農桿菌介導bar基因轉化水稻胚性愈傷組織的研究
2017-02-22 06:50:59何少海姚盛存豐宇凱李飛飛
浙江農林大學學報 2017年1期
關鍵詞:水稻
何少海,姚盛存,豐宇凱,汪 軻,李飛飛
(浙江農林大學 農業與食品科學學院 浙江省農產品品質改良技術研究重點實驗室,浙江 臨安 311300)
農桿菌介導bar基因轉化水稻胚性愈傷組織的研究
何少海,姚盛存,豐宇凱,汪 軻,李飛飛
(浙江農林大學 農業與食品科學學院 浙江省農產品品質改良技術研究重點實驗室,浙江 臨安 311300)
為建立以草丁膦為選擇標記的農桿菌Agrobacterium tumefaciens介導的水稻Oryza sativa胚性愈傷組織轉化體系,通過農桿菌介導法將 bar基因導入了 ‘日本晴’Oryza sativa ‘Nipponbare’ ‘秀水134’ ‘Xiushui 134’和‘中花11’ ‘Zhonghua 11’等3個水稻品種的胚性愈傷組織。在分別以選擇壓15.0,10.0和10.0 mg·L-1的草丁膦質量濃度進行了3次選擇后,聚合酶鏈式反應(polymerase chain reaction,PCR)發現 ‘日本晴’ ‘秀水134’和 ‘中花11’的愈傷組織轉化效率分別是77.9%,67.5%和92.2%,獲得的抗性愈傷組織可以進一步分化成苗。它們的分化率依次是76.1%,44.4%和86.7%;成苗率分別為65.6%,66.7%和30.7%。在以草丁膦作為篩選標記時,首選‘日本晴’作為轉化受體進行遺傳轉化。該體系為以抗除草劑基因作為篩選標記的水稻遺傳轉化和基因功能驗證提供了技術基礎,獲得的抗草丁膦轉化植株為水稻育種提供了材料。圖5表3參32
分子生物學;草丁膦;bar基因;胚性愈傷組織;粳稻;農桿菌介導法轉化
草丁膦(glufosinate,PPT)又叫草銨膦,是一種低毒且廣譜性的有機磷類除草劑。它被植物吸收后,能有效抑制谷酰胺合成酶的活性,使銨在細胞里迅速積累,最后導致整個植物體死亡[1-2]。其抗性bar基因(bialaphos resistance gene)來自土壤吸水鏈霉菌Streptomyces hygroscopicus,它編碼膦絲菌素乙酰轉移酶(phosphinthricin acetyltransferase,PAT),PAT能使PPT的氨基發生乙酰化,進而解除PPT對植物的毒性[3]。因此,含有bar基因的植物對草丁膦除草劑具有一定的抗性[4]。將bar基因轉入農作物,不僅有效去除雜草,而且對施用作物無毒害。轉bar基因農作物的培育和應用受到育種者的廣泛重視[5-7],轉抗草丁膦的油菜Brassica campestris和玉米Zea mays已經商業化生產[8]。水稻Oryza sativa是世界性的糧食作物,中國是世界上最大的水稻生產和消費國。隨著雜交育種和分子輔助選擇育種技術的應用和發展,水稻產量不斷提高,品質也得到了極大改良[9]。然而,目前仍有一些因素制約水稻生產。例如病蟲害和田間雜草等生物逆境以及一些旱災等非生物逆境都會造成水稻嚴重的減產。特別是雜草不僅與水稻爭奪水分及養料,而且提高人工成本,已經成為水稻生產上的一大難題[10]。bar基因不僅可以作為選擇標記基因,而且能使水稻具有抗除草劑的優良特征。轉bar基因水稻能有效除去雜草,大大減少勞動力[11]。1996年美國對抗除草劑Liberty(草丁膦的商業名)轉基因水稻進行抗性鑒定試驗,噴施Liberty后,雜草被有效控制,而轉基因水稻不受影響。與此同時,國內外開展了抗草丁膦基因轉化水稻的相關研究。于志華等[12]采用基因槍法獲得了抗bar基因旱稻植株,試圖解決旱稻田間直播的雜草問題。CAO等[13]將bar基因作為篩選標記,利用微粒轟擊法轉化水稻的懸浮細胞,獲得轉化植株。PARK等[14]選用水稻莖尖分生組織作為轉化受體,通過農桿菌介導法,獲得幾棵轉基因植株,在后代中可以穩定遺傳。黃大年等[15]用基因槍分別對水稻的幼胚進行轉化,選取20.0~40.0 mg·L-1的草丁膦進行選擇,獲得6株轉基因水稻,轉化效率很低。簡玉瑜等[16]通過基因槍將蠶抗菌肽基因轉化發芽種胚、未成熟胚和愈傷組織3種組織,選用4.0 mg·L-1的Basta進行抗性篩選,只有發芽種胚獲得5株抗性植株,分子檢測3株為陽性株。許新萍等[17]用基因槍對水稻的愈傷組織進行轉化,選取2.0~4.0 mg·L-1的Basta進行選擇,并獲得轉基因植株,轉化效率為89.6%。CHEN等[18]用農桿菌介導法對 ‘明恢63’的愈傷組織進行轉化,先用25.0 mg·L-1的Basta篩選8~9周后,再用20.0 mg·L-1的Basta選1次,獲得轉基因植株的陽性率為70%。綜上所述,所選的受體部位是懸浮細胞、莖尖分生組織、幼胚、成熟胚和愈傷組織等,草丁膦的篩選質量濃度為0.5~40.0 mg·L-1,沒有一個確定的轉化濃度。目前,以潮霉素作為選擇壓的水稻轉基因體系已經非常成熟。但利用草丁膦作為篩選劑轉化水稻胚性愈傷組織的報道較少。基于此,本研究選取具有成功再生體系的 ‘日本晴’ ‘秀水134’ ‘中花11’的胚性愈傷組織作為受體,篩選出最適的草丁膦轉化質量濃度,再利用農桿菌介導法,將bar基因轉化這些水稻的胚性愈傷組織,獲得抗除草劑轉基因水稻,建立以草丁膦為選擇壓的高效轉化體系,為后續選育生產中農藝性狀良好的抗性品種提供轉基因材料。
1 材料和方法
1.1 材料
粳稻 ‘日本晴’Oryza sativa‘Nipponbare’ ‘中花11’Oryza sativa‘Zhonghua 11’和 ‘秀水134’Oryza sativa‘Xiushui 134’種子;具有草丁膦抗性的pKF111雙元載體;純度為99%草丁膦;EHA105農桿菌菌株。
1.2 方法
1.2.1 胚性愈傷組織的誘導 ‘日本晴’ ‘秀水134’和 ‘中花11’成熟種子用質量濃度為30%次氯酸鈉表面消毒,無菌水洗滌2~3次,接種在胚性愈傷組織誘導的培養基R1[NB(N6培養基+B5維生素)+ 2.5 mg·L-12.4-D+0.5 g·L-1脯氨酸+0.3 g·L-1水解酪蛋白+30 g·L-1蔗糖+4.0 g·L-1非特膠(phytalgel)上,15 d后種子表面出現少量的愈傷組織,挑取出來繼代在相同的培養基(2.4-D減為2 mg·L-1)上,胚性愈傷組織組織進一步增殖,待用做轉化[19-20]。
1.2.2 3種胚性愈傷組織的草丁膦敏感性試驗和水稻遺傳轉化 將誘導獲得的3種基因型胚性愈傷組織分別接種在含有0,5,10,15,20和50 mg·L-1草丁膦的胚性愈傷組織誘導培養基上,接種25塊·培養皿-1大小均一的新鮮愈傷組織,3個重復,26℃培養25 d后,觀察胚性愈傷組織的死亡數,并統計死亡率,確定合適的草丁膦篩選濃度。將含有bar基因載體的農桿菌菌液培養過夜,用菌液稀釋培養基R2(NB+500 mg·L-1脯氨酸+300 mg·L-1水解酪蛋白+30 g·L-1蔗糖+20 mg·L-1乙酰丁香酮)稀釋光密度值至0.6~0.8,侵染水稻的胚性愈傷組織15 min,期間不停晃動,之后用濾紙吸干多余的菌液,放在鋪有濾紙的共培養培養基R3(NB+500 mg·L-1脯氨酸+300 mg·L-1水解酪蛋白+30 g·L-1蔗糖+4.0 g·L-1phytalgel+20 mg·L-1乙酰丁香酮)上暗培養2 d,培養后的胚性愈傷組織用含有500 mg·L-1頭孢霉素的無菌水洗2次,用濾紙吸干水分,轉接抗性選擇培養基R4(NB+2 mg·L-12.4-D+500 mg·L-1脯氨酸+300 mg·L-1水解酪蛋白+500 mg·L-1谷氨酰胺+30 g·L-1蔗糖+4.0 g·L-1phytalgel+500 mg·L-1頭孢霉素+篩選出的合適草丁膦質量濃度)上篩選3次。每15 d繼代1次。3輪選擇后挑取抗性愈傷組織系,進行編號,并轉接在分化培養基R5(NB+0.5 mg·L-1NAA+3 mg·L-16-BA+500 mg·L-1脯氨酸+300 mg·L-1水解酪蛋白+500 mg·L-1谷氨酰胺+30 g·L-1蔗糖+4.0 g·L-1phytalgel)上進行分化培養,同時提取抗性愈傷組織系的DNA,進行聚合酶鏈式反應(PCR)檢測,統計轉化率。胚性愈傷組織出現的綠點進一步分化出不定芽,待不定芽長到2~3 cm,轉接在生根培養基R6(1/2 NB+20 g·L-1蔗糖+8.0 g·L-1瓊脂)上,待苗長到10 cm,并有大量的根,移栽營養缽成活后,再移至大田成活,收種。
1.2.3 抗性愈傷組織系和葉片的DNA提取和PCR分子檢測 利用十六烷基三甲基溴化銨(CTAB)法提取抗性愈傷組織系和再生植株葉片的DNA,用bar基因的引物(F:ACCATCGTCAACCACTACATCG,R:GCTGCCAGAAACCCACGTCAT)進行PCR檢測,擴增的目的片段是430 bp。
1.2.4 T0代植株的田間草丁膦篩選 對照和轉基因的植株的展開葉片用棉球蘸取質量濃度分別為50,100和300 mg·L-1的草丁膦,5 d后觀察葉片的變化。
2 結果和分析
2.1 3個水稻品種的愈傷組織對草丁膦抗性篩選
將 ‘日本晴’ ‘秀水134’和 ‘中花11’成熟胚誘導的米黃色且有光澤的胚性愈傷組織接種在分別含有6個質量濃度的草丁膦的選擇培養基R4上,死亡率為50%~60%的愈傷組織對應的草丁膦質量濃度即可用作選擇的質量濃度。結果如表1和圖1所示:3個水稻品種的胚性愈傷組織在空白對照上全部存活,且大量增殖;在5.0 mg·L-1草丁膦的選擇壓下,死亡率為19%~23%,發生褐變的愈傷組織數量較少,且大部分愈傷組織有明顯增殖,沒有起到篩選作用;隨著草丁膦質量濃度的增加,在10.0 mg·L-1時, ‘秀水134’和 ‘中花11’2個品種死亡率為51%~56%,且與5.0 mg·L-1相比存在極顯著性差異,達到選擇效果; ‘日本晴’在15.0 mg·L-1下,死亡率54.53%,且與10.0 mg·L-1相比存在極顯著性差異,達到選擇效果;但 ‘日本晴’在20 mg·L-1下,死亡率60%以上,選擇效果偏大; ‘秀水134’和‘中花11’在15.0 mg·L-1下,死亡率為55%~60%,選擇效果偏大;而質量濃度50.0 mg·L-1時,3個水稻品種死亡率達到90%以上,表明該質量濃度已經超過臨界質量濃度,選擇壓力太大。綜上所述, ‘日本晴’的草丁膦選擇質量濃度應為15.0 mg·L-1, ‘秀水134’和 ‘中花11’的草丁膦選擇質量濃度都為10.0 mg·L-1。
2.2 抗草丁膦bar基因轉化3個水稻品種和植株再生
用粳稻誘導出的米黃色且有光澤的胚性愈傷組織(圖2A)作為受體,利用農桿菌介導法轉化bar基因。將侵染后 ‘日本晴’ ‘秀水134’和 ‘中花11’的胚性愈傷組織分別在質量濃度為15,10和10mg·L-1的草丁膦下進行3次選擇培養。第1次選擇后胚性愈傷組織大部分出現褐變(圖2B);第2次選擇后,部分愈傷組織不抗繼續褐化和變黑,部分周邊長出新的抗性愈傷組織(圖2C);挑取新生的愈傷組織系進行第3輪選擇,出現增殖(圖2D),抗性愈傷組織系分化出綠點(圖2E),進一步長出幼葉(圖2F),待葉片伸長到2~3 cm時,再生苗快速生根,獲得完整的再生植株(圖2G),移栽成活(圖2H)。從胚性愈傷組織進行農桿菌介導法的轉化到獲得抗性胚性愈傷組織約2~3個月,抗性愈傷組織分化成苗約2個月。整個轉化過程需要4~6個月。
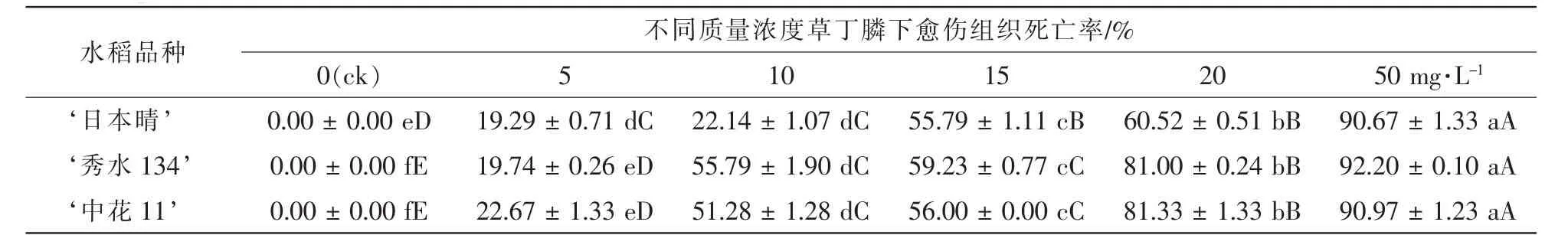
表1 3個水稻品種胚性愈傷組織在不同質量濃度草丁膦的培養基上培養25 d后死亡率統計Table 1 Mortality statistics of three kinds of rice EC(embryogenic callus)cultured on mediums containing 0,5,10,15,20,50 mg·L-1glufosinate after 25 d

圖1 3個水稻品種的胚性愈傷組織對草丁膦的敏感性試驗Figure 1 Sensitivity test of three cultivars of rice EC to glufosinate

圖2 水稻胚性愈傷組織為受體轉化到再生圖Figure 2 Figures of rice embryogenic callus from transformation to regeneration
2.3 轉化率、分化率和成苗率的比較
抗性選擇后, ‘日本晴’ ‘秀水134’和 ‘中花11’分別獲得86,40和90塊抗性愈傷組織系,提取這些抗性愈傷組織系的DNA,進行PCR檢測,分別有67,27和83塊愈傷組織系均擴增出目的條帶,轉化率分別為77.9%,67.5%和92.2%(表2)。圖3A是 ‘日本晴’隨機選擇的10塊抗性愈傷組織的檢測圖,除了泳道8和13,均擴增出和陽性對照(泳道3)一樣的條帶,轉化率均達到67%以上。說明篩選的草丁膦質量濃度是適合轉化的質量濃度,同時淘汰非抗性愈傷組織系,減少后續的繼代工作量。
將這些陽性愈傷組織系進行分化培養發現:3個水稻品種的分化能力是不同的。 ‘秀水134’分化綠點最少,分化能力最弱(圖4A),共12塊愈傷組織分化出綠點,分化率為44.4%; ‘中花11’(圖4B)有72塊愈傷組織系分化出綠點,分化率為86.7%; ‘日本晴’分化綠點最多,分化能力最強(圖4C),51塊愈傷組織系分化出綠點,分化率為76.1%(表2)。
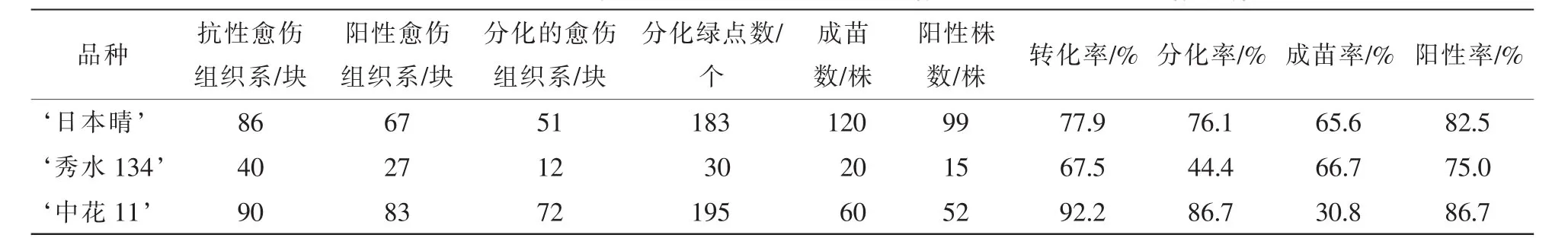
表2 3個水稻品種的轉化率、分化率和出苗率比較Table 2 Transformation rate,differentiation rate and emergence rate in three rice genotypes
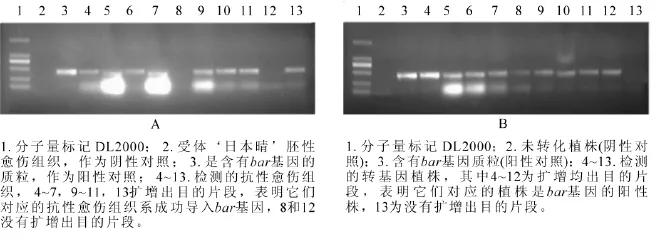
圖3 抗性愈傷組織系(A)和轉基因植株葉片(B)的PCR檢測圖Figure 3 Detection of bar gene in glyphosate-resistant EC lines by PCR analysis
2.4 轉基因植株分子檢測結果
對獲得的轉基因植株葉片提DNA進行PCR擴增,以bar基因引物引物進行PCR擴增。 ‘日本晴’‘秀水134’和 ‘中花11’分別獲得120株,20株和60株轉基因小植株,其中bar基因陽性株有99株、15株和52株,陽性率為82.5%,75%和86.7%(表2)。圖3B為部分轉基因植株的檢測結果。
2.5 轉基因植株的草丁膦抗性鑒定結果
對移栽成活的轉基因植株進行草丁膦抗性檢測,將蘸有不同質量濃度草丁膦的棉球粘附在轉基因水稻的葉片上,5 d后,轉基因和非轉基因水稻出現了明顯不同的反應。圖5是 ‘日本晴’葉片在低質量濃度為50.0 mg·L-1草丁膦處理時,未轉化植株葉片上出現枯黃的斑點,在100.0 mg·L-1草丁膦處理后,出現較大面積枯黃的斑,在高質量濃度300.0 mg·L-1草丁膦處理后大面積變枯且嚴重灼傷。 但轉基因水稻葉片在50.0,100.0和300.0 mg·L-1草丁膦處理后仍無變化,表明轉基因水稻抗性提高了約6倍。
‘日本晴’ ‘秀水134’和 ‘中花11’分別獲得的99株、15株和52株PCR陽性株,其中分別有10株、5株和12株的葉片變黃枯,其余均表現綠色,草丁膦陽性率分別為89.9%,66.7%和76.9%(表3),轉基因水稻表現出草丁膦抗性。

表3 bar轉基因植株抗性鑒定分析Table 3 Glufosinate-resistant analysis of transgenic plants
3 討論
3.1 水稻遺傳轉化中草丁膦篩選質量濃度的確定和轉化效率
轉基因水稻的獲得依賴于篩選體系的選擇效果。 bar基因常作為選擇標記基因,具有草丁膦抗性,除了選擇效率高外,還可以帶來抗除草劑的農藝性狀。將bar基因或與其他目的基因串聯在一起轉化水稻,獲得抗草丁膦水稻。確定一個合適的草丁膦篩選質量濃度,是提高轉化效率和優化轉化體系的關鍵步驟。本實驗研究發現不同基因型水稻的胚性愈傷組織對草丁膦具有明顯不同的抗性。為了確定以胚性愈傷組織為受體最佳的草丁膦選擇質量濃度,本研究選取了5個 5.0,10.0,15.0,20.0和50.0 mg·L-1草丁膦質量濃度梯度,對 ‘日本晴’ ‘秀水134’和 ‘中花11’的胚性愈傷組織進行草丁膦敏感試驗,并確定有效的抗性篩選草丁膦質量濃度分別為15.0,10.0和10.0 mg·L-1(圖1),轉化效率分別為77.9%,67.5%和92.2%,達到了一個很好的選擇效果。本研究為以不同的水稻胚性愈傷組織為轉化受體,提供可靠的草丁膦選擇質量濃度。選取合適質量濃度的草丁膦進行選擇,是成功轉化的關鍵之一。

圖5 轉基因植株葉片草丁膦抗性檢測Figure 5 Glufosinate-resistant analysis of the leaves for transgenic plants
3.2 3個水稻品種分化率和再生效率的比較
‘日本晴’ ‘秀水134’和 ‘中花11’的3個水稻品種均能獲得成功的再生和轉化體系,是目前水稻轉基因常用的受體材料。本研究發現:當以草丁膦為選擇標記時,它們的分化效率是不同的。其中‘中花11’的分化能力最強,分化率最高,高達86.7%; ‘秀水134’的分化能力最差,分化率只有44.4%; ‘日本晴’介于兩者之間,分化率為76.1%。同時也對它們的再生效率進行了統計,研究發現,‘日本晴’有51個愈傷組織系分化,分化出183綠點,成苗數是120株,成苗率是65.6%; ‘秀水134’有12個愈傷組織系分化,分化出30綠點,成苗數20株,成苗率是66.7%; ‘中花11’有72個愈傷組織系分化,分化出195綠點,成苗數60株,成苗率是30.7%。 ‘中花11’分化率最高,成苗率最低;‘日本晴’的分化率居中, ‘秀水134’的分化率最低; ‘日本晴’和 ‘秀水134’的成苗率基本一樣高。據此,當以草丁膦作為篩選標記時,首選 ‘日本晴’作為轉化受體進行遺傳轉化。
3.3 以胚性愈傷組織作為轉化受體的轉化植株的變異率問題的解決
水稻轉基因除了常用的基因槍法[21-23],農桿菌介導法也是目前一種可行有效的方法。用農桿菌轉化水稻的胚性愈傷組織,可以減少嵌合體,提高轉化效率[24-30],但是仍然出現一些再生植株的變異問題。例如多分蘗,無花粉等[31]。采取以下措施:①縮短抗性選擇時間,加強分化選擇力度。經過1次抗性選擇,進行分化培養,分化培養基上加入減半的草丁膦質量濃度,直到分化出綠點為止。②選用最新誘導出的、有光澤、淡黃色的胚性愈傷組織,剛從愈傷組織誘導出的胚性愈傷組織,及時挑出,增殖一定數量,用于轉化[32];③減少繼代次數。從轉化到獲得再生苗約4~5個月,期間繼代5~6次;統計再生植株的變異率低于20%。本研究通過對水稻草丁膦敏感實驗,成功建立了以草丁膦為篩選標記的農桿菌介導轉化水稻胚性愈傷組織的體系。整個轉化周期 4~6個月。獲得抗300 mg·L-1草丁膦的 T0轉基因植株。該體系為以除草劑作為選擇壓的水稻遺傳轉化提供了技術支持,獲得的抗草丁膦水稻為水稻育種提供了材料。
[1] THOMPSON C J,MOVVA N R,TIZARD R,et al.Characterization of the herbicide-resistance gene bar from streptomyces hygroscopicus[J].EMBO J,1987,6(9):2519-2523.
[2] AHRENS W H.Herbicide Handbook[M].7th ed.Illinois:Weed Science Society of America,1994:147-149.
[3] 段發平,梁成鄴,黎垣慶.Bar基因和轉Bar基因作物對研究進展[J].廣西作物,2001,21(2):166-172.
DUAN Faping,LIANG Chengye,LI Yuanqing.Research advances of bar gene and its transgenic crops[J].Gucihaia, 21(2):166-172.
[4] RATHORE K S,CHOWDHURY V K,HODGES T K.Use of bar as a selectable marker gene and for the production of herbicide-resistant rice plants from protoplasts[J].Plant Molec Biol,1993,21(5):871-884.
[5] VENGADESAN G,AMUTHA S,MURUGANATHAM M,et al.Transgenic Acacia sinuate from Agrobacterium tumefaciens-mediated transformation of hypocotyls[J].Plant Cell Rep,2006,25(11):1174-1180.
[6] YI G,SHIN Y M,CHOE G,et al.Production of herbicide-resistant sweet potato plants transformed with the bar gene[J].Biotechnol Lett,2007,29(4):669-675.
[7] LIU Yu,YANG Shixin,CHENG Yan,et al.Production of herbicide-resistant medicinal plant Salvia miltiorrhiza transformed with the bar gene[J].Appl Biochem Biotechnol,2015,177(7):1456-1465.
[8] DUKE S.Taking stock of herbicide-resistant crops ten years after introduction[J].Pest Manage Sci,2005,61(3):211-218.
[9] 楊瑞芳,白建江,方軍,等.分子標記輔助選擇選育高抗性淀粉水稻新品種[J].核農學報,2015,29(12):2259-2267.
YANG Ruifang,BAI Jianjiang,FANG Jun,et al.Establishment of marker-assisted selection system for breeding rice varieties with high resistant starch content[J].J Nucl Agric Sci,2015,29(12):2259-2267.
[10] 黎垣慶,劉剛,嚴文貴,等.轉bar基因水稻除草劑抗性遺傳研究及其應用[J].雜交水稻,2000,15(1):40-43.
LI Yuanqing,LIU Gang,YAN Wengui,et al.Inheritance of the herbicide resistance in bar-transgenic rice and its application[J].Hybrid Rice,2000,15(1):40-43.
[11] 吳明國,黃大年,林建榮,等.抗除草劑轉基因水稻穩定系TR4的獲得及其遺傳研究[J].中國水稻科學,1999,13(3): 173-175.
WU Mingguo,HUANG Danian,LIN Jianrong,et al.Establishment and genetic analysis of a stable herbicide resistant transgenic rice line TR4[J].Chin J Rice Sci,1999,13(3):173-175.
[12] 于志華,陳敏勇,顧紅雅,等.抗除草劑旱稻轉基因植株的獲得[J].北京大學學報(自然科學版),1996,32(4):499-504.
YU Zhihua,CHEN Minyong,GU Hongya,et al.Upland rice transformation[J].Acta Sci Nat Univ Pekin,1996,32(4):499-504.
[13] CAO Jun,DUAN Xiaolan,MCELROY D,et al.Regeneration of herbicide resistant transgenic rice plants following microprojectile-mediated transformation of suspension culture cells[J].Plant Cell Rep,1992,11(11):586-591.
[14] PARK S H,PINSON S R M,SMITH R H.T-DNA integration into genomic DNA of rice following Agrobcterium inoculation of isolated shoot apices[J].Plant Mol Biol,1996,32(6):1135-1148.
[15] 黃大年,朱冰,楊煒,等.抗菌肽B基因導入水稻及轉基因植株的鑒定[J].中國科學(生命科學),1997,27(1):55-62.
HUANG Danian,ZHU Bing,YANG Wei,et al.Introduction of cecropin B gene into rice (Oryza sativa L.)by particle bombardment and analysis of transgenic plants[J].Sci China Ser Life Sci,1997,27(1):55-62.
[16] 簡玉瑜,吳新榮,莫豪葵,等.應用基因槍將蠶抗菌肽基因導入水稻獲抗白葉枯病株系[J].華南農業大學學報, 1997,18(4):1-7.
JIAN Yuyu,WU Xinrong,MO Haokui,et al.Transfer of cecropin B gene into rice (Oryza sativa L.)by the biolistic method and transgenic progeny line for resistance to bacterial blight[J].J South China Agric Univ,1997,18(4): 1-7.
[17] 許新萍,衛劍文,范云六,等.基因槍法轉化秈稻胚性愈傷組織獲得可育的轉基因植株[J].遺傳學報,1999,26(3):219-227.
XU Xinping,WEI Jianwen,FAN Yunliu,et al.Fertile transgenic indica rice from micoprojectile bombardment of embryogenic callus[J].Acta Genet Sin,1999,26(3):219-227.
[18] CHEN Hao,TANG Wei,XU Caiguo,et al.Transgenic indica rice plants harboring a synthetic cry2A*gene of Bacillus thuringiensis exhibit enhanced resistance against lepidopteran rice pests[J].Theor Appl Genet,2005,111(7): 1330-1337.
[19] SAHOO R K,TUTEJA N.Development of Agrobacterium-mediated transformation technology for mature seed-derived callus tissues of indica rice cultivar IR64[J].GM Crops Food,2012,3(2):123-128.
[20] SAIKA H,TOKI S.Mature seed-derived callus of the model indica rice variety Kasalath is highly competent in A-grobacterium-mediated transformation[J].Plant Cell Rep,2010,29(12):1351-1364.
[21] BREITLER J C,LABEYRIE A,MEYNARD D,et al.Efficient microprojetile bombardment-mediated transformation of rice using gene cassettes[J].Theor Appl Genet,2002,104(4):709-719.
[22] VAIN P,JAMES V,WORLAND B,et al.Transgene behavior across two generations in a large random population of transgenic rice plants produced by particle bombardment[J].Theor Appl Genet,2002,105(6):878-889.
[23] ZHAO Yan,QIAN Qian,WANG Huazhong,et al.Hereditary behavior of bar gene cassette is complex in rice mediated by particle bombardment[J].J Genet Genom,2007,34(9):824-835.
[24] RAINERI D M,BOTTINO P,GORDON M P,et al.Agrobacterium-mediated transformation of rice (Oryza sativa L.)[J].Nat Biotechnol,1990,8(1):33-38.
[25] HIEI Y,OHTA S,KOMARI T,et al.Efficient transformation of rice (Oryza sativa L.)mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA[J].Plant J,1994,6(2):271-282.
[26] HIEI Y,KOMARI T.Agrobacterium-mediated transformation of rice using immature embryos or calli induced from mature seed[J].Nat Protocol,2008,3(3):824-834.
[27] OZAWA K,KAWAHIGASHI H.Positional cloning of the nitrite reductase gene associated with good growth and regeneration ability of calli and establishment of a new selection system for Agrobacterium-mediated transformation in rice(Oryza sativa L.)[J].Plant Sci,2006,170(2):384-393.
[28] DUAN Yongbo,ZHAI Chenguang,LI Hao,et al.An efficient and high-throughput protocol for Agrobacterium-mediated transformation based on phosphopmannose isomerase positive selection in Japonica rice (Oryza sativa L.)[J]. Plant Cell Rep,2012,31(9):1611-1624.
[29] KARTHIKEYAN A,PANDIANS S K,RAMESH M.Agrobacterium-mediated transformation of leaf base derived callus tissues of popular indica rice(Oryza sativa L.subsp.indica cv.ADT 43)[J].Plant Sci,2011,181(3):258-268.
[30] OZAWA K,TAKAIWA F.Highly efficient Agrobacterium-mediated transformation of suspension-cultured cell clusters of rice(Oryza sativa L.)[J].Plant Sci,2010,179(4):333-337.
[31] LEE K W,CHOI G J,KIM K Y,et al.Genotypic variation of Agrobacterium-mediated transformation of Italian ryegrass[J].Electron J Biotechnol,2010,13(3):8-9.
[32] SAHOO K K,TRIPATHI A K,PAREK A,et al.An improved protocol for efficient transformation and regeneration of diverse indica rice cultivars[J].Plant Methods,2011,7(1):49.doi:10.1186/1746.4811.7.49.
Transformation of the bar gene into embryogenic calli of rice via Agrobacterium tumefaciens
HE Shaohai,YAO Shengcun,FENG Yukai,WANG Ke,LI Feifei
(The Key Laboratory for Quality Improvement of Agricultural Products of Zhejiang Province,School of Agriculture and Food Science, Zhejiang A&F University,Lin’an 311300,Zhejiang,China)
To establish the embryogenic calli(EC)transformation system of rice with Basta as a selectable agent,the glufosinate-resistant gene (bar gene)was transformed into EC of rice cultivars, ‘Nipponbare’,‘Xiushui 134’ and ‘Zhonghua 11’ by Agrobacterium-mediated transformation.After 3 cycles of glufosinate-resistant screening with 15,10 and 10 mg·L-1glufosinate,transgenic EC was detected by PCR.The transformation efficiency of ‘Nipponbare’,‘Xiushui 134’ and ‘Zhonghua 11’ was 77.9%,67.5%and 92.2%respectively.The results showed that the selectable concentrations were effective for resistant EC screening and the resistant EC subsequently differentiated into seedlings.The frequency of EC differentiation of‘Zhonghua 11’ was 86.7%and the seedling rate was 30.7%;the frequency of EC differentiation of‘Xiushui 134’ was 44.4%,and the seedling rate was 66.7%;the frequency of EC differentiation of‘Nipponbare’ was 76.1%;and the seedling rate was 65.6%.Therefore,of the 3 cultivars,the EC of‘Nipponbare’ for genetic transformation with glufosinate as a selectable agent was most efficient.The glufosinate selection transformation system not only provides a technical basis for rice genetic transformation and functional validation,but also provides transgenic materials with herbicide resistance for rice breeding.[Ch,5 fig.3 tab.32 ref.]
molecular biology;glufosinate;bar gene;embryogenic calli(EC);Oryza sativa;Agrobacterium-mediated transformatiom
S511;Q785
A
2095-0756(2017)01-0129-08
2016-02-22;
2016-05-25
國家自然科學基金青年基金資助項目(31401461);浙江省大學生科技創新活動計劃(新苗人才計劃)項目(2015R412002);2015年浙江農林大學農學院大學生創新訓練計劃項目
何少海,從事水稻轉基因研究。E-mail:642836550@qq.com。通信作者:李飛飛,副教授,從事植物基因工程研究。E-mail:lifei-fei@163.com
浙 江 農 林 大 學 學 報,2017,34(1):137-144
Journal of Zhejiang A&F University
10.11833/j.issn.2095-0756.2017.01.019
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
