節水灌溉稻田雜草群落多樣性分析
2017-03-22 05:15:13付浩龍羅玉峰熊玉江
中國農村水利水電 2017年2期
關鍵詞:雜草
付浩龍,羅玉峰,余 琪,熊玉江,喬 偉
(1.長江科學院農業水利研究所,武漢 430010;2.河海大學水文水資源與水利工程科學國家重點實驗室,南京 210098;3.武漢大學水資源與水電工程科學國家重點實驗室,武漢 430072;4.長江水利委員會人事勞動局,武漢 430015 )
雜草作為農田生態系統中的重要組成之一,與作物間存在對光照、水分和生長空間資源的競爭。據統計,在我國農田雜草危害面積高達0.43 億hm2,由雜草危害所帶來的農作物減產占10%以上[1,2]。因此,進行合理的農田管理,降低雜草危害,對保障作物良好生長具有重要意義。
隨著雜草危害認識的不斷深入,學者對稻田雜草群落也進行了大量研究,但研究多集中在其危害性和防治途徑方面[3,4],且在施肥模式、種植制度和耕作方式等管理措施上來開展雜草多樣性研究[5-7]。然而,隨著節水灌溉技術的大力推廣,作為人為重要管理措施的節水灌溉下稻田雜草群落多樣性研究卻相對較少,且以往的研究也多只反映在田間雜草的種類及發生程度上,沒有對雜草危害度優勢種、均勻度和多樣性指數等進行深入的探討。因此,本文在高郵灌區開展控制灌溉和常規灌溉雜草群落多樣性調查,在明確雜草群落結構組成的基礎上開展稻田雜草多樣性研究,以期為節水灌溉稻田雜草控制提供科學的依據。
1 材料與方法
1.1 試驗區概況
本試驗于2013年6-10月在江蘇省高郵灌區試驗田進行。該灌區內土壤肥沃,氣候溫和,陽光充足、雨量充沛,年平均氣溫在14.8 ℃左右,年平均降水量為1 000 mm,平均風速基本保持在3.5 m/s左右,適合水稻的生長。灌區水稻作物種植面積達6 300 hm2,有效灌溉面積達5 900 hm2,罐區實行稻麥輪作,同時該灌區正在推廣控制灌溉技術,能夠很好地保障試驗的開展。
1.2 試驗處理與方法
試驗當中節水灌溉實行控制灌溉,以當地常規灌溉為對照。共設控制灌溉和常規灌溉2個處理,每個處理2個重復,每塊稻田種植面積約為0.233 hm2,水稻生長過程中為了防止除草劑對植物生長的影響,保持基本相同的農藝措施 (見表1),水稻品種為當地普遍種植的鎮稻99。
本試驗調查方法以典型水稻田塊為依托,在踏查的基礎上,對不同灌溉模式水稻逐生育期內隨機抽取5個代表性樣方(為避免非稻田生鏡對雜草群落的影響,每次樣點的選擇應該避開田埂附近,至少保持0.8 m的距離),樣方大小為1 m×1 m,采用7級目測法及分類標準[8],在每個樣方內先目測出總的雜草危害級別,然后逐一地記錄每個樣方中雜草種類、高度及其密度,并計算出雜草出現頻率(雜草種類以《中國雜草志》的種類鑒定為依據)。雜草與水稻的株高采用鋼尺直接進行測量,雜草覆蓋度通過數碼相機拍攝獲取,并在地理信息系統軟件中通過數字化后,計算面積比求得。
從分蘗前期(返青期群落剛剛遷入且田間水層較高雜草植株生長還未成熟故不考慮)開始,對常規灌溉典型田塊和控制灌溉典型田塊水稻各生育期內雜草植株群落種類、數量進行調查,各生育期調查時間見表2。

表1 農藝措施(農藥、除草劑)情況Fig.1 The table of agronomic measures ( pesticides, herbicides)
注:①單位為mL/hm2。
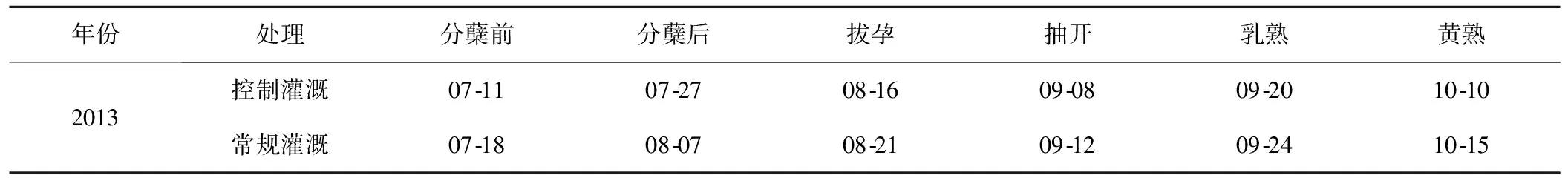
表2 不同灌溉模式下稻田各生育期雜草植株群落調查時間Fig.2 The research time of the weed communities with each growth period of rice under different irrigation modes
1.3 試驗數據分析
通過調查記錄各雜草的種類、高度、蓋度及其水稻植株高度,并計算出相應的重要值,運用物種豐富度S、Shannon-Wiener多樣性指數和Pielou均勻度指數來對雜草群落多樣性進行分析[9,10]。同時調查每平米內各生育期雜草的數量即雜草分布密度,并把該雜草密度作為反映雜草植株發生程度的重要因子。雜草優勢種的界定則是以雜草相對分布密度的10%為基準,當某種雜草密度占所有雜草密度的百分比(相對密度)有3次大于10%則定位優勢種,小于等于10%的雜草則定位為非優勢種。其計算公式如下:
雜草重要值IV=相對多度+相對高度+相對蓋度
豐富度S:
S=群落物種個體數
Shannon-Wiener多樣性指數H′:
(1)
Pi=ni/N
式中:Pi為第i種個體占群落個體總數的比例;ni為群落中第i個物種的個體總數;N為群落個體總數量。
Pielou均勻度指數E:
(2)
Hmax=lnS
式中:Hmax為物種當中最大的多樣性指數。
同時,本試驗數據分析和處理均采用EXCEL軟件,計算物種多樣性指數、均勻度指數及雜草密度等,用LSD進行不同處理下雜草群落各指標顯著性差異分析(p=0.05)[11,12]。
2 結果與分析
2.1 稻田水層及土壤含水率變化
本試驗實行控制灌溉和常規灌溉2種灌溉模式。其中常規灌溉按當地習慣進行淺濕調控灌溉,分別于水稻泡田栽插期、分蘗期和曬田以后,每隔4、5 和6 d灌水一次,每次灌15~20 h,自流灌溉至田間有4~8 cm薄水層;而控制灌溉是指秧苗本田移栽后,田面保留5~25 mm薄水層返青,此后各生育階段灌水后田面不建立水層,以根層土壤水分為控制指標,確定灌水時間和灌水定額。
不同灌溉模式A、B田塊水層與土壤含水率分布見圖1。由圖1可知,控制田塊土壤含水率明顯要高于常規灌溉,常規灌溉田塊大部分時間處于有水層狀態。由此可見,不同灌溉模式稻田水環境顯著不同,從而影響田間雜草種類及群落特點。

圖1 不同灌溉模式田間水層和土壤含水率變化情況Tab.1 The changes of water layer and soil moisture with different irrigation modes in field
2.2 稻田雜草群落組成及發生密度
通過對不同灌溉模式下稻田間雜草群落進行調查,其調查結果見表3。由表3可知,控制灌溉稻田雜草種類共有10科13種,常規灌溉則有9科11種,控制灌溉雜草種類較常規灌溉多;其中水虱草(Fimbristylis miliaceae)、合萌(Aeschynomene indica)及鯉腸(EcIipta prostrata L.)為控制灌溉稻田特有雜草,紫萍(Spirodela)則為常規灌溉特有雜草。不難看出,由于控制灌溉稻田田間均不建立水層(除返青期),這種控制水層灌溉方式比起長期淹灌的常規灌溉而言給予了雜草生長更多的環境選擇,促進了部分喜濕性旱地雜草的生長,結果導致控制灌溉稻田間雜草植株科、種要多于常規灌溉。而常規灌溉田間作為水層長期存在,使的田間環境單一,野荸薺(Heleocharis plantagineiformis)、紫萍(Spirodela)和輪藻(Chara)等典型喜濕性水生植株得到有利生長。
圖2則反映了不同灌溉模式下稻田雜草群落總密度分布情況。由圖2可知,控制灌溉和常規灌溉稻田雜草密度在時間上存在幾乎相同的動態變化,雜草密度隨生育期都呈現先增加后減少的變化趨勢。其中,控制灌溉雜草密度最大出現在拔節孕穗期為37.8 莖/m2,最小則出現在分蘗前期為16.8 莖/m2;而常規灌溉雜草密度最大為60.0 莖/m2,最小為22 莖/m2,分別出現在分蘗后期和分蘗前期。且在分蘗后期和抽穗開花期控制灌溉和常規灌溉稻田雜草密度分布呈現差異顯著(F2,18=11.99,p=0.002 778;F2,18=4.84,p=0.041 098),生育后期(乳熟期和黃熟期)控制灌溉雜草密度略大于常規灌溉但差異不顯著(F2,18=0.18,p=0.674 315;F2,18=0.004 9,p=0.944 751),分蘗前期和拔節孕穗期控制灌溉雜草密度低于常規灌溉且差異也同樣不顯著(F2,18=0.85,p=0.368 135;F2,18=1.45,p=0.244 511)。
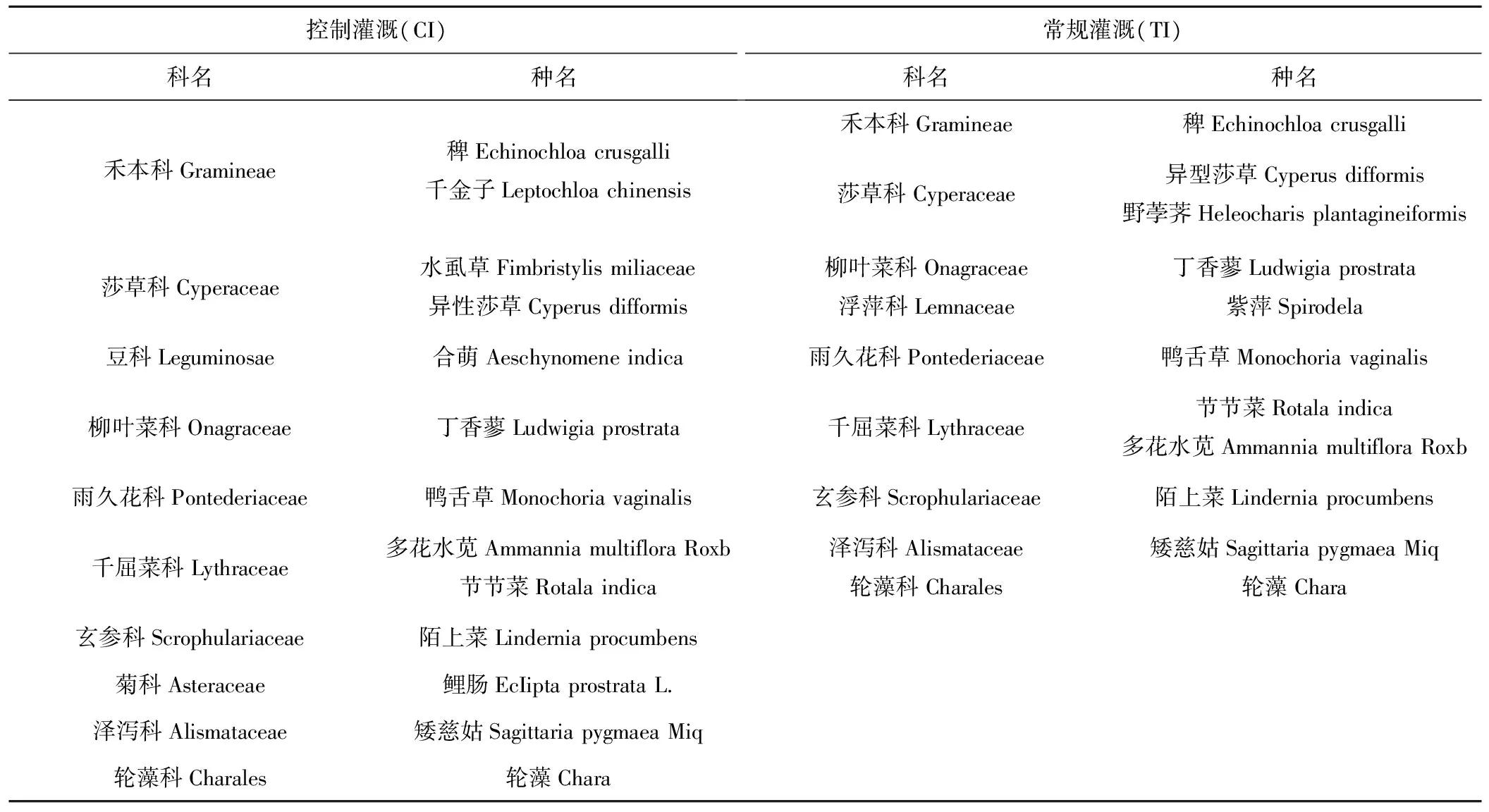
表3 不同灌溉模式田間雜草種類Fig.3 The field weed species with different irrigation modes

圖2 不同灌溉模式稻田雜草群落總密度Tab.2 The density of paddy weed community with different irrigation modes
因此,可以看出常規灌溉稻田雜草各生育期密度總體是要大于控制灌溉,除生育前期和生育后期控制灌溉雜草密度略大于常規灌溉,其余各生育期尤其是中期控制灌溉稻田雜草密度均小于常規灌溉,且差異顯著。
控制灌溉和常規灌溉稻田各生育期雜草群落的相對密度分布情況見表4。調查結果顯示,控制灌溉雜草優勢種為稗(Echinochloa crusgalli)、丁香蓼(Ludwigia prostrata)、鴨舌草(Monochoria vaginalis)和陌上菜(Lindernia procumbens),且其相對密度分別為10.0%~26.9%、10.7%~28.2%、10.2%~14.3%和14.3%~53.3%。而常規灌溉雜草優勢種則為丁香蓼(Ludwigia prostrata)、鴨舌草(Monochoria vaginalis)、陌上菜(Lindernia procumbens)、輪藻(Chara)和紫萍(Spirodela),且相對密度分別為12.7%~50.9%、10.0%~25.3%、11.0%~42.7%、10%~22.1%和14.1%~40.1%。控制灌溉稻田雜草優勢種要少于常規灌溉。
從雜草密度隨生育期時間動態變化來看,作為控制灌溉特有優勢種稗(Echinochloa crusgalli)的密度在生育后期呈現下降的變化趨勢,而作為常規灌溉特有優勢種的紫萍(Spirodela)和輪藻(Chara)則在整個生育期內相對密度均處于較高,相對穩定,變化趨勢不明顯。同時作為控制灌溉和常規灌溉共有的優勢種丁香蓼(Ludwigia prostrata)、鴨舌草(Monochoria vaginalis)和陌上菜(Lindernia procumbens),其控制灌溉田間雜草密度要低于常規灌溉,說明控制灌溉能夠有效地抑制稻田優勢種雜草的生長,降低雜草密度。

表4 不同灌溉模式稻田各生育期雜草群落相對密度 %
注:各生育期內3次以上的相對密度>10%的為優勢種。
2.3 稻田雜草群落多樣性分析
均勻度指數反映了群落分布均勻情況,而多樣性指數能夠體現出群落的穩定性。不同灌溉模式下,稻田各生育期雜草群落均勻度分布情況見圖3。由圖3可知,控制灌溉和常規灌溉雜草群落均勻度整體上在各生育期內變化幅度不大,在生育前期控制灌溉稻田雜草群落與常規灌溉幾乎相同甚至低于常規灌溉,但隨著生育期的推進在水稻生育后期控制灌溉雜草群落均勻度要略大于常規灌溉,雜草群落穩定性得到提高。控制灌溉稻田雜草群落均勻度指數最大出現在拔節孕穗期,最小則出現在分蘗末期,常規灌溉雜草均勻度指數最大在分蘗前期,最小則在分蘗末期。從差異顯著性分析來看,除了拔節孕穗期和抽穗開花期差異顯著外(F2,18=10.18,p=0.005 058;F2,18=5.35,p=0.032 698),其余各生育期差異均不顯著。
在拔節孕穗期我們看到2種不同灌溉模式雜草群落均勻度出現反差,且差異呈現出顯著性,這可能與控制灌溉田間水環境的改變及其拔節孕穗期間出現的曬田有關。控制灌溉無水層環境較常規灌溉淹水環境更接近于曬田期間環境,這樣迫使一些對環境改變適應性差的雜草植株死亡,導致常規灌溉田間其他適應強的雜草有足夠的生長空間和資源,打破了原有的雜草群落分布均勻的特征,從而降低了常規灌溉雜草群落均勻度。

圖3 不同灌溉模式稻田雜草群落均勻度Tab.3 The values of evenness of paddy weed community with different irrigation modes
圖4則反映了不同灌溉模式下稻田各生育期雜草群落多樣性指數分布情況。由圖4可知,控制灌溉和常規灌溉稻田雜草群落多樣性指數隨生育期出現相同的趨勢走向,均呈現先增加在減少的變化。在生育前期常規灌溉雜草群落多樣性指數略大于控制灌溉,但到生育后期控制灌溉雜草群落多樣性指數則又略大于常規灌溉。從差異顯著性分析來看,除了拔節孕穗期差異顯著外(F2,18=6.42,p=0.020 808),其余各生育期差異均不顯著。拔節孕穗期多樣性指數差異顯著很可能與此階段均勻度指數變化有關。

圖4 不同灌溉模式稻田雜草群落多樣性指數Tab.4 The values of diversity index of paddy weed community with different irrigation modes
3 結 論
本文通過對不同灌溉模式下稻田雜草群落進行調查及多樣性分析,得到如下結論。
(1)控制灌溉稻田雜草組成及豐富度要多于常規灌溉,控制灌溉共有10科13種,常規灌溉則有9科11種。
(2)控制灌溉田間雜草密度總體要低于常規灌溉,且隨生育期推進控制灌溉稻田優勢種雜草密度呈現下降趨勢,控制灌溉能夠有效地抑制稻田優勢種雜草的生長,降低雜草發生密度。
(3)不同灌溉模式下稻田雜草Shannon-Wiener多樣性指數各生育期內變化不大,且隨生育期變化趨勢基本相同,但控制灌溉稻田雜草Pielou均勻度在生育后期整體大于常規灌溉,雜草群落趨于相對穩定。
□
[1] 張朝賢. 農田雜草與防控[M]. 北京:中國農業出版社, 2011.
[2] 牛新勝, 劉美菊, 張宏彥,等. 不同耕作、秸稈及氮素管理措施對冬小麥-夏玉米輪作田雜草生物量影響的研究[J]. 中國土壤與肥料, 2011,(6): 49-53.
[3] 戴偉民, 宋小玲, 吳 川,等. 江蘇省雜草稻危害情況的調研[J]. 江蘇農業學報, 2009,25(3):712-714.
[4] 王 強, 何錦豪, 李妙壽,等. 浙江省水稻田雜草發生種類及危害[J]. 浙江農業科學,2000,12(6):317-324.
[5] 蔣 敏, 沈明星, 沈新平,等. 長期不同施肥方式對麥田雜草群落的影響[J]. 生態學報, 2014,34(7):1 746-1 756.
[6] 沈建凱, 黃 璜, 傅志強,等. 規模化稻鴨生態種養對稻田雜草群落組成及物種多樣性的影響[J]. 中國生態農業學報, 2010,18(1):123-128.
[7] 楊濱娟, 黃國勤, 徐 寧,等. 長期水旱輪作條件下不同復種方式對稻田雜草群落的影響[J]. 應用生態學報2013,24(9):2 533-2 538.
[8] 強 勝. 中國農田雜草多樣性及其危害發生規律的研究[D]. 南京:南京農業大學,2001.
[9] Derksen D, Thomas A, Lafond G, et al. Impact of post-emergence herbicides on weed community diversity within conservation-tillage systems[J]. Weed Research, 1995,35(4):311-320.
[10] Stevenson FC, Légère A, Simard RR, et al. Weed species diversity in spring barley varies with crop rotation and tillage, but not with nutrient source[J]. Weed Science, 1997,45(4):798-806.
[11] 劉雨芳. EXCEl在群落生物多樣性參數計算中的應用[J]. 湘潭師范學院學報(自然科學版), 2003,(2):80-82.
[12] Barberi P, Silvestri N, Bonari E. Weed communities of winter wheat as influenced by input level and rotation[J]. Weed Research, 1997,37(5):301-313.
猜你喜歡
科教新報(2022年22期)2022-07-02 12:34:28
今日農業(2021年5期)2021-11-27 17:22:19
當代作家(2021年12期)2021-01-15 21:39:15
高中生學習·閱讀與寫作(2019年2期)2019-09-10 07:22:44
現代園藝(2017年11期)2017-06-28 11:32:57
新農業(2016年19期)2016-08-16 12:00:18
雜草學報(2015年2期)2016-01-04 14:58:05
種業導刊(2016年9期)2016-01-03 01:27:11
現代農業(2015年5期)2015-02-28 18:40:49
雜草學報(2012年1期)2012-11-06 07:08:33
