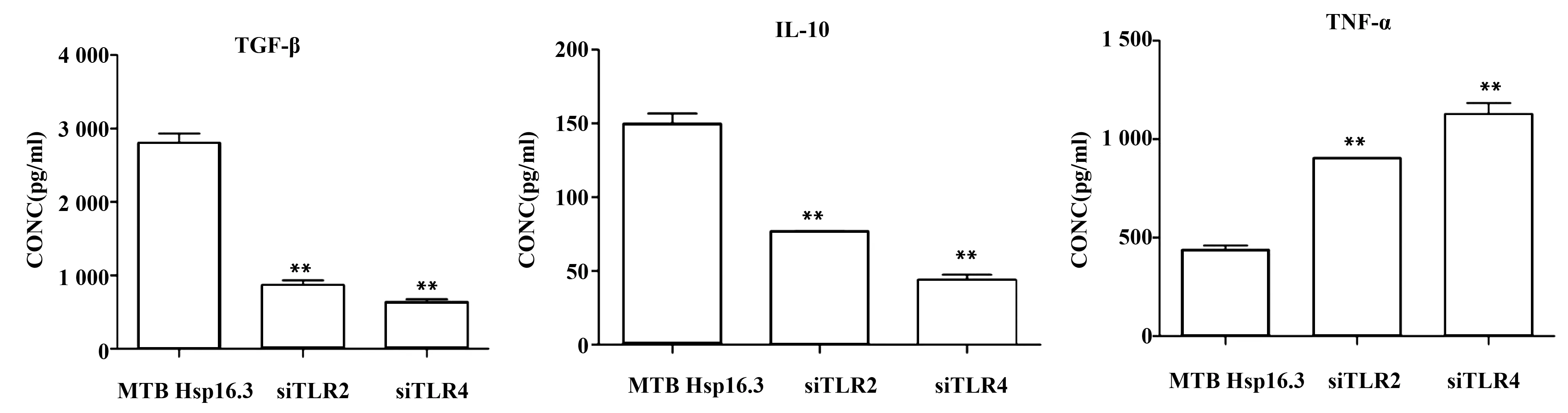
圖5 MTB Hsp16.3刺激沉默TLR2/4的巨噬細胞各細胞因子的分泌水平Fig.5 Secretion levels of cytokine mRNA in siRNA silencing macrophages after MTB Hsp16.3 stimulatedNote: **.P<0.01.
2.2.3 細胞培養(yǎng)上清進行ELISA檢測 結(jié)果顯示:MTB Hsp16.3刺激沉默TLR2/4的巨噬細胞,TGF-β和IL-10水平顯著降低(P<0.05),TNF-α水平明顯增加(P<0.05,圖5),上述各細胞因子ELISA結(jié)果與qRT-PCR結(jié)果一致。
3 討論
MTB入侵宿主后主要在巨噬細胞內(nèi)寄居,一方面巨噬細胞可以抑制或殺滅MTB,另一方面MTB可以通過逃避巨噬細胞的吞噬,抑制巨噬細胞的凋亡等機制躲避巨噬細胞的殺傷。研究發(fā)現(xiàn),MTB在逃避巨噬細胞吞噬的過程中,可以抑制吞噬小體與溶酶體融合,抑制吞噬小體的酸化以及誘導巨噬細胞轉(zhuǎn)化為低吞噬功能的M2型巨噬細胞等[8]。其中,誘導巨噬細胞轉(zhuǎn)化為低吞噬功能的M2型巨噬細胞受到了關注。本實驗室前期發(fā)現(xiàn)了MTB Hsp16.3與巨噬細胞有著一定的聯(lián)系[9]。MTB Hsp16.3是由hspX 基因編碼的,存在于MTB膜上的主要抗原蛋白,在MTB休眠階段大量表達[10]。研究發(fā)現(xiàn)MTB Hsp16.3可以抑制巨噬細胞自噬體的形成[11],抑制小鼠肺泡巨噬細胞的凋亡[12],從而使得MTB在巨噬細胞內(nèi)存活。前期實驗研究發(fā)現(xiàn),MTB Hsp16.3可以刺激巨噬細胞向低吞噬的M2型轉(zhuǎn)化,為深入探討MTB逃避巨噬細胞的吞噬作用的機制奠定基礎。
Toll樣受體是一種重要的模式識別受體,是宿主天然免疫系統(tǒng)的關鍵成分。MTB的某些成分可以介導細胞活化,啟動天然免疫系統(tǒng)的吞噬和殺菌的效應機制。研究發(fā)現(xiàn),在小鼠慢性結(jié)核病的模型中,TLR4低表達的小鼠可轉(zhuǎn)變?yōu)槁苑窝祝腋巍⑵ⅰ⒛I等器官可見MTB的擴散[13];Lacavé等[14]發(fā)現(xiàn)鞭毛蛋白可以通過TLR5誘導巨噬細胞向M2型極化。提示Toll受體與MTB、M2型巨噬細胞有著一定的關系。為此,本研究將從TLR2/4信號通路探討MTB Hsp16.3刺激巨噬細胞向M2型轉(zhuǎn)化的作用機制。
實驗結(jié)果顯示:MTB Hsp16.3刺激M0型巨噬細胞后,鏡下觀察巨噬細胞出現(xiàn)了較短的偽足,整體形態(tài)較為收攏,而沉默TLR2/4后巨噬細胞出現(xiàn)了較長的偽足;Real-time PCR檢測MTB Hsp16.3刺激M0型巨噬細胞后TLR2/4表達情況,顯示刺激后巨噬細胞高表達TLR2/4;Real-time PCR檢測沉默TLR2/4的巨噬細胞在MTB Hsp16.3刺激后,相關細胞因子的表達情況,顯示沉默TLR2后,M0型巨噬細胞高表達iNOS、IL-6和TNF-α,低表達M2相關細胞因子Ym-1、Fizz1和IL-10;沉默TLR4后,iNOS、IL-6和TNF-α表達水平顯著上調(diào)Fizz1、TGF-β和IL-10的表達水平顯著下降。ELISA結(jié)果與real time PCR結(jié)果一致。
上述研究發(fā)現(xiàn),在MTB Hsp16.3刺激沉默TLR2/4的巨噬細胞后,M2型巨噬細胞相關細胞因子呈現(xiàn)低表達狀態(tài),提示在MTB Hsp16.3刺激巨噬細胞向M2轉(zhuǎn)化的過程中,TLR2/4起到一定的作用。為此我們猜測MTB Hsp16.3可能通過與巨噬細胞膜上TLR2/4結(jié)合,激活相關的信號通路,從而使得巨噬細胞向M2型轉(zhuǎn)化,分泌相關的細胞因子。本實驗僅在體外細胞水平初步探討了TLR2/4在MTB Hsp16.3刺激小鼠巨噬細胞過程中表達及作用,然而具體的信號途徑及在體內(nèi)感染MTB模型中,MTB Hsp16.3誘導巨噬細胞向M2轉(zhuǎn)化的過程是通過MTB Hsp16.3直接釋放到胞外或者其他方式與巨噬細胞的TLR2/4結(jié)合都需要進行后續(xù)的深入探討。本研究將有助于闡明MTB逃避巨噬細胞的吞噬作用的機制,為結(jié)核病免疫防治提供合適的候選分子靶點。
[1] 彭衛(wèi)生,王英年,肖成志.新編結(jié)核病學[M].第2版.北京:中國醫(yī)藥科技出版社,2003:83.
[2] West AP,Kob lansky AA,G hosh S.Recognition and signaling by toll-like receptors[J].Annu Rev Cell Dev Biol,2006,22:409-437.
[3] Sutherland JS,Adetifa IM,Hill PC,etal.Pattern and diversity of cytokine production differentiates between Mycobacterium tuberculosis infection and disease[J].Eur J Immunol,2009,39:1-7.
[4] Lopes RL,Borges TJ,Araújo JF,etal.Extracellular mycobacterial DnaK polarizes macrophages to the M2-like phenotype[J].PLoS One,2014,9(11):e113441.
[5] 李姍姍,秦 歡,丁陳波,等.結(jié)核分枝桿菌Hsp16.3刺激小鼠巨噬細胞向M2分化[J].中國免疫學雜志,2015,32(12):1595-1606.
[6] Harding CV,Boom WH.Regulation of antigen presentation by Mycobacterium tuberculosis:a role for Toll-like receptors[J].Nat Rev Microbiol,2010,8(4):296-307.
[7] Bonfim CV,Mamoni RL,Blotta MH.TLR-2,TLR-4 and dectin-1 expression in human monocytes and neutrophils stimulated by Paracoccidioides brasiliensis[J].Med Mycol,2009,47(7):722-733.
[8] Goldberg MF,Saini NK,Porcelli SA.Evasion of innate and adaptive immunity by Mycobacterium tuberculosis[J].Microbiol Spectr,2014,2(5):1-24.
[9] 秦 歡,高紹瑩,袁建波,等.結(jié)核分枝桿菌小分子熱休克蛋白16.3蛋白的原核表達及功能檢測[J].細胞與分子免疫學雜志,2014,30(5):480-485.
[10] Cunningham AF,Spreadbury CL.Mycobacterialstationary phase induced by low oxygen tension:cell wall thickening and localization of the 16-kilod alton alpha-crystallin homolog[J].J Bacteriol,1998,180(4):801-808.
[11] 師長宏,江 鷹,張 海,等.結(jié)核分枝桿菌Hsp16.3蛋白影響小鼠巨噬細胞自噬形成的實驗研究[J].細胞與分子免疫學雜志,2011,27(12):1301-1303.
[12] 姚 楠,董江濤,張萬江,等.結(jié)核分枝桿菌Hsp16.3與感染巨噬細胞凋亡的相關性研究[J].中國病原生物學雜志,2012,7(8):569-573.
[13] Abel B,Thieblemont N,Quesniaux VJ,etal.Toll-Like receptor 4 expression is required to control chronic mycobacterium tuberculosis infection in mice[J].J Immunol,2002,169(6):3155-3162.
[14] Lacavé-Lapalun JV,Benderitter M,Linard C.Flagellin or lipopolysaccharide treatment modified macrophage populations after colorectal radiation of rats[J].J Pharmacol Exp Ther,2013,346(1):75-85.
[收稿2016-06-06 修回2016-08-22]
(編輯 倪 鵬)
Study of expression and regulation of TLR2/4 in mycobacterium tuberculosis heat shock proteins 16.3 effect on mouse bone marrow-derived macrophages
LIShan-Shan,QINHuan,LIUQian-Yi,XULin,ZHANGJi-Dong,F(xiàn)ENGJi-Hong,LILong-Mei,PANHong-Fei,LUOJun-Min.
DepartmentofImmunology,ZunyiMedicalCollege,ImmuneMoleculesApplicationResearchCenterinGuizhouProvince,Zunyi563099,China
Objective:To study the expression and regulation of TLR2/4 in mycobacterium tuberculosis heat shock proteins 16.3 (mycobacterium tuberculosis heat shock proteins 16.3,MTB Hsp16.3) effect on mouse bone marrow-derived macrophages in vitro.Methods: Bone marrow cells were isolated from tibia and femurs of BALB/c mice and incubated with GM-CSF,then detected the expression of CD11b and F4/80 with flow cytometry and observed morphology.The M0 macrophages were stimulated with MTB Hsp16.3 for 0 h,12 h,24 h,36 h,48 h and 72 h.Real-time PCR detected the expression of TLR2/4 in intracellular at different time point.Silencing macrophages cell surface TLR2/4 molecules by siRNA technology which stimulated with MTB Hsp16.3 for 0 h,12 h,24 h,36 h,48 h and 72 h.Real-time PCR detected the expression of TLR2/4,Ym-1,Fizz1,IL-10,TNF-α,iNOS and TGF-β in intracellular at different time point.Results: Morphology analysis showed that MTB Hsp16.3 stimulated macrophages were round cells stretching out pseudopodia,whereas MTB Hsp16.3 stimulated silencing TLR2/4 macrophages had elongated fibroblastoid.Real time PCR detected the expression of TLR2/4 were upregulated after MTB Hsp16.3 stimulated M0 macrophages.MTB Hsp16.3 stimulated silencing TLR2/4 macrophages the expression of IL-6,TNF-α,iNOS were upregulated,whereas IL-10,TGF-β,Ym-1 and Fizz1 were downregulated.Conclusion: MTB Hsp16.3 may stimulated M0 macrophages to M2 macrophages and suppress M1 macrophages through binding with TLR2/4 receptor,which may be involved the progresss of MTB evaded macrophage phagocytosis.
Mycobacterium tuberculosis;Toll like receptors;Mycobacterium tuberculosis heat shock proteins 16.3;Alternatively activated macrophage
10.3969/j.issn.1000-484X.2017.01.007
①本文受國家自然科學基金地區(qū)項目(81460249)資助。
李姍姍(1990年-),女,在讀碩士,主要從事感染免疫學方面的研究,E-mail:lss2958@126.com。
及指導教師:羅軍敏(1970年-),女,碩士,教授,碩士生導師,主要從事感染免疫學方面的研究,E-mail: luojm128@163.com。
R392.11
A
1000-484X(2017)01-0036-05
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
學苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
人大建設(2019年12期)2019-05-21 02:55:32
文苑(2018年21期)2018-11-09 01:23:06
中國衛(wèi)生(2015年9期)2015-11-10 03:11:12
中國衛(wèi)生(2014年3期)2014-11-12 13:18:12
云南中醫(yī)學院學報(2014年3期)2014-07-31 18:57:34
中國火炬(2014年4期)2014-07-24 14:22:19
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00