響應曲面法分析低鹽條件下溫度與pH對斜帶石斑魚幼魚能量代謝的聯合效應
2017-03-02 12:15:59黃建盛張健東湯保貴王忠良
海洋漁業 2017年1期
黃建盛,陳 剛,張健東,湯保貴,王忠良,周 暉
(廣東海洋大學水產學院,湛江 524088)
響應曲面法分析低鹽條件下溫度與pH對斜帶石斑魚幼魚能量代謝的聯合效應
黃建盛,陳 剛,張健東,湯保貴,王忠良,周 暉
(廣東海洋大學水產學院,湛江 524088)
采用二因素五水平的中心復合設計和響應曲面分析方法,以溫度(21~33℃)和pH(6.5~8.5)為因子,以耗氧率和排氨率為響應指標,探討了在低鹽條件下溫度和pH對斜帶石斑魚(Epinephelus coioides)幼魚能量代謝的聯合影響。結果表明,本實驗條件下,隨著溫度的上升,耗氧率(MO2)和排氨率(MTAN)呈上升的變化;隨著pH的上升,MO2和MTAN呈先上升后下降的變化。溫度一次效應對幼魚的MO2影響極顯著(P<0.01),二次效應對幼魚的MO2影響顯著(P<0.05);pH一次效應對幼魚MO2影響顯著(P<0.05),二次效應影響極顯著(P<0.01);溫度與pH互作效應對MO2影響顯著(P<0.05)。溫度一次效應對幼魚的MTAN影響極顯著(P<0.01),二次效應對幼魚的MTAN無顯著影響(P>0.05);pH一次效應和二次效應對幼魚MTAN影響顯著(P<0.05);溫度與pH互作效應對MTAN無顯著影響(P>0.05)。所考察因子與幼魚MO2、MTAN間二次多項回歸過程的決定系數分別為0.981 1、0.954 6(P<0.01),可用于分析與預測。通過對所建立的回歸方程同時優化,得到溫度與pH的最佳組合為:溫度31.2℃和pH 7.6。在此組合條件下,幼魚MO2、MTAN最大值分別為188.44 mgO2·(kg·hr)-1;7.01 mgTAN·(kg·hr)-1,可靠度為0.902。實驗結果提示,低鹽條件下斜帶石斑魚養殖可在本實驗中得到的最優因子組合下進行,以提高生產效益。
斜帶石斑魚;耗氧率;排氨率;低鹽水體;響應面法
魚類攝取食物后,營養物質經過一系列復雜的化學反應為機體提供能量,這一過程受到內在因素和外在因素的共同影響。水溫和水體pH是魚類生物能量學的重要影響因子,水溫主要通過改變魚體體溫,從而對魚體組織細胞內的各種酶活力產生影響,進一步影響魚體新陳代謝的水平;水體pH也是主要通過改變代謝酶活力而影響能量代謝[1]。關于溫度、pH對魚類能量代謝的研究已有一些報道,如,鮸(Miichthys miiuy)幼魚的耗氧率隨溫度升高而增大并呈指數相關,pH對鮸幼魚耗氧率無顯著影響[2];條石鯛(Oplegnathus fasciatus)幼魚耗氧率、排氨率隨溫度的升高而增大[3]。卵形鯧鲹(Trachinotus ovatus)幼魚隨著溫度的升高,耗氧率和排氨率均是先增大后減少;隨著pH的升高,耗氧率無顯著性變化,排氨率呈先增大后減少的趨勢[4]。上述這些研究結果揭示了溫度和pH對魚類能量代謝的變化規律。然而,上述的研究主要集中在單因素的溫度和pH幾個孤立的水平上對能量代謝的影響,沒有涉及環境因子之間的互作效應,同時,上述這些研究只分析了溫度或pH對能量代謝的一次效應,并未建立模型和考察溫度或水體pH的二次效應,使得研究結果缺乏可靠的預測性。
目前,斜帶石斑魚(Epinephelus coioides)作為廣東、海南、福建等地區的海水網箱養殖的主要品種之一正在蓬勃發展著,并成為沿海部分地區水產養殖經濟增長的支柱產業之一。然而,近年來為了避免沿海地區的惡劣環境因素對其生長造成的風險,逐漸發展池塘低鹽度養殖并取得成功。目前,關于水溫和水體pH對低鹽條件下斜帶石斑魚幼魚能量代謝的聯合效應尚未見有報道。本文在已有研究基礎上,應用中心復合試驗設計(central composite design,CCD)分析方法考察溫度與水體pH對斜帶石斑魚幼魚在低鹽環境下耗氧率和排氨率的聯合影響,并對各因子的二次效應及其交互作用進行分析。并結合響應曲面法(response surface methodology,RSM)分析耗氧率、排氨率隨溫度和pH的變化規律,建立相應模型,旨在為斜帶石斑魚低鹽養殖生理生態及養殖過程中水質管理提供一定的參考。
1 材料與方法
1.1 實驗魚和低鹽適應
實驗于2013年6月10日~8月10日于廣東海洋大學魚類實驗室進行,共60 d。實驗魚為廣東海洋大學魚類實驗室在廣東省湛江市東海島生物研究基地繁育的幼魚,選取健康、活力好的幼魚600 ind充氧運回實驗室,幼魚平均體質量23.67 g,平均體長10.3 cm。在實驗室水槽中暫養,水槽共40個,規格為70 cm×50 cm×60 cm,每個水槽15 ind魚,暫養期間,持續充氣,自然光照,每天8∶00時和16∶00時各飽食投喂1次,溶解氧保持在6 mg·L-1以上,水溫為(27.5 ±0.3)℃,pH 7.8,氨氮<0.01 mg·L-1,自然海水的鹽度為28~31,每天換水1次。暫養2周后,待幼魚攝食正常后每個水槽中開始加入經曝氣后的自來水,以每天2~4個鹽度單位逐漸降低水體鹽度至12左右,并保持在此鹽度下穩定適應1周。鹽度測定使用手持式鹽度計(HSS-100,上海精密儀器儀表有限公司)進行測定。每天換水一次,并盡量保持水體環境因子一致,換水量根據水質情況而定。
實驗所用的飼料為東丸石斑魚專用C6配合飼料(廣東越群海洋生物研究開發有限公司),近似營養成分:水分9.01%,粗蛋白52.94%,粗脂肪12.21%,粗灰分14.51%,粗纖維3.0%以下,鈣2.0%以下,磷1.8%以下。
1.2 實驗方法
1.2.1 實驗魚馴化
實驗魚經過低鹽適應后,先開始溫度馴化,采用漸變方式進行,每天溫度升降控制在2℃范圍之內,用電子恒溫棒和冰袋控溫,馴化至相應溫度后開始進行相應的pH馴化。水體pH也采用漸變方式調節,用鹽酸(0.2 mol·L-1)和氫氧化鈉(0.2 mol·L-1)調節,每天變幅不超過0.5,pH測定采用pH計(PHS-3C,上海精密儀器儀表有限公司)進行測定。馴化至相應pH后,穩定適應1周后開始實驗。
1.2.2 實驗設計
正式實驗前先進行預實驗,通過單因子實驗確定低鹽條件下斜帶石斑魚可以正常攝食和生長的水體溫度和pH范圍。實驗采用中心復合實驗設計,因子為溫度(21~33℃)和pH(6.5~8.5)。該實驗設計因子點數4,軸點數4,中心點數5,星號臂α=1.41,共計13次實驗組合,每個實驗組合重復3次。能量代謝為耗氧率及排氨率為響應指標,具體的溫度和pH組合見表1。
1.2.3 響應指標測定
耗氧率測定所用測定裝置是間歇性流水式呼吸儀(AQUARE SP,Qubit Systems Inc.,Canada,下文簡稱呼吸儀)(圖1)。此套系統測定過程包括測定、水體交換及等待3個階段,在測定階段,呼吸室完全封閉,測定時間可根據時間需要調節,通常最長以水中溶氧量下降不大于呼吸前溶氧的10%為基礎。測定階段結束后是水體交換階段啟動,以更新呼吸室內的水保持溶解氧飽和度,接著是短暫的滯后等待階段。如此往復,實現長時間實時測定。溶解氧探頭(OxyGuard Probes,德國)安裝在呼吸室里面,通過數據采集器連接到計算機,在計算機軟件(RESP-EDU)中可直接讀出耗氧率的數據。
將馴化至實驗設置的溫度-pH組合條件后,暫養的實驗魚禁食3 d(以確保胃排空)。禁食后隨機取實驗魚稱量濕重后,輕輕地轉入相應條件呼吸室內,待魚穩定適應后進行呼吸測定和水樣采集(用于測定氨氮)。從取魚稱重到轉入呼吸室過程時間在5 min內,而且盡量避免實驗魚反應過激。每個實驗組合梯度測定6 ind魚,有4套呼吸裝置,其中3套呼吸裝置放置1 ind魚,剩下的1套呼吸裝置不放置魚,留作空白對照組。耗氧率數據直接由呼吸儀在線測定讀取,至少讀取5個數據取平均值作為該尾魚耗氧率。呼吸儀持續測定每尾魚耗氧率,每次需要時間13~19 min,包括水體交換時間6~9 min,測定時間6~9 min,等待時間1 min。

圖1 間歇性流水呼吸系統原理結構圖Fig.1 Map of intermittent flow respirometry
排氨率測定用于測定排氨率的水樣采集時間與耗氧率測定時間相對應。在呼吸室進水時間點(測起始氨氮含量)和出水時間點(測終末氨氮含量)迅速取10 mL的水樣用于氨氮測定。氨氮測定用Test Kits氨氮測試盒(默克公司,原理:水楊酸分光光度法)采用分光光度計(6600,UVVIS,德國)在340 nm光波下測定。一般而言,氨態氮是氮排泄的主要物質,故不考慮其它形式的氮化合物。排氨率的計算公式如下:
式中,V是呼吸室的有效體積(L);△ATAN是在測定期間氨氮質量濃度的變化量(mg·L-1);△T是測定時間(h);W是幼魚濕重(kg)。
氧氮比計算根據測定的耗氧率、排氨率的數據計算氧氮比。計算公式如下:
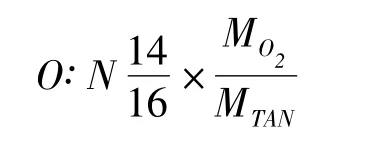
1.2.4 數據處理
數據測定結果用平均值±標準差(Mean± SD)表示,通過最小二乘法擬合因子與響應值之間的二次多項回歸方程為:
Y=b0+b1T+b2pH+b3T×pH+b4T2+b5pH2式中,Y是響應變量(在此響應為耗氧率或排氨率);b0為回歸常數;b1為溫度的一次效應;b2為pH的一次效應;b3為溫度與pH間的互作效應;b4為溫度的二次效應;b5為pH的二次效應。
采用Design-Expert(v8.0)軟件進行實驗設計與統計分析,對溫度、pH與響應值進行二次多項回歸擬合并繪制響應曲面圖和等高線圖。并對回歸模型方差分析和系數進行顯著性檢驗,當P<0.05時認為差異顯著,P<0.01時認為差異極顯著。
2 結果與分析
2.1 耗氧率
溫度與pH不同組合對低鹽水體下斜帶石斑魚幼魚耗氧率的測定結果見表1,并對所測定的結果采用最小二乘法進行回歸擬合,得到幼魚耗氧率與溫度、pH之間實際的二次多項回歸模型為:
MO2=-1 644.19+31.31T+341.46pH-1.84T×pH-0.20T2-19.15pH2
式中,R2=0.981 1,校正系數R2=0.967 6,說明回歸模型能解釋98.11%響應值的變化,僅有總變異的1.89%不能用此模型解釋,精密度(adeq precisior)是指有效信號與噪聲的比值,大于4.0視為合理,耗氧率模型精密度達到27.284,因此說明回歸模型擬合程度良好。
對幼魚耗氧率回歸模型方差分析進行顯著性檢驗,結果見表2。表2中的數據顯示,回歸模型F=72.77,P=0.000 1<0.01,表明回歸模型極顯著;失擬項F=0.36,P=0.784 2>0.05,說明所建立的模型有效。因此,可以用上述回歸模型來分析和預測溫度和pH對幼魚的耗氧率的結果。
幼魚耗氧率回歸模型系數顯著性檢驗結果見表3。表3中數據顯示,溫度一次效應對幼魚的耗氧率有極顯著影響(P<0.01),二次效應對幼魚的耗氧率有顯著影響(P<0.05);pH一次效應對幼魚耗氧率有顯著影響(P<0.05),二次效應有極顯著影響(P<0.01);溫度與pH互作效應對耗氧率有顯著影響(P<0.05)。
溫度與pH和幼魚的耗氧率之間的響應曲面及其等高線圖見圖2所示。通過圖2能直觀地評價溫度與pH對幼魚耗氧率的相互作用,等高線的性狀可反映出兩因子交互效應的強弱,橢圓形表示兩因子交互效應顯著,而圓形則表示兩因子交互效應不顯著。在本實驗條件下,幼魚耗氧率在溫度為21~33℃范圍內隨著溫度的上升而上升;在pH為6.5~7.5范圍內隨著pH的上升而上升,pH高于7.5則開始下降;圖2-A中等高線為橢圓形反映溫度與pH對幼魚的耗氧率有顯著的互作效應(P<0.05)。
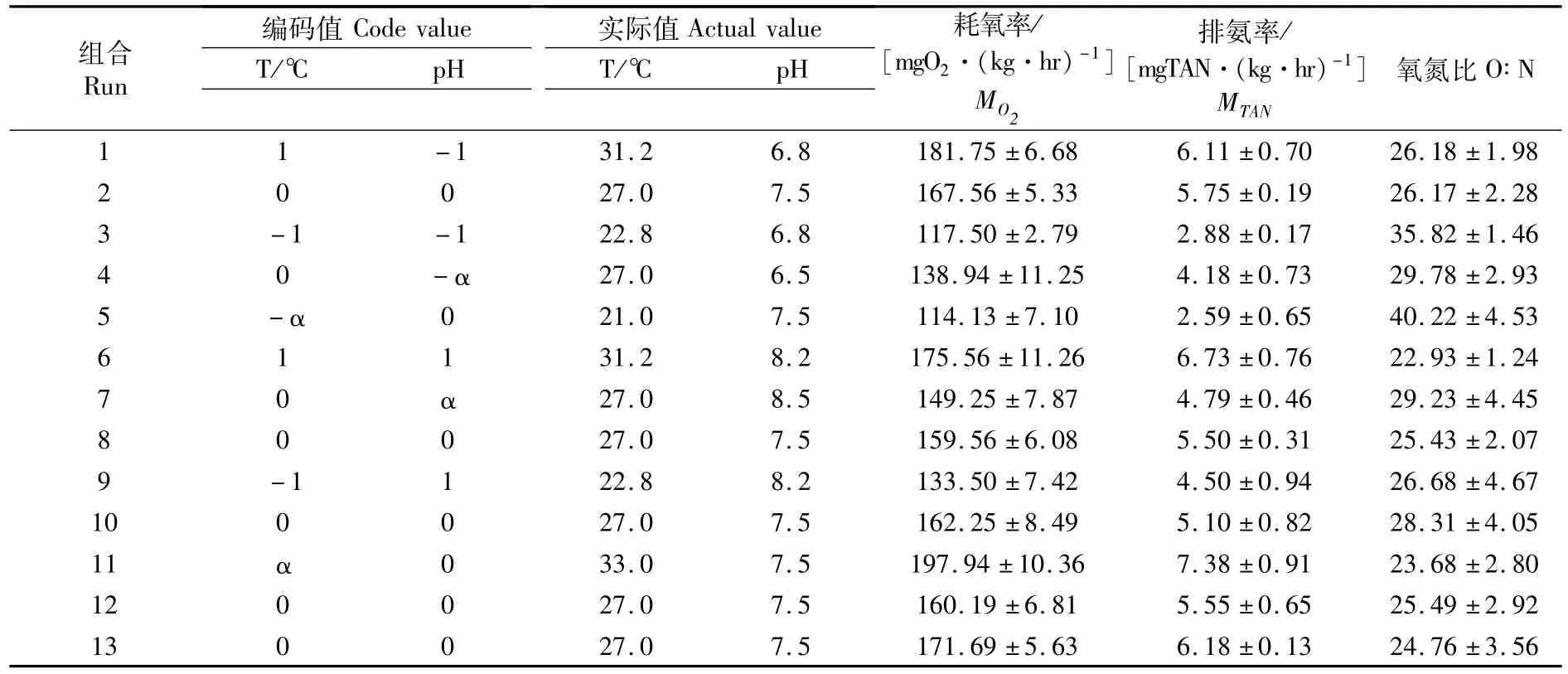
表1 溫度與pH的實驗設計組合及結果Tab.1 Experimental design of temperature(T)and pH and results
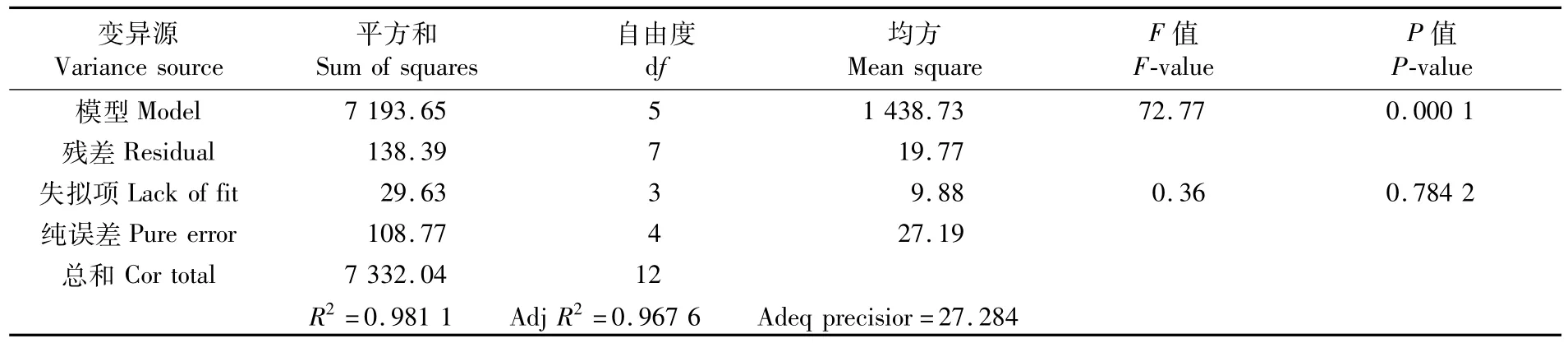
表2 溫度與pH對斜帶石斑魚幼魚耗氧率的回歸模型方差分析Tab.2 Variance analysis for the effects of temperature(T)and pH on oxygen consumption rate of juvenile E.coioides under low-salinity culture
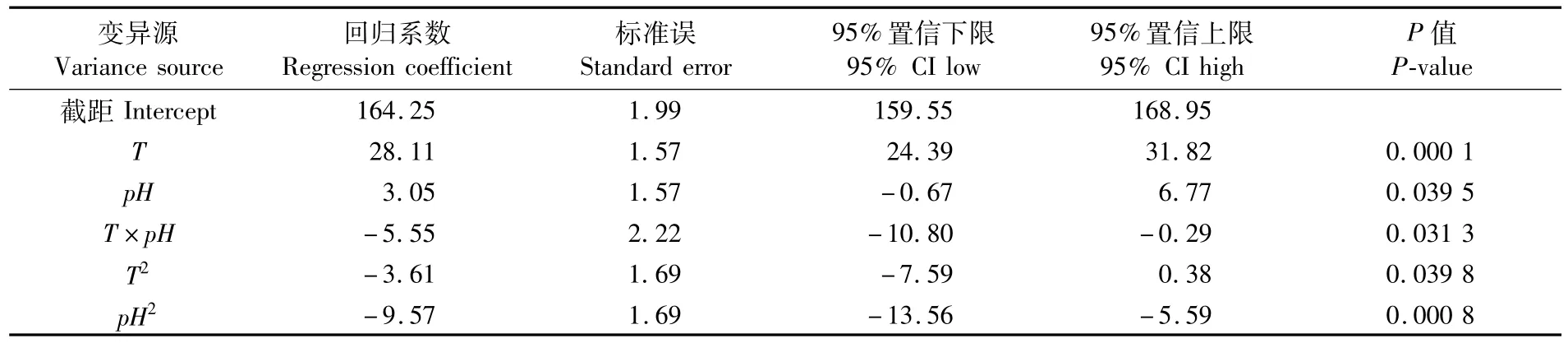
表3 溫度與pH對斜帶石斑魚幼魚耗氧率的模型回歸方程系數顯著性檢驗Tab.3 Test of significance for regression coefficient of temperature(T)and pH on oxygen consumption rate of juvenile E.coioides under low-salinity culture
2.2 排氨率
溫度與pH不同組合對低鹽環境下斜帶石斑魚幼魚排氨率的測定結果見表1,并對所測定的結果采用最小二乘法進行回歸擬合,得到幼魚排氨率與溫度、pH之間實際的二次多項回歸模型為:
MTAN=-89.46+1.70T+17.39pH-0.08T ×pH-0.01T2-0.97pH2
式中,R2=0.954 6,校正系數R2=0.905 6,說明回歸模型能解釋95.46%響應值的變化,僅有總變異的4.54%不能用此模型解釋,精密度(adeq precisior)是指有效信號與噪聲的比值,大于4.0視為合理,排氨率模型精密度達到15.852,因此說明排氨率回歸模型擬合程度良好。
對幼魚排氨率回歸模型方差分析進行顯著性檢驗,結果見表4。表4中的數據顯示,回歸模型F=24.33,P=0.000 3<0.01,表明回歸模型極顯著;失擬項F=1.41,P=0.363 6>0.05,說明所建立的模型有效。因此,可以用上述回歸模型來分析和預測溫度和pH對幼魚的排氨率的結果。
幼魚排氨率回歸模型系數顯著性檢驗結果見表5。表5中數據顯示,溫度一次效應對幼魚的排氨率有極顯著影響(P<0.01),二次效應對幼魚的排氨率無顯著影響(P>0.05);pH一次效應和二次效應對幼魚排氨率有顯著影響(P<0.05);溫度與pH互作效應對排氨率無顯著影響(P>0.05)。
溫度與pH和幼魚的排氨率之間的響應曲面及其等高線見圖3所示。通過圖3能直觀地評價溫度與pH對幼魚排氨率的相互作用。在本實驗條件下,幼魚排氨率在溫度為21~33℃范圍內隨著溫度的上升而上升;在pH為6.5~7.5范圍內隨著pH的上升而上升,pH高于7.5則開始下降;圖3-A中等高線為橢圓形反映溫度與pH對幼魚的排氨率無顯著的互作效應(P>0.05)。
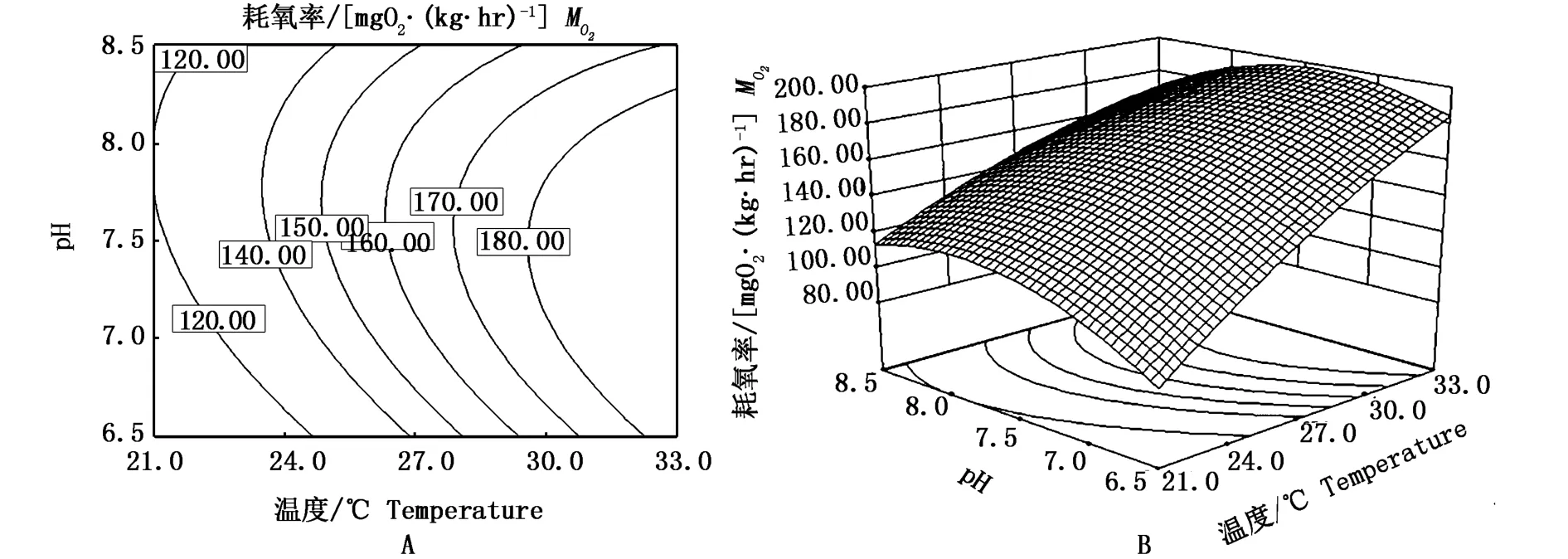
圖2 溫度與pH及其交互作用對低鹽條件下斜帶石斑魚幼魚耗氧率影響的等高線(A)和響應面(B)Fig.2 Response of contour plot(A)and its surface plot(B)of the effects of temperature(T)and pH and their mutual interactions on oxygen consumption rate of juvenile E.coioides under low-salinity culture
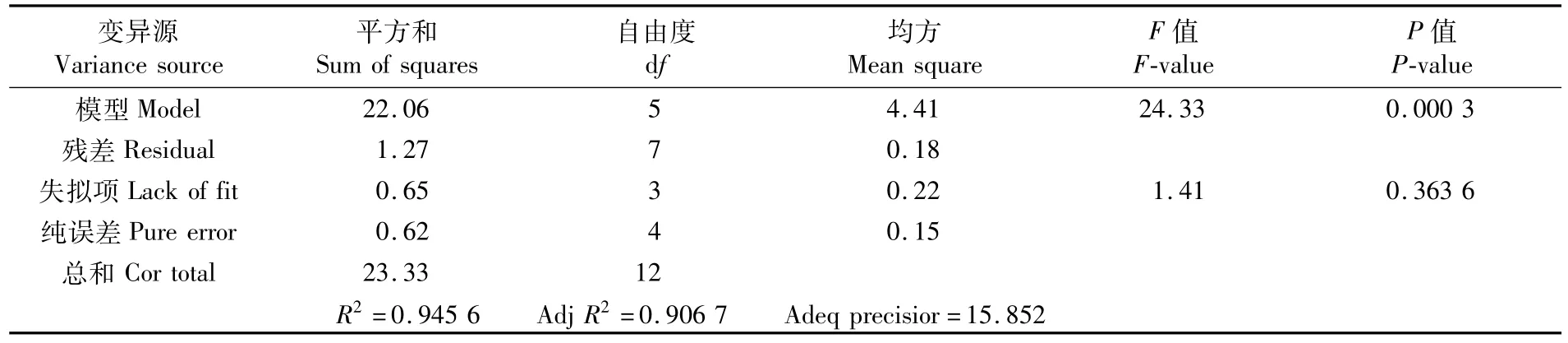
表4 溫度與pH對斜帶石斑魚幼魚排氨率的回歸模型方差分析Tab.4 Variance analysis for the effect of temperature(T)and pH on ammonia excretion rate of juvenile E.coioides under low-salinity culture
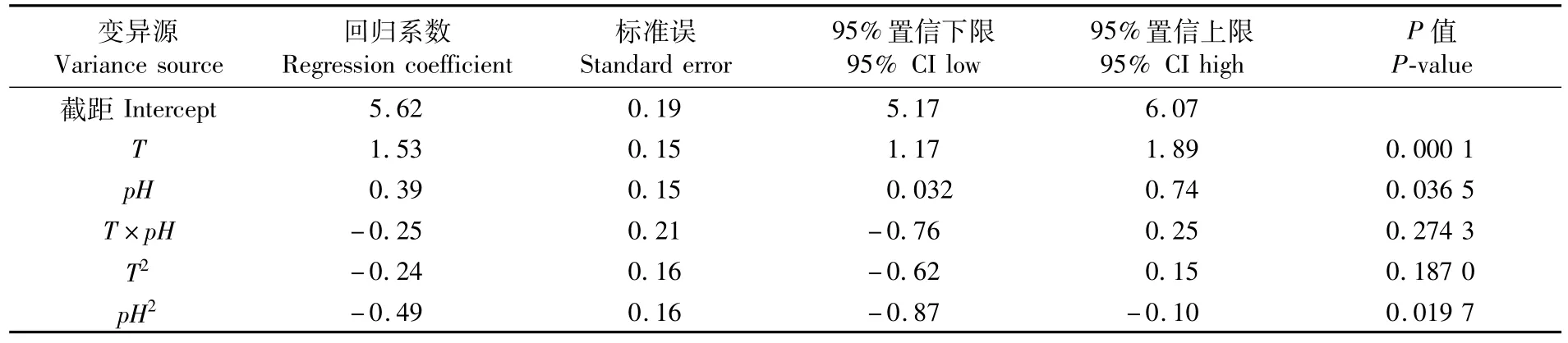
表5 溫度與pH對斜帶石斑魚幼魚排氨率的模型回歸方程系數顯著性檢驗Tab.5 Test of significance for regression coefficient of temperature(T)and pH on ammonia excretion rate of juvenile E.coioides under low-salinity culture
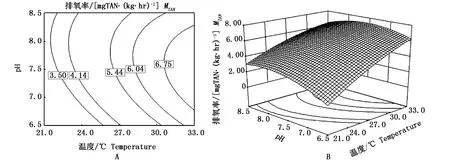
圖3 溫度與pH及其交互作用對低鹽條件下斜帶石斑魚幼魚排氨率影響的等高線(A)和響應面(B)Fig.3 Response of contour p lot(A)and its surface p lot(B)of the effects of temperature(T)and pH and their mutual interactions on ammonia excretion rate of juvenile E.coioides under low-salinity culture
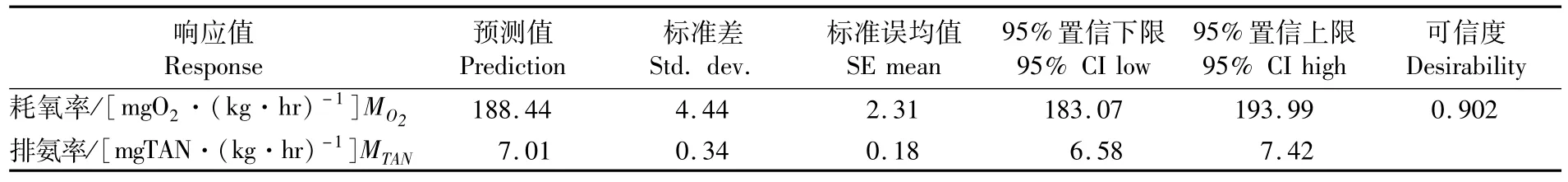
表6 溫度與pH對低鹽條件下斜帶石斑魚幼魚耗氧率和排氨率影響的優化置信區間Tab.6 Optimization of confidence interval of the effects of temperature and pH on ammonia excretion rate of juvenile E.coioides under low-salinity culture
2.3 氧氮比
斜帶石斑魚幼魚氧氮比的計算結果見表1。幼魚在溫度21~33℃范圍和pH 6.5~8.5范圍,不同組合條件下其氧氮比值范圍為22.93~40.22。
2.4 響應值優化
參考MONTGOMERY的方法[5],對幼魚的耗氧率、排氨率的最適溫度和pH進行同時優化(表6),結果顯示,溫度和pH的最優組合為31.2℃和7.6,此時幼魚耗氧率、排氨率最大值為188.44 mgO2·(kg·hr)-1、7.01 mgTAN·(kg·hr)-1,可靠度為0.902。
3 討論
溫度與pH作為魚類生長發育的重要環境因子,直接影響魚類能量代謝[2-4,6-11]。研究認為,溫度和pH主要通過影響魚體組織細胞內代謝酶活力而影響能量代謝[3]。本實驗結果表明,采用中心復合設計是一種有效的實驗設計與統計分析的技術,所得的回歸方程精度高,具有可預測性。在本實驗條件下,溫度與pH對低鹽環境下的斜帶石斑魚幼魚的耗氧率、排氨率均有顯著的影響,二次回歸模型決定系數分別為0.981 1、0.945 6和0.988 0,方程擬合較好,可以進行分析與預測。
溫度對魚類代謝酶活性起控制作用[12]。本實驗中,pH為6.5~8.5范圍,溫度為21~33℃范圍,幼魚的耗氧率、排氨率均隨著溫度的上升而上升。這說明在該溫度范圍,隨著溫度的上升,斜帶石斑魚幼魚維持生命代謝的各種組織器官活性增強,代謝酶活性增大,因此表現為耗氧率升高、產生的代謝產物氨氮增多的現象。這也符合魚類能量代謝隨溫度變化的一般規律。與鮸[2]、條石鯛[3]、黃鱸(Percaf luviatilis)[6]、大口黑鱸(Micropterus salmoides)[13]等研究結果相類似。
水體pH對魚類的耗氧率的影響,目前存在兩種不同觀點。第一種觀點認為,耗氧率隨水體pH升高而升高,當pH超過適宜范圍,耗氧率則隨之下降。在紅鮑(Haliotis rubra)[14]、雜交鱘(Huso huso♀×Acipenser baerii♂)[15]、胖頭鱥(Pimephales promelas)[16]的研究結果支持第一種觀點。第二種觀點認為,水體pH在一定范圍內變化,對耗氧率無顯著性影響。如,對日本黃姑魚(Nibea japonica)[8]、鮸[2]的研究結果支持第二種觀點。這反映了魚類隨著生活環境、生態習性及發育階段不同對pH的調節能力存在著差異。本實驗發現,pH對低鹽環境下的斜帶石斑魚幼魚的耗氧率有顯著影響,隨pH的升高而上升,pH超過7.5則開始下降,說明適宜pH范圍內,維持生命代謝的組織器官的代謝酶活性增強,基礎代謝旺盛,表現為耗氧率的升高,而pH超過7.5可能會引起水中一些化學物質含量的變化,阻礙魚類的呼吸,表現為耗氧率下降,但下降的幅度較小,是否會引起代謝紊亂還有待進一步研究證實。雖然魚類具備一定的酸堿平衡調節能力,但是從能量利用的角度來看,對于魚類長期處于酸堿平衡的調節耗能狀態,勢必導致其它生理功能的降低。上述研究結果及本實驗結果都是利用鹽酸驅動的水體酸化進行的,這與CO2驅動的海水酸化的結果會存在差異,這是因為CO2驅動的海水酸化不僅增加氫離子的含量,而且增加了碳酸氫根離子的濃度,降低了碳酸根離子的濃度,改變了海水的化學性質[17]。另有研究將真鯛(Pagrus major)的受精卵及仔魚分別在由鹽酸和CO2驅動相同pH值的海水中進行實驗,結果發現與鹽酸組死亡率為1%~4%,而CO2組表現為更高的死亡率,為60%~100%,說明不是海水pH值降低而是CO2產生了碳效應導致了真鯛仔魚的大量死亡[18]。關于CO2驅動和鹽度驅動的水體pH下降對斜帶石斑魚幼魚能量代謝的影響差異,需進一步探究。
以往的研究表明,環境因子對魚類的生長發育有重要的影響。但環境因子之間的互作效應更應受到關注。在因子互作效應存在的情況下,往往比因子主效應更重要[5]。在尼羅羅非魚(Oreochromis niloticus)[10]、大菱鲆(Scophthalmus maximus)[19]、犬齒牙鲆(Paralichthys dentatus)[20]、花狼魚(Anarhichas minor)[21]等研究中證實環境因子對魚類生長有明顯的互作效應。本實驗中,斜帶石斑魚幼魚耗氧率、排氨率回歸模型系數顯著性檢驗結果顯示(表3、表5),溫度和pH的互作效應對幼魚耗氧率顯著影響(P<0.05),這說明溫度與水體pH對幼魚的耗氧率是緊密關聯的。可能原因是溫度與水體pH對酶活力的影響存在關聯。pH影響酶活力主要有兩個途徑,一是改變酶的構象;二是改變酶與底物的結合,而溫度影響酶活力主要是通過改變酶與底物的結合,溫度與pH以相同的途徑影響酶活力。另外,溫度會改變水中的二氧化碳含量,從而影響水體的pH,因此溫度與pH存在著明顯的互作效應。而溫度和pH的互作效應對幼魚排氨率卻無顯著影響(P>0.05),可以推測溫度和pH對氨氮排泄的影響是獨立進行的,其確切的機理還待深入研究。
響應曲面法最大的優點在于把因素和實驗結果的關系函數化,可以研究因子與響應值之間、因子與因子之間的相互關系,并進行優化,找到最佳因子組合[22]。然而,國內外學者關于環境因子對魚類能量代謝影響的研究,大部分只針對某個環境因子的幾個孤立水平點進行分析,具有很大局限性。而傳統正交試驗方法雖能尋找最佳因子水平組合,但不能建立因子與響應值間明確的函數關系,從而無法找到因子最佳組合和最優響應值。本實驗中,采用響應曲面法建立可靠的回歸模型,可對響應進行連續的預測。方差分析結果(表2、表4)顯示,回歸模型均極顯著(P<0.01),回歸方程決定系數高,模型失擬項檢驗結果不顯著(P>0.05),說明建立的模型有意義,可用于分析與預測。并獲得模型優化后得出最優的因子組合為溫度31.2℃、pH 7.6,在此組合下,幼魚耗氧率、排氨率的最大值為188.44 mgO2·(kg·hr)-1、7.01 mgTAN·(kg·hr)-1。一般而言,魚類耗氧率、排氨率表現較高的現象,反映了魚類的新陳代謝旺盛,生存環境適宜,有利于魚類生長。說明低鹽條件下的斜帶石斑魚幼魚在溫度31.2℃、pH 7.6左右時,生長性能較好。
應用響應面法分析水生動物能量代謝的聯合效應,該方法具有實驗次數少、靈活、方便、全面考察各因素及因素間的交互作用等優點,在實際應用中大大節約了實驗時間及費用,可提前達到預期的實驗目的,為我們提供了一種新的數學分析方法。在斜帶石斑魚低鹽養殖生理生態及養殖的水質管理過程中,還涉及其它因子的影響,如氨氮、亞硝酸鹽、養殖密度、光照強度等。因此,還有待進一步深入研究這些因子的影響及因子間是否存在互作效應,以便更好地指導生產實踐。
[1] 強 俊,王 輝,李瑞偉,等.酸堿脅迫對不同體質量尼羅羅非魚幼魚能量代謝的影響[J].應用生態學報,2011,22(9):2438-2446.QIANG J,WANG H,LI R W,et al.Effects of acid and alkaline stress on energy metabolism of Oreochromis niloticus juveniles with different body mass[J].Chinese Journal of Applied Ecology,2011,22(9):2438-2446.
[2] 閆茂倉,單樂州,邵鑫斌,等.溫度及體重對鮸魚幼魚耗氧率和排氨率的影響[J].熱帶海洋學報,2007,26(1):44-49.YAN M C,SHAN L Z,SHAO X B,et al.Influences of temperature and weight on respiration and excretion of Miichthymiiuy juvenile[J].Journal of Tropical Ocean Nography,2007,26(1):44-49.
[3] 閆茂倉,單樂州,謝起浪,等.溫度,鹽度及體重對條石鯛幼魚耗氧率和排氨率的影響[J].海洋科學進展,2009,26(4):486-496.YAN M C,SHAN L Z,XIE Q L,et al.Influence of temperature,salinity and body weight on oxygen consumption and ammonia excretion of Oplegnathus fasciatus juvenile[J].Advances in Marine Science,2009,26(4):486-496.
[4] 王 剛,李加兒,區又君,等.環境因子對卵形鯧鲹幼魚耗氧率和排氨率的影響[J].動物學雜志,2012,46(6):80-87. WANG G,LI J E,OU Y J,et al.Influence of environmental factors on oxygen consumption and ammonia excretion of juvenile Trachinotus ovatus[J].Chinese Journal of Zoology,2012,46(6):80-87.
[5] MONTGOMERY D C.Design and analysis of experiments[M].New York:John Wiley and Sons,2008.
[6] ZAKE S′Z,DEMSKA-ZAKE S′K,KATA K.Rates of oxygen consumption and ammonia excretion of juvenile Eurasian perch Perca fluviatilis L.[J].Aquaculture International,2003,11(3):277-288.
[7] 曾令清,張耀光,付世健,等.雙向急性變溫對南方鲇幼魚靜止耗氧率和臨界游泳速度的影響[J].水生生物學報,2011,35(2):276-282.ZENG LQ,ZHANG Y G,FU S J,et al.Effects of acute temperature change on resting oxygen consumption rate and critical swimming speed in juvenile southern catfish(Silurus meridionalis CHEN)[J].Acta Hydrobiologica Sinica,2011,35(2):276-282.
[8] 柴學軍,胡則輝,徐君卓,等.鹽度和pH對日本黃姑魚幼魚耗氧率和排氨率的影響[J].浙江海洋學院學報(自然科學版),2009,28(2):146-150.CHAIX J,HU Z H,XU J Z,et al.Effect of salinity and pH on oxygen consumption rate and ammonia excretion rate in juvenile Nibea japonica[J].Journal of Zhejiang Ocean University(Natural Science),2009,28(2):146-150.
[9] 唐道軍,徐善良,馬 斌.溫度對黑鱾幼魚耗氧率和排氨率的影響[J].動物學雜志,2013,48(2):256-260.TANG D J,XU S L,MA B.Influence of temperature on oxygen consumption and ammonia excretion of juvenile Girella melanichthys[J].Chinese Journal of Zoology,2013,48(2):256-260.
[10] 王 輝,強 俊,王海貞,等.溫度與鹽度對吉富品系尼羅羅非魚幼魚能量代謝的聯合效應[J].中國水產科學,2012,19(1):51-61.WANG H,QIANG J,WANG,H Z,et al.Combined effect of temperature and salinity on energy metabolismof GIFT Nile tilapia(Oreochromis niloticus)juveniles[J].Journal of Fishery Sciences of China,2012,19(1):51-61.
[11] 閆茂倉,單樂州,邵鑫斌,等.鹽度和pH值對鮸魚幼魚耗氧率和氨氮排泄率的影響[J].臺灣海峽,2007,26(1):85-91.YAN M C,SHAN L Z,SHAO X B,et al.Influence of salinity and pH on the rates of oxygen consumption and ammonia excretion of juvenile Miichthy mijuy[J].Journal of Oceanography in Taiwan Strait,2007,26(1):85-91.
[12] LIKONGWE JS,STECKO T D,STAUFFER JR,et al.Combined effects of water temperature and salinity on growth and feed utilization of juvenile Nile tilapia Oreochromis niloticus(Linneaus)[J].Aquaculture,1996,146(1):37-46.
[13] 徐如衛,江錦坡,肖建忠.大口黑鱸仔、稚魚窒息點與耗氧率的初步測定[J].浙江水產學院學報,1994,13(2):110-115.XU R W,JIANG J B,XIAO J Z.A preliminary measurement on suffocation point and oxygen consumption rate of Micropterus salmoides[J].Journal of Zhejiang College of Fisheries,1994,13(2):110-115.
[14] HARRIS JO,MAGUIRE G B,EDWARDSS J,et al.Effect of pH on growth rate,oxygen consumption rate,and histopathology of gill and kidney tissue for juvenile greenlip abalone,Haliotis laevigata donovan and blacklip abalone,Haliotis rubra leach[J].Journal of Shellfish Research,1999,18(2):611-619.
[15] 吳常文,朱愛意,趙向炯.海水養殖雜交鱘耗氧量、耗氧率和窒息點的研究[J].浙江海洋學院學報(自然科學版),2005,24(2):100-104.WU CW,ZHU A Y,ZHAO X J.Study on oxygen uptake,oxygen uptake rate and asphyxiation point of mari-culture hybrid sturgeon[J].Journal of Zhejiang Ocean University(Natural Science),2005,24(2):100-104.
[16] IGRAM R,WARESII W D.Oxygen consumption in the fathead minnow(Pimephales promelas rafinesque)—II effects of pH,osmotic pressure,and light level[J].Comparative Biochemistry and Physiology Part A:Physiology,1979,62(4):895-897.
[17] 劉洪軍,張振東,官曙光,等.海洋酸化效應對海水魚類的綜合影響評述[J].生態學報,2012,32(10):3233-3239.LIU H J,ZHANG ZD,GUAN SG,et al.A review of comprehensive effect of ocean acidification on marine fishes[J].Acta Ecologica Sinica,2012,32(10):3233-3239.
[18] KIKKAWA T,KITA J,ISHIMATSU A.Comparison of the lethal effect of CO2and acidification on red sea bream(Pagrus major)during the early developmental stages[J].Marine Pollution Bulletin,2004,48(1):108-110.
[19] IMSLAND A K,FOSS A,GUNNARSSON S,et al.The interaction of temperature and salinity on growth and food conversion in juvenile turbot(Scophthalmus maximus)[J].Aquaculture,2001,198(3):353-367.
[20] MALLOY K D,TARGETT T E.Feeding,growth and survival of juvenile summer flounder Paralichthys dentatus:Experimental analysis of the effects of temperature and salinity[J].Marine Ecology Progress Series,1991,72(3):213-223.
[21] MAGNUSSEN A B,IMSLAND A K,FOSS A.Interactive effects of different temperatures and salinities on growth,feed conversion efficiency,and blood physiology in juvenile spotted wolffish,Anarhichas minor Olafsen[J].Journal of World Aquacult Society,2008,39(6):804-811.
[22] 國 蓉,李劍君,國 亮,等.采用響應曲面法優化甘草飲片中甘草酸的超聲提取工藝[J].西北農林科技大學學報(自然科學版),2006,34(9):187-192.GUO R,LI J J,GUO L,et al.Optimization of the ultrasonic extraction technique of glycyrrhizic acid by using response surface methodology[J].Joural of Northwest Sci-ech University of Agriculture and Forestry(Natural Science),2006,34(9):187-192.
Combined effects of temperature and pH on energy metabolism of Epinephelus coioides juveniles cultured in low salinity using response surface methodology
HUANG Jian-sheng,CHEN Gang,ZHANG Jian-dong,TANG Bao-gui,WANG Zhong-liang,ZHOU Hui
(Fisheries College,Guangdong Ocean University,Zhanjiang 524088,China)
Orange-spotted grouper,which inhabit subtropical and tropical areas,have become a very popular cultured species of marine teleost currently in China.Metabolic rate is the most fundamental biological rate as it represents the rate of energy uptake,transformation and allocation.Oxygen consumption is a widely studied indicator of metabolic rate,and measurements of oxygen consumption rate are often used to examine energy utilization and stress in aquatic animals.We evaluated the combined effects of temperature and pH on the energy metabolism of Epinephelus coioides juveniles cultured in low salinity.Two-factor central composite experimental design and response surface methodology were used to test the effects of temperature(21-33℃)and pH(6.5-8.5)on the oxygen consumption rate(MO2)and ammonia excretion rate(MTAN)under laboratory conditions.Results showed that MO2and MTANincreased with the increasing temperature,and increased at the beginning then decreased with the increasing pH.The linear effects of temperature on MO2were highly statistically significant(P<0.01),the quadratic effects of temperature were significant(P<0.05);the linear effects of pH on MO2were significant(P<0.05),the quadratic effects of pH were highly statistically significant(P<0.01).Interaction effects of temperature and pH on MO2were significant(P<0.05).The linear effects of temperature on MTANwere highly statistically significant(P<0.01),but the quadratic effects of temperature were not significant(P>0.05);the linear and quadratic effects of pH on MTANwere significant(P<0.05);interaction effects of temperature and pH on MTANwere significant(P<0.05).The optimal temperature-pH combination was obtained through statistical optimization approach,and it was 31.2℃/7.6,at which the maximal MO2,MTANreached 188.44 mgO2·(kg·hr)-1,7.01 mgTAN·(kg·hr)-1respectively,with the desirability of 0.902.The data obtained suggested that producing profit can be improved by rearing at an optimal combination of temperature and pH of juvenile E.coioides under low-salinity culture.
Epinephelus coioides;oxygen consumption rate;ammonia excretion rate;low salinity water;response surface methodology
S 917
:A
1004-2490(2017)01-0041-10
2016-02-01
廣東省教育廳育苗工程(自然科學)項目(No.2013LYM-0038);廣東省海洋漁業科技推廣專項項目[No.A2010-08D04,A2010004-016(c)];廣東省海洋經濟創新發展區域示范專項“石斑魚等名貴海水魚工廠化循環水高效養殖產業化”(No.GD2012-A02-003)
黃建盛(1981-),男,廣東湛江人,漢族,博士,研究方向:水產經濟動物養殖生理生態學。E-mail:fishaqu@126.com
陳 剛,教授。Tel:0759-2382462,E-mail:cheng@gdou.edu.cn
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
核科學與工程(2021年4期)2022-01-12 06:30:26
當代陜西(2021年2期)2021-03-29 07:41:24
今日農業(2020年19期)2020-12-14 14:16:52
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
中學物理·高中(2016年12期)2017-04-22 11:53:03
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
中國塑料(2016年3期)2016-06-15 20:30:00
