不同喚醒度負性刺激抑制控制特點
2017-03-11 05:44:36趙紹晨鄭希付
心理研究 2017年1期
趙紹晨鄭希付
(1公安邊防部隊高等專科學校,廣州 510663;2華南師范大學心理學院,廣州 510631)
不同喚醒度負性刺激抑制控制特點
趙紹晨1鄭希付2
(1公安邊防部隊高等專科學校,廣州 510663;2華南師范大學心理學院,廣州 510631)
本研究運用事件相關電位(ERP)技術,采用雙選擇情緒oddball范式,探討不同喚醒度負性刺激的行為抑制特點。實驗選取15名大學生作為被試,要求被試對標準刺激和偏差刺激(75%vs.25%)分別作不同的按鍵反應,記錄按鍵前后的腦電。偏差刺激由高、低喚醒兩種來自國際情緒圖片系統的情緒圖片組成。本研究主要分析反應抑制過程的N2和P3的波幅和潛伏期。結果顯示:對高、低喚醒刺激的抑制反應誘發的N2波幅和潛伏期沒有差異,對高喚醒刺激抑制反應誘發的P3波幅大于低喚醒的P3波幅,并且差異顯著。結論:對高喚醒刺激進行抑制控制的結果是產生更大的P3波幅,這一過程的結果顯示,人類對高喚醒刺激更難進行抑制控制。
喚醒度;行為抑制;oddball;ERP
1 引言
行為抑制作為執行功能的一個重要方面,反映了個體對不恰當刺激行為反應的合理控制,在人類的生存適應中起著重要的作用。Barkley認為,行為抑制包括對優勢反應的抑制、停止正在進行的無效的或是與目標無關的行為、對內源性或外源性干擾的控制[1]。而行為抑制能力的缺失或異常常導致沖突或犯罪行為的發生[2]。近年來,關于行為抑制控制的研究發現,執行行為抑制任務的神經基礎是額葉-基底神經節回路[3],但是對額葉各區在抑制過程中的作用看法不一[4]。其中一種觀點認為,右側腹外側前額皮層(ventrolateral prefrontal cortex,VLPFC)負責抑制無關行為,而背側前額葉(dorsolateral prefrontal cortex,DLPFC)參與工作記憶,前扣帶回(anterior cingulate cortex,ACC)探測與目標不一致的沖突[5,6]。其中,右側VLPFC廣泛參與反應抑制、記憶提取抑制、任務定勢切換等抑制控制任務,因而被認為是具有一般抑制功能的腦區[7]。最近,Mostofsky和Simmonds[4,7]提出前部輔助運動區(pre-supplementary motor area,pre-SMA)是選擇適當的行為的關鍵區域,其中包括選擇投入到適當的運動反應和選擇抑制不適當的運動反應,而VLPFC參與維持工作記憶中的信息,DLPFC則參與操作工作記憶中的信息。另外,Hirokazu和Bekkker等研究者發現眶額葉皮層與前扣帶回在行為抑制活動的完成中具有重要作用[8]。也有相關的研究發現,在行為抑制過程中,前扣帶回的主要作用是沖突監控和錯誤監控[9,10]。采用go/nogo及雙選擇oddball范式的研究顯示,額-中央區N2、中央-頂區P3成分是與行為抑制控制過程密切相關的兩個ERP成分[11]。行為控制相關N2成分被認為主要與沖突覺察與監控有關;而行為控制相關P3的出現則是行為抑制加工過程的直接指標,比如,Go/Nogo任務中P3峰值的出現往往代表著行為抑制加工的完成[8,11]。
人類的情緒活動對行為抑制有著重要的影響[12,13],如負性情緒會導致攻擊行為的發生[14]。有研究顯示,沖動性人格特質與個體的情緒穩定性存在著顯著的負相關[15]。有行為失控癥狀的精神病患者,如邊緣型人格障礙患者、反社會人格障礙患者,在負性情緒下其行為控制缺陷癥狀更為明顯[16],說明個體的情緒狀態可能對行為控制過程產生影響。事實上,情緒與行為控制的交互作用近期得到了腦成像研究的支持[17-20]。
情緒是人格系統的動力核心,不同的情緒狀態會對行為、記憶、注意等活動有著不同程度的調節作用,負性情緒可引發對環境中負性刺激的注意偏向。Goldstein等提出負性情緒喚起個體的運動行為(為了生存而戰斗或逃跑),因此抑制行為反應變得困難。而在相關研究中發現,腦島前葉在行為抑制過程中受到情緒線索的選擇性激活[21,22]。隨著抑制難度的增大,負性情緒線索對左側腦島的激活也逐漸增大[23]。這些結果表明,腦島在情緒抑制任務中的激活反映了負性情緒行為抑制線索編碼需要更多的認知資源[14]。那么,同樣是負性情緒刺激,如果喚醒度不同,行為抑制會表現出什么樣的特點呢?
近年來,出現了大量關于情緒與行為抑制的研究[4,18,23,22],但研究一方面主要集中于外顯、內隱和情緒干擾任務的效價研究[21,24,25,26-31],另一方面主要集中于生理機制的研究[3,4,32,33-36]。 但是研究很少涉及情緒喚醒度的效應,盡管Goldstein與Shafritz等人的fMRI研究揭示了情緒條件下反應抑制的腦機制,卻難以回答神經結構的激活是來源于情緒加工本身,還是來源于情緒對反應抑制的影響[14,21]。相關研究發現,一定程度的消極情緒對任務加工有促進作用。而對于情緒刺激的研究大都集中于效價的維度上,關于消極情緒刺激對任務加工的影響尚未有進一步的研究,根據情緒的維度分類,對于同一種效價,喚醒度維度可能也會產生一定程度的影響。本研究采用高時間分辨率ERP技術,及情緒雙選擇oddball范式考察不同喚醒度負性情緒的行為抑制特點。
2 實驗方法
2.1 被試
15名在校大學生作為被試,其中7名男性,8名女性,年齡范圍在18~23歲,平均年齡約20歲,所有被試身心健康,均為右利手,視力正常或矯正后正常,均簽署了實驗知情同意書,實驗結束后給予少許報酬。
2.2 刺激材料
采用雙選擇oddball范式。在實驗中,刺激材料均分為標準刺激和偏差刺激 (其比例為75%vs 25%)兩類。標準刺激為一個杯子的自然景物圖片,偏差刺激為高喚醒度負性刺激,低喚醒度負性刺激各51張。所有刺激圖片均來自國際情緒圖片系統。負性圖片分為高喚醒度組和低喚醒度組。高低喚醒度圖片的效價保持一致,對高低兩種喚醒度圖片的效價進行t檢驗,t=1.07,差異不顯著;對兩組圖片的喚醒度進行比較,t=-3.63,p<0.00,差異十分顯著。其中高喚醒度組的效價M=3.43,SD=0.32;喚醒度M= 5.00,SD=0.76;低喚醒度組的效價M=3.54,SD=0.70;喚醒度M=4.36;SD=1.00。圖片分辨率均為100像素/英寸,大小統一為341像素×256像素。被試雙眼與屏幕距離約100cm。
2.3 ERP記錄
實驗儀器為Brain Product腦電記錄系統,參考電極置于雙側乳突連線,前額接地,采用32導電極帽記錄腦電,同時記錄水平眼電和垂直眼電,濾波帶通為DC-100 Hz,采樣頻率為500Hz/導,頭皮電阻降至5千歐以下。進行離線分析(off-line analysis),分析時程(epoch)為700ms,含基線100ms,自動矯正眨眼等偽跡,波幅大于±80μV者在疊加中被自動剔除。
2.4 程序和任務
實驗共有3個block,每個block包括136個刺激試次(trial)。各喚醒度條件刺激圖片呈現順序完全隨機。在實驗開始之前,告知被試本實驗考察個體的反應選擇及對優勢反應進行抑制的能力。首先,在電腦屏幕中央出現十字注視點,隨機持續時間為600~800ms。然后呈現1000ms的隨機刺激。被試的任務是當標準刺激出現時用左(右)手食指按F鍵反應;而當偏差刺激出現時,用右(左)手食指按J鍵反應。反應手進行block和被試間平衡。刺激呈現時間上限1000ms,刺激呈現隨被試按鍵終止。因此,當刺激呈現之后,被試需要在1000ms之內盡可能快速而準確的作按鍵反應,在練習階段提供被試對兩類刺激圖片的反應正確反饋。然后是不定間隔的注視點,以此循環。正式實驗之前被試先進行20個試次的練習,所有被試都必須在練習階段反應正確率達到100%再進入正式實驗。記錄標準刺激及各喚醒度偏差刺激誘發的EEG活動。實驗結束后,對各個被試進行有關實驗情況的訪談。
2.5 ERP數據分析和統計
分析時程為700ms,其中刺激呈現前100ms作為基線。對各喚醒度的負性偏差刺激及標準刺激誘發的ERP分別進行疊加和平均。由圖1可知,情緒喚醒度效應主要出現在頭皮中央及額-中央記錄區域。因此,本研究選擇以下的14個電極位置記錄的ERP波形用于統計分析:FZ,FCZ,CZ,PZ,FP1,FP2,F7,F3,F4,F8,FC5,CP5,CP6,FC6。偏差刺激與標準刺激誘發的ERP從大約100ms開始分離,該差異一直持續到大約500ms結束。這些差異在差異波上集中表現為與晚期抑制控制相關N2(210~300ms)及P3(380~430ms)成分。因此,本研究首先通過平均波幅分析在 210~300ms及 380~430ms兩窗口檢驗行為控制ERP效應的顯著性。分析因素為刺激類別(偏差刺激,標準刺激)與電極點(14個記錄點)。在行為控制效應顯著的基礎上,通過測量各喚醒度條件下N2及P3的潛伏期(刺激呈現到波峰之間的時間距)和波幅(基線到波峰),進一步分析負性情緒不同喚醒度對行為控制過程的影響。對上述各成分的波幅與潛伏期進行兩因素重復測量方差分析 (ANOVA)。重復測量因素為喚醒度(高,中,低)及電極(14個電極點)。采用Greenhouse-Geisser法矯正自由度以及P值。
3 實驗結果
3.1 行為結果
被試對標準刺激及偏差刺激的反應正確率均超過95.84%,對于正確率,刺激喚醒度及刺激類型主效應均不顯著。對反應時間,被試對偏差刺激的反應時間(M=426.49±52.35)顯著大于對標準刺激的反應時間(M=499.70±46.57),F(1,28)=16.38,p<0.000。此外,對于反應時間,喚醒度主效應顯著,F(2,42)=11.74,p<0.001。高喚醒條件下的反應時顯著大于中性條件下和低喚醒條件下的反應時間。被試在標準刺激、高喚醒的偏差刺激和低喚醒的偏差刺激條件 下的反應時間分別為:422.98ms,496.48ms,493.67ms。由此可知,被試在各偏差刺激條件下均出現了顯著的反應延遲效應,該效應在高喚醒刺激條件下尤為突出。
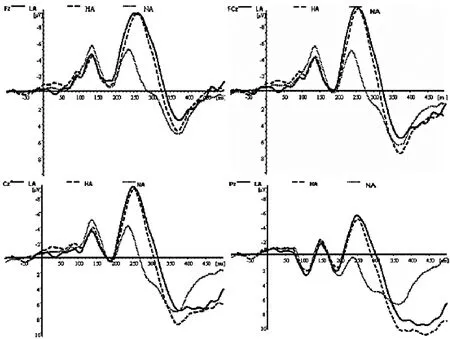
圖1 LA(低喚醒度),HA(高喚醒度),NA(中性圖片)
3.2 ERP成分與統計分析
對表現行為控制效應的原始波形而言,對N2在210~300ms時間窗的平均波幅重復測量方差分析顯示,刺激類別的主效應顯著[F(2,28)=11.20;p<0.01],電極點主效應顯著[F(13,182)=7.71;p<0.01]。偏差刺激誘發出比標準刺激更大的負偏向,該偏向在差異波中集中體現為顯著的額中央區N2活動。高、低喚醒刺激誘發的N2波幅差異不顯著。平均波幅在中前部記錄點顯著大于后部記錄點,說明出現了抑制效應。此外,刺激類別與電極點交互效應顯著[F(26,364)=4.06;p<0.01]。對不同條件的潛伏期進行重復測量方差分析顯示出顯著的刺激類別差異 [F(2,28)=16.60;p<0.000]。對P3在380~430ms時間窗的平均波幅統計分析表現出顯著的刺激類別 [F(2,28)=10.46;p<0.01]和電極點 [F(13,182)=2.22;p<0.05]主效應。其中高喚醒刺激的P3波幅大于低喚醒刺激的P3波幅。對不同條件的潛伏期進行重復測量方差分析,刺激類別差異不顯著。總的來講,中央-頂區的波幅顯著大于前部記錄區域的波幅。偏差刺激,尤其在中央區及后頂區,誘發出相比標準刺激更大的顯著正偏向,該偏向在差異波中表現為明顯的P3活動。由此可知,本實驗條件下,偏差刺激誘發出明顯的行為抑制效應,該效應集中體現為額-中央區域的N2活動差異及中央頂區P3活動差異(見圖1)。
4 討論
本研究采用雙選擇的oddball范式,結果發現,個體對偏差刺激的反應時間顯著長于對標準刺激的反應時間,說明被試對偏差刺激進行了行為抑制。同時也發現不同喚醒度的視覺情緒刺激對個體的行為控制過程有顯著影響。該影響在行為層面上表現為,隨著負性情緒喚醒度的增強,個體對偏差刺激反應延遲效應明顯,說明個體對偏差刺激的加工除了知覺注意、刺激辨別、反應決策與執行等過程,也包括對優勢反應的行為抑制。雙選擇oddball范式和go/no go范式都是oddball范式的變式,雙選擇oddball范式中對偏差刺激的反應相當于go/no go范式的no go任務,均是反應對優勢行為的抑制控制過程[37,38]。這一范式引發出兩個成分 N2 (200-400ms)和P3(300-500ms),一般在前額腦區波幅最明顯,并且no go引發的成分均小于go任務引發的成分[39]。No go的N2和P3成分反映了反應抑制的不同過程,N2是對認知過程的一個廣泛體現,并不是嚴格反映抑制控制過程本身。而P3成分則主要是對抑制控制過程的結果進行評估[38]。
以往研究發現,行為抑制任務中N2與反應沖突檢測有關,由反應沖突任務誘發的N2顯著大于無反應沖突的任務。本研究在210~300ms窗口觀測到顯著的中央區N2活動,統計結果顯示N2波幅喚醒度主效應顯著,高喚醒度誘發的波幅較大。由于N2波幅代表著沖突監控中的注意分配[37],這表明高喚醒的負性刺激最大程度的引起了對刺激監控的注意分配。另一方面,偏差刺激中高喚醒度和低喚醒度的負性刺激誘發的腦電比標準的中性刺激顯著延遲,由于潛伏期代表著反應沖突覺察的快慢[37],因此,個體對高喚醒度的刺激有更強的抑制。此外,N2還與反應抑制有關[11,38],代表抑制過程的早期階段,較大的N2即代表在nogo測試條件下中前扣帶回的活躍程度較大。時間壓力增大意味著反應趨勢的增大,N2-nogo也隨之增大。該結果提示高喚醒度負性刺激的條件下反應沖突監控需要消耗個體較多的注意資源,覺察反應沖突的速度比中性刺激條件更慢。負性情緒事件具有重要的生存適應價值,人腦對該類事件存在加工偏向。近期國內的一些研究也表明,隨著效價強度的增強,負性情緒對任務相關維度加工的影響也增大。本研究結果顯示,高、低喚醒度條件誘發的N2的波幅沒有差異。這表明,對于不同喚醒的負性刺激,早期的沖突監控過程沒有差異,根據本研究的結果,我們可以認為,人類對于高和低喚醒的負性刺激的抑制控制過程早期投入的注意資源是沒有差異的。
對行為抑制控制而言,行為抑制條件下中央頂區P3活動的出現是抑制控制加工的直接指標,行為抑制條件誘發的波幅顯著大于無控制條件。而且P3被認為是抑制過程的晚期階段的重要指標。事實上,本研究中很多峰值在刺激呈現后200~300ms之間的ERP成分都有表現。我們的研究結果是,在前額葉的FZ點,情緒刺激的P3波幅比中性刺激的波幅小,偏差刺激相比標準刺激引發較小的P3波幅,也就是說情緒刺激比中性刺激耗費了較少的腦資源。進一步對高、低喚醒刺激行為抑制誘發的P3波幅分析表明,高喚醒條件下的P3波幅相比低喚醒條件下的大,說明在行為控制的晚期,被試在對高喚醒的刺激進行行為抑制時,需要付出更多的資源,高喚醒度的負性情緒刺激更難進行行為抑制。如前所述,人腦對負性事件具有優先加工性。以往的研究顯示,情緒刺激影響注意,并易化了接下來的加工。因此高喚醒刺激更多的激活相關腦區,可能影響接下來的行為抑制過程,這些都會導致對高喚醒負性刺激比低喚醒刺激更難抑制控制。
5 結論
本研究采用雙選擇oddball范式,發現高喚醒度視覺情緒對行為抑制控制的效果具有顯著影響。該影響主要表現為對高喚醒度的負性情緒存在更難抑制控制的執行效果。對于行為抑制而言,高喚醒度負性刺激出現后,在抑制控制的早期和低喚醒刺激沒有差異,這個時候兩種喚醒的N2波幅沒有差異。而在行為抑制的晚期,對高喚醒度的負性刺激抑制控制引發的P3波幅明顯大于對低喚醒負性刺激引發的P3波幅,這說明,對高喚醒負性刺激的抑制控制,被試要付出更大的努力,因此更難抑制控制。本研究結果表明,對高喚醒負性刺激的行為抑制更難。參考文獻
1 Aron A R,Robbins T W,Poldrack R A.Inhibition and the right inferior frontal cortex.Trends in Cognitive Sciences,2004,8(4):170-177.
2 T?rnquist K,Malm A,Pasternack M,et al.Tumor necrosis factor-α,sphingomyelinase,and ceramide inhibit store-operated calcium entry in thyroid FRTL-5 cells.JournalofBiologicalChemistry, 1999, 274(14):9370-9377.
3 Dillon D G, PizzagalliD A.Inhibition ofaction,thought,and emotion:A selective neurobiological review.Applied and Preventive Psychology,2007,12(3):99-114.
4 Mostofsky S H,Simmonds D J.Response inhibitionand response selection:Two sides of the same coin.JournalofCognitive Neuroscience, 2008, 20(5):751-761.
5 Buchsbaum B R,Greer S,Chang W L,et al.Metaanalysis ofneuroimaging studies ofthe Wisconsin Card-Sorting task and component processes.Human Brain Mapping,2005,25(1):35-45.
6 Petrides M,Alivisators B,Freys.Differential activation of the human orbital,mid-ventrolateral,and mid-dorsolateral prefrontal cortex during the processing of visual stimuli.Proceedings of the National Academy of Sciences,2002,99(8):5649-5654.
7 Aron A R,Robbins T W,Poldrack R A.Inhibition and the right inferior frontal cortex.Trends in Cognitive Sciences,2004,8(4):170-177.
8 Bekker E M,Kenemans J L,Verbaten M N.Source analysis of the N2 in a cued Go/NoGo task.Cognitive Brain Research,2005,22(2):221-231.
9 Garavan H,Ross T J,Murphy K,et al.Dissociable executive functions in the dynamic control of behavior:Inhibition,error detection,and correction.Neuroimage,2002,17(4):1820-1829.
10 Rubia K,Russell T,Bullmore E T,et al.An fMRI study of reduced left prefrontal activation in schizophrenia during normal inhibitory function.Schizophrenia Research,2001,52(1):47-55.
11 Falkenstein M,Hoormann J,Hohnsbein J.ERP components in Go/Nogo tasks and their relation to inhibition.Acta Psychologica,1999,101(2):267-291.
12 Rowe G,Hirsh J B,Anderson A K.Positive affect increases the breadth of attentional selection.Proceedings of the National Academy of Sciences,2007,104(1):383-388.
13 Tsetsenis T,Ma X,Iacono L L,et al.Suppression of conditioning to ambiguous cues by pharmacogenetic inhibition ofthe dentategyrus.NatureNeuroscience,2007,10(7):896-902.
14 Shafritz K M,Collins S H,Blumberg H P.The interaction of emotional and cognitive neural systems in emotionally guided response inhibition.Neuroimage,2006,31(1):468-475.
15 Scheier M F,Carver C S,Bridges M W.Distinguishing optimism from neuroticism(and trait anxiety,selfmastery,and self-esteem):A reevaluation of the Life Orientation Test.Journal of Personality and Social Psychology,1994,67(6):1063.
16 Posner M I,Rothbart M K,Vizueta N,et al.Attentional mechanisms of borderline personalitydisorder. Proceedingsofthe NationalAcademy ofSciences,2002,99(25):16366-16370.
17 Silbersweig D,Clarkin J F,Goldstein M,et al.Failure of frontolimbic inhibitory function in the context of negative emotion in borderline personality disorder.American JournalofPsychiatry, 2007, 164(12):1832-1841.
18 Shachaf C M,Perez O D,Youssef S,et al.Inhibition of HMGcoA reductase by atorvastatin prevents and reverses MYC-induced lymphomagenesis.Blood, 2007,110(7):2674-2684.
19 Shafritz K M,Collins S H,Blumberg H P.The interaction of emotional and cognitive neural systems in emotionally guided response inhibition.Neuroimage,2006,31(1):468-475.
20 Stadler A,Schima W,Ba-Ssalamah A,et al.Artifacts in body MR imaging:Their appearance and how to eliminate them.European Radiology,2007,17(5):1242-1255.
21 Goldstein M,Brendel G,Tuescher O,et al.Neural substrates of the interaction of emotional stimulus processing and motorinhibitory control: An emotional linguistic go/no-go fMRI study.Neuroimage,2007,36(3):1026-1040.
22 Shafritz K M,Collins S H,Blumberg H P.The interaction of emotional and cognitive neural systems in emotionally guided response inhibition.Neuroimage,2006,31(1):468-475.
23 Schulz E,Maurer U,van der Mark S,et al.Impaired semantic processing during sentence reading in children with dyslexia:Combined fMRI and ERP evidence.Neuroimage,2008,41(1):153-168.
24 Nunez J M,Casey B J,Egner T,et al.Intentional false responding shares neural substrates with response conflict and cognitive control.Neuroimage,2005,25(1):267-277.
25 Chiu P H,Holmes A J,Pizzagalli D A.Dissociable recruitmentofrostralanteriorcingulate and inferior frontal cortex in emotional response inhibition.Neuroimage,2008,42(2):988-997.
26 Hare T A,Tottenham N,Davidson M C,et al.Contributions of amygdala and striatal activity in emotion regulation.Biological Psychiatry,2005,57(6):624-632.
27 Hare T A,Tottenham N,Galvan A,et al.Biological substratesofemotionalreactivity and regulation in adolescence during an emotional go-nogo task.Biological Psychiatry,2008,63(10):927-934.
28 Hare T A,Tottenham N,Galvan A,et al.Biological substratesofemotionalreactivity and regulation in adolescence during an emotional go-nogo task.Biological Psychiatry,2008,63(10):927-934.
29 Lynch J C,Ferreira L A S,Welling L,et al.Multiple intracranial meningiomas:Diagnosis,biological behavior and treatment.Arquivos de Neuro-psiquiatria,2008,66(3B):702-707.
30 Müller J L,Sommer M,Dhnel K,et al.Disturbed prefrontal and temporal brain function during emotion and cognition interaction in criminal psychopathy.Behavioral Sciences&the Law,2008,26(1):131-150.
31 Shafritz K M,Collins S H,Blumberg H P.The interaction of emotional and cognitive neural systems in emotionally guided response inhibition.Neuroimage,2006,31(1):468-475.
32 Aron A R.The neural basis of inhibition in cognitive control.The Neuroscientist,2007,13(3):214-228.
33 Dolan R J.The human amygdala and orbital prefrontal cortex in behavioural regulation.Philosophical Transactions of the Royal Society of London B:Biological Sciences,2007,362(1481):787-799.
34 Elliott R,Rubinsztein J S,Sahakian B J,et al.Selective attention to emotional stimuli in a verbal go/no-go task:An fMRI study.Neuroreport,2000,11(8):1739-1744.
35 Garavan H,Ross T J,Murphy K,et al.Dissociable executive functions in the dynamic control of behavior:Inhibition,error detection,and correction.Neuroimage,2002,17(4):1820-1829.
36 Rubia K,Russell T,Bullmore E T,et al.An fMRI study of reduced left prefrontal activation in schizophrenia during normal inhibitory function.Schizophrenia Research,2001,52(1):47-55.
37 Nagy E,Potts G F,Loveland K A.Sex-related ERP differences in deviance detection.International Journal of Psychophysiology,2003,48(3):285-292.
38 Roche R A,Garavan H,Foxe J J,et al.Individual differences discriminate event-related potentials but not performance during response inhibition.Experimental Brain Research,2005,160(1):60-70.
39 Bokura H,Yamaguchi S,Kobayashi S.Electrophysiological correlates for response inhibition in a Go/NoGo task.Clinical Neurophysiology, 2001, 112(12):2224-2232.
The Behavior Inhibition Characteristics of Different Arousal Negative Stimulation
Zhao Shaochen1,Zheng Xifu2
(1 Public Security Frontier Defense Forces College,Guangzhou 510663;2 College of Psychology,South China Normal University,Guangzhou 510631)
This research discussed the behavior inhibit characteristics of different arousal negative stimulations with the event related potential technology and classical oddball paradigm.This study selected 15 undergraduate and graduate students as the subjects.They were asked to press the key as fast and accurately as possible according to different stimulations(high sensei stimulation,low sensei stimulation and standard stimulus)and corresponding brain wave of these different types of incentives were recorded.The results showed that there was a significant difference among different arousal negative stimulations.In 210~300ms time interval,average amplitude was prominently different as well as latency;In 380~430ms time interval,there was a significant difference in average amplitude,too;It was found from the analysis of behavior inhibition process that the average amplitude of N2 and P3 wave exist significant differences in average volatility.These results reflect that there is a notable difference in behavior inhibition of different arousal negative stimulation.In the condition of high arousal stimulation,it is individual reaction monitoring conflicts and subsequent brain activity of behavior inhibit that were enhanced.The high arousal negative emotion has an obvious facilitated effect to behavior inhibition.
arousal degree;behavior inhibition;oddball;ERP
鄭希付,男,教授,博士生導師。Email:zhengxf@scnu.edu.cn
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
核科學與工程(2021年4期)2022-01-12 06:30:26
今日農業(2020年19期)2020-12-14 14:16:52
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
風流一代·青春(2018年2期)2018-02-26 15:27:06
風流一代·青春(2017年6期)2018-02-14 19:28:55
風流一代·青春(2017年5期)2018-02-14 09:32:37
中學物理·高中(2016年12期)2017-04-22 11:53:03
商業評論(2014年6期)2015-02-28 04:44:25
