去甲斑蝥素作用早期對人肝癌細胞活性氧及NF-E2相關因子2/抗氧化反應元件信號通路的激活
2017-03-28 11:18:33崔寶弟孫震曉
中國藥理學與毒理學雜志 2017年1期
陳 靜,崔寶弟,孫震曉
(北京中醫藥大學中藥學院,北京 100102)
去甲斑蝥素作用早期對人肝癌細胞活性氧及NF-E2相關因子2/抗氧化反應元件信號通路的激活
陳 靜*,崔寶弟*,孫震曉
(北京中醫藥大學中藥學院,北京 100102)
目的探究去甲斑螯素(NCTD)誘導人肝癌HepG2細胞凋亡及G2/M期阻滯的早期事件,分析NCTD作用早期HepG2細胞內活性氧自由基(ROS)的變化規律及NCTD對NF-E2相關因子/抗氧化反應元件(Nrf2/ARE)信號通路的影響。方法NCTD30,60和120 μmol·L-1分別作用于體外培養的人肝癌HepG2細胞3,6,12,24,48和72 h,MTT法檢測NCTD對細胞存活的影響;流式細胞術檢測NCTD 60 μmol·L-1作用細胞12,24和48 h對細胞周期和細胞凋亡的影響;DCFH-DA探針結合流式細胞術檢測NCTD 30,60和120 μmol·L-1作用于3,6和12 h對HepG2細胞內ROS的影響;熒光素酶法測定NCTD同時轉染ARE和熒光素酶報告基因的HepG2C8細胞的熒光強度;實時熒光定量PCR檢測對血紅素氧合酶-1(HO-1)和醌氧化還原酶-1(NQO1)mRNA表達的影響。結果NCTD 30,60和120 μmol·L-1作用3和6 h對HepG2細胞存活無明顯影響,而作用24,48和72 h對HepG2細胞有明顯生長抑制作用(P<0.01);NCTD 60 μmol·L-1作用12 h后可誘導HepG2細胞發生凋亡及G2/M期阻滯,12,24和48 h凋亡細胞比例分別由12 h細胞對照組(4.00±1.98)%增加到(12.10±1.70)%,24 h對照組(4.05±0.21)%增加到(31.80± 6.50)%,48 h對照組(3.90±0.85)%增加到(33.30±1.41)%;12,24和48 h G2/M期細胞比例分別由12 h對照組的(16.51±1.58)%增加到(40.89±0.18)%,24 h對照組的(16.99±1.32)%增加到(55.29±3.99)%,48 h對照組的(14.45±0.59)%增加到(50.66±5.88)%,相應各時相NCTD處理組G1期細胞比例明顯下降(P<0.01);NCTD 30,60和120 μmol·L-1作用HepG2細胞3,6和12 h,ROS無明顯變化,作用HepG2C8細胞6和12 h可明顯激活Nrf2/ARE信號通路,下游基因HO-1和NQO1 mRNA表達顯著上調(P<0.05)。結論NCTD作用HepG2細胞早期可明顯激活Nrf2/ARE信號通路;ROS激活可能不是NCTD誘導人肝癌HepG2細胞凋亡及G2/M期阻滯的主要原因。
去甲斑蝥素;細胞凋亡;細胞周期;Nrf2/ARE信號通路;活性氧自由基;血紅素加氧酶-1;NAD(P)H:醌氧化還原酶-1
去甲斑蝥素(norcantharidin,NCTD)是我國自主研發的一種新型抗腫瘤藥物,兼有抗腫瘤和升高外周血白細胞的作用,近年來主要用于消化道腫瘤和肺癌的治療,其中對肝癌的治療效果最好[1],也可與其他化療藥物聯用來提高療效、減少副作用[2]。盡管有關NCTD抗癌活性研究已經有不少報道[3-8],但對其細胞毒作用的早期事件仍缺乏研究。
研究表明,高濃度活性氧(reactive oxygen species,ROS)或低濃度ROS長時間作用于細胞,均可導致細胞周期阻滯[9]或細胞凋亡[10],而低濃度ROS短期作用可以引起細胞產生氧化應激,激活NF-E2相關因子2/抗氧化反應元件(NF-E2-related factor 2/ antioxidant response element,Nrf2/ARE)信號通路[11],Nrf2/ARE信號通路在機體抵抗內外氧化和化學刺激等方面均發揮重要作用。本研究主要在NCTD體外作用人肝癌HepG2細胞的時效、量效關系基礎上,進一步探索NCTD對細胞內ROS及細胞氧化應激信號通路Nrf2/ARE的影響,進一步揭示NCTD的抗癌機制。
1 材料與方法
1.1 試劑和藥品
1640培養基,胎牛血清,MEM非必需氨基酸購自美國Gibco公司;Trizol Reagent購自美國Invitrogen公司;無RNA酶水購自美國AMERESCO公司;PrimeScriptTM1ststrand cDNA Synthesis試劑盒購自日本TaKaRa公司;PCR引物購自生工生物工程(上海)股份有限公司;細胞凋亡檢測試劑盒購自南京凱基生物科技發展有限公司;RNA酶抑制劑購自美國Promega公司;DCFH-DA探針購自美國Sigma公司。
1.2 主要儀器
低溫高速臺式離心機購自美國Kendro儀器公司,小動物活體成像儀購自美國Caliper公司,熒光定量PCR(qRT-PCR)儀購自日本Bio-Rad公司,流式細胞儀購自美國Becton Dickinson公司,微量RNA濃度檢測儀購自美國Biotek公司。
1.3 細胞系
人肝癌HepG2細胞,北京中醫藥大學生物制藥實驗室凍存;HepG2C8細胞為轉染了抗氧化元件ARE和熒光素酶基因的HepG2細胞,由北京大學醫學部余四旺老師惠贈。
1.4 細胞培養
HepG2細胞用含10%(V/V)胎牛血清,1%雙抗(青霉素100 U·mL-1和鏈霉素100 μg·mL-1)的RPMI 1640培養基,在37℃,5%CO2的飽和濕度培養箱中培養,2~3 d換液傳代1次。取對數生長期細胞為實驗對象。HepG2C8細胞培養參照文獻[12-13],用含10%(V/V)胎牛血清,1%雙抗,1% MEM非必需氨基酸的RPMI 1640培養基,在37℃,5%CO2的飽和濕度細胞培養箱中培養,2~3 d換液傳代1次。取對數生長期細胞為實驗對象。
1.5 MTT法檢測細胞存活
將HepG2細胞以每孔2×103的密度鋪于96孔板中,培養24 h后用NCTD 30,60和120 μmol·L-1處理。每個濃度設置8個平行孔,細胞對照組的孔內加入正常培養基作為對照。分別于3,6,12,24,48和72 h加入終濃度為0.5 g·L-1的MTT繼續孵育4 h后,吸去孔內MTT。每孔加入150 μL DMSO,置微量振蕩器上振蕩10 min,在酶標儀570 nm波長處讀取吸光度值(A),并計算NCTD對HepG2細胞的增殖抑制率(%)。抑制率(%)=(1-加藥組A570nm/對照組A570nm)×100%。
1.6 流式細胞儀檢測細胞凋亡與周期
將HepG2細胞以每孔6×104的密度鋪于6孔板中,24 h后用NCTD 60 μmol·L-1處理12,24和48 h后,收集每個孔中的全細胞,每個時間點設置3個平行對照,實驗完全獨立得重復3次。細胞對照組細胞加入正常培養基作為對照。按說明書用流式細胞儀檢測細胞凋亡和周期變化。
1.7 細胞內ROS水平檢測
將HepG2細胞以每孔3×105的密度接種于6孔板中,24 h后實驗組換含濃度分別為NCTD30,60和120 μmol·L-1的完全培養基,細胞對照組細胞加入正常培養基作為對照。分別于3,6和12 h后收集每個孔中的全細胞,加入終濃度為100 μmol·L-1的DCFH-DA探針溶液黑暗孵育30 min后,洗去多余的探針,用流式細胞儀在488 nm激發波長,525 nm發射波下檢測熒光強度[14-15],熒光強度數值與ROS水平成正相關關系。每個時間點、每個濃度設置3個平行對照。
1.8 熒光素酶法檢測HepG2C8細胞熒光強度
將轉染ARE及熒光素酶報告基因的HepG2細胞HepG2C8細胞以每孔1×104的密度常規接種于96孔板中,24 h后實驗組換含濃度分別為NCTD 30,60和120 μmol·L-1的完全培養基,叔丁基對苯二酚(tert-butylhydroquinone,tBHQ)為Nrf2的激活劑,以tBHQ為陽性藥,分別在3,6和12 h時,在96孔板中添加100 μL用PBS稀釋的熒光素酶底物,立即用小動物成像儀檢測96孔板中每個孔的總熒光強度。
1.9 qRT-PCR技術檢測血紅素加氧合酶-1(heme oxygenase-1,HO-1)、NAD(P)H:醌氧化還原酶-1〔NAD(P)H:quinoneoxidoreductase 1,NQO1〕mRNA的表達
將HepG2細胞以每孔3×105的密度接種于6孔板中,24 h后實驗組換含濃度分別為NCTD 30,60和120 μmol·L-1的完全培養基,分別在3,6和12h后收集每個孔中全細胞,按照Trizol說明書提取RNA,合成cDNA;實時qRT-PCR的反應條件:95℃,10 min;95℃,15 s;60℃,1 min,共40個循環,以β肌動蛋白為內參,2-ΔΔCt法計算mRNA的相對表達水平。qRT-PCR引物見表1。
1.10 統計學分析
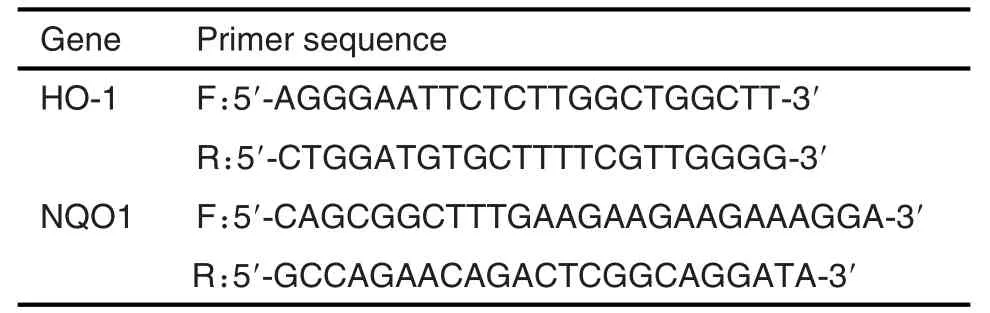
Tab.1 Prime sequence of qRT-PCR
2 結果
2.1 去甲斑蝥素對HepG2細胞存活的影響
NCTD 30,60和120 μmol·L-1在3和6 h內,NCTD 30和60 μmol·L-1在12 h內對HepG2細胞生長無明顯抑制作用;NCTD 120 μmol·L-1作用12 h、NCTD 30,60和120 μmol·L-1作用24,48和72 h對HepG2細胞生長有明顯抑制作用(P<0.01),抑制作用與用藥濃度及時間呈正比(圖1)。3,6,12,24,48和72 h的濃度效應相關系數分別為0.479,0.544,0.892(P<0.01),0.797(P<0.01),0.900(P<0.01)和0.941(P<0.01)。30,60和120 μmol·L-1的時間效應相關系數分別為0.829(P<0.01),0.899(P<0.01)和0.933(P<0.01)。
NCTD對HepG2細胞存活的影響發現,NCTD 30,60和120 μmol·L-1分別作用3,6和12 h,除120 μmol·L-1作用12 h對HepG2細胞生長有一定抑制作用外,其他作用濃度和時間對該細胞生長均無明顯抑制作用,故選擇NCTD 30,60和120 μmol·L-1作用3,6和12 h研究其對HepG2細胞內ROS含量的影響。
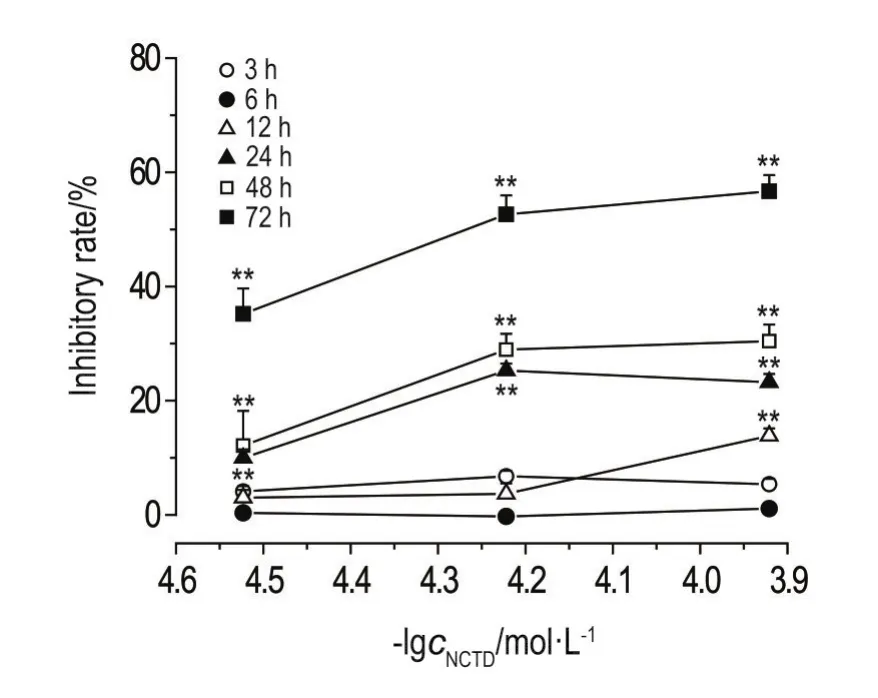
Fig.1 Inhibitory rate of norcantharidin(NCTD)on HepG2 cells for 3-72 h by MTT assay.,n=3.**P<0.01,compared with cell control group.The inhibitory rate of cell control was taken as 0%.
2.2 去甲斑蝥素對HepG2細胞凋亡的影響
流式結果(圖2)顯示,NCTD 60 μmol·L-1作用HepG2細胞12 h即可以檢測到明顯的細胞凋亡,12,24和48 h凋亡細胞比例分別由12 h由細胞對照組的(4.00±1.98)%增加到NCTD 60 μmol·L-1組的(12.10±1.70)%(P<0.05),24 h細胞對照組的(4.05±0.21)%增加到NCTD 60 μmol·L-1組的(31.80±6.50)%(P<0.01),48 h細胞對照組的(3.90±0.85)%增加到NCTD 60 μmol·L-1組的(33.30±1.41)%(P<0.01)。
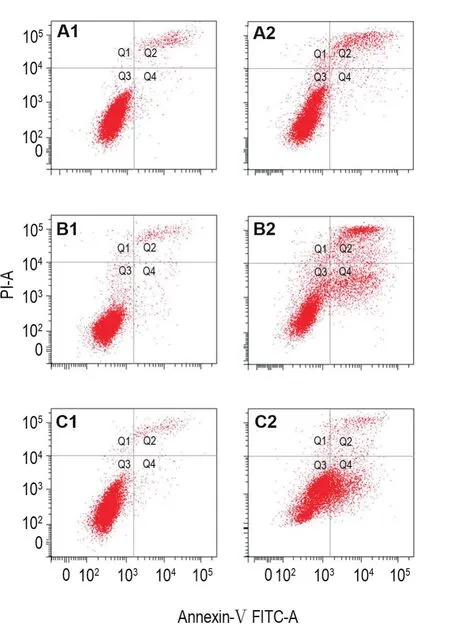
Fig.2Apoptosis of HepG2 cells induced by NCTD detected by flow cytometry analysis.A1:cell control group for 12 h;B1:cell control group for 24 h;C1:cell control group for 48 h;A2:NCTD 60 μmol·L-1for 12 h;B2:NCTD 60 μmol·L-1for 24 h;C2:NCTD 60 μmol·L-1for 48 h.
2.3 去甲斑蝥素對HepG2細胞周期的影響
如流式結果(圖3)所示,NCTD 60 μmol·L-1處理HepG2細胞12 h即誘導細胞發生明顯的G2/M期阻滯,由細胞對照組的(16.51±1.58)%增加到NCTD 60 μmol·L-1組的(40.89±0.18)%(P<0.01),24 h由細胞對照組的(16.99±1.32)%增加到NCTD 60 μmol·L-1組的(55.29±3.99)%(P<0.01),48 h由細胞對照組的(14.45±0.59)%增加到NCTD 60 μmol·L-1組的(50.66±5.88)%(P<0.01)。
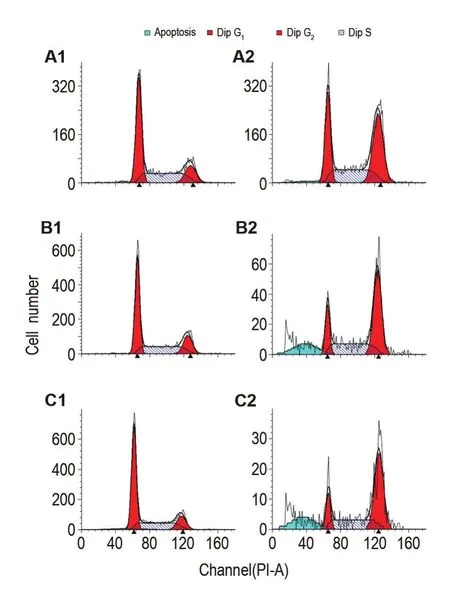
Fig.3 Cell cycle of HepG2 cells induced by NCTD. A1:cell control group for 12 h;B1:cell control group for 24 h;C1;cell control group for 48 h;A2:NCTD 60 μmol·L-1for 12 h;B2:NCTD 60 μmol·L-1for 24 h;C2:NCTD 60 μmol·L-1for 48 h.
2.4 去甲斑蝥素對HepG2細胞內ROS含量的影響
NCTD在所選濃度范圍內作用HepG2細胞3~12 h,對細胞ROS含量的影響無統計學意義(圖4)。
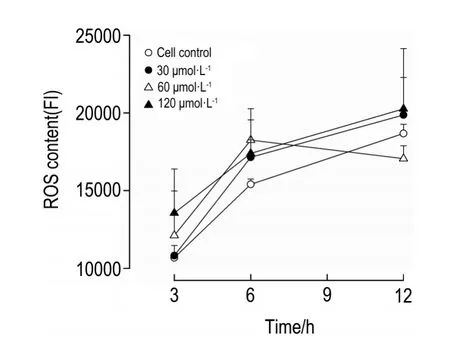
Fig.4 Effect of NCTD on reactive oxygen species(ROS)content in HepG2 cells.FI:fluorescence intensity.
2.5 去甲斑蝥素對HepG2C8細胞中Nrf2/ARE信號通路的激活作用
NCTD 30 μmol·L-1作用6和12 h對HepG2C8細胞中ARE均有顯著激活作用(P<0.05,P<0.01),NCTD 60和120 μmol·L-1在3,6和12 h內對HepG2C8細胞中ARE均有明顯激活作用(P< 0.05,P<0.01)。隨著NCTD濃度的增加,同一時間點激活作用逐漸增強。與細胞對照組相比,NCTD 60 μmol·L-1作用6 h對ARE的激活作用達到1.7倍,NCTD 120 μmol·L-1作用6和12 h對ARE的激活作用分別達到3.3和5.4倍(圖5)。
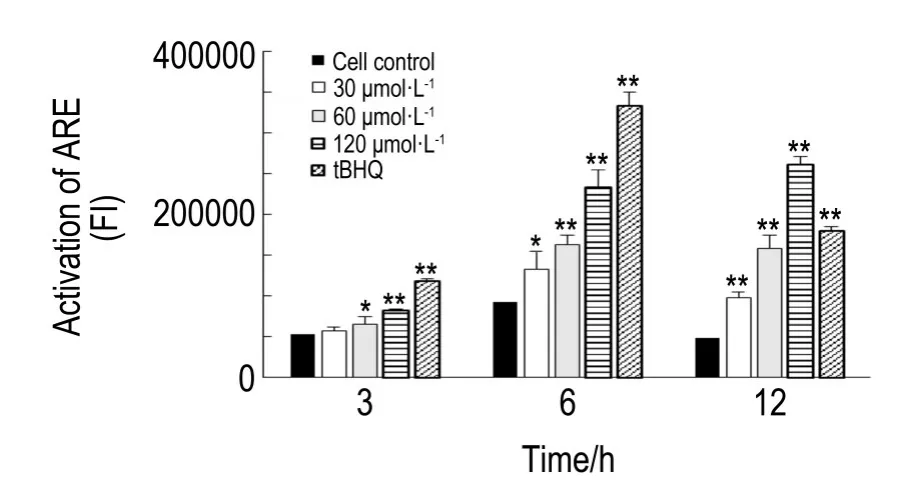
Fig.5 Effect of NCTD on activation of ARE in epG2C8 cells trasfected with ARE and luciferase report gene in HepG2 cells.tBHQ:tert-butylhydroquinone,positive control.n=3.*P<0.05,**P<0.01,compared with cell control group.
2.6 去甲斑蝥素對HO-1和NQO1 mRNA表達的影響
與細胞對照組比較,NCTD30,60和120μmol·L-1作用HepG2細胞3 h對HO-1和NQO1 mRNA表達無明顯影響,作用6 h,對HO-1和NQO1 mRNA表達均有明顯激活作用(對HO-1 mRNA表達激活分別為P<0.05,P<0.05,P<0.01;對NQO1 mRNA表達激活均為P<0.01),作用12 h仍保持明顯激活作用(對HO-1 mRNA表達激活分別為P<0.05,P<0.01,P<0.01;對NQO1 mRNA表達激活分別為P<0.05,P<0.05,P<0.01),HO-1mRNA表達與6 h基本持平(30 μmol·L-1)或顯著增強(60和120 μmol·L-1)(均P<0.01),而NQO1 mRNA的表達12 h比6 h明顯減弱(30 μmol·L-1)(P<0.01)(圖6)。
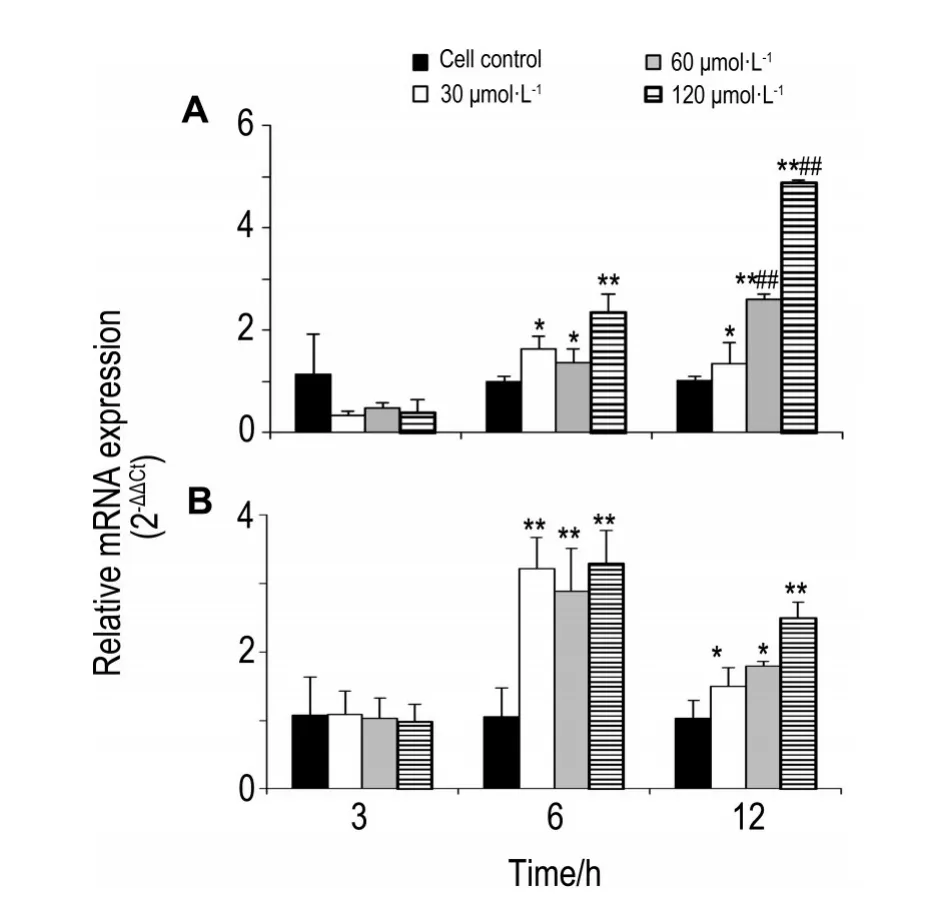
Fig.6 Effect of NCTD on mRNA expression of heme oxygenase-1(HO-1)(A)and NAD(P)H:quinoneoxido?reductase 1(NQO1)(B)in HepG2 by qRT-PCR.*P<0.05,**P<0.01,compared with cell control group;##P<0.01,compared with corresponding 6 h treatment group.
3 討論
本研究結果表明,NCTD在一定作用時間和濃度下對HepG2細胞存活有明顯抑制作用,且抑制效果與藥物作用時間和濃度成正相關。從NCTD對HepG2細胞凋亡和周期的影響可見,HepG2細胞在NCTD作用12 h后發生明顯的G2/M期阻滯及凋亡,因而探究發生在NCTD作用HepG2細胞12 h內的早期生物學事件對理解NCTD誘導細胞凋亡和G2/M期阻滯有重要價值。
曾有報道,NCTD可以促進腫瘤細胞ROS的產生并進一步引起細胞凋亡[19-20],但本研究通過流式細胞術檢測發現,NCTD 30~120 μmol·L-1作用3~12 h對HepG2細胞中ROS水平無明顯影響,推測這個階段ROS產生可能不是NCTD誘導人肝癌HepG2細胞凋亡及G2/M期阻滯的主要原因,NCTD對腫瘤細胞內ROS水平的影響可能受細胞系[19]及ROS分析方法的影響[20],目前研究數據雖不支持細胞內ROS激活與NCTD誘導細胞凋亡與周期阻滯有正相關性,但不排除NCTD作用腫瘤細胞更早期時產生的ROS已打開細胞凋亡或周期阻滯相關信號通路,二者的關系尚需進一步研究。我們在NCTD對其他腫瘤細胞的抑制作用中曾發現,NCTD對腫瘤細胞中微管的聚合有明顯的抑制作用[21-22],推測NCTD直接作用于細胞骨架等結構繼而引起細胞周期阻滯或凋亡在NCTD誘導HepG2細胞周期阻滯和凋亡中占主要地位。
本研究根據Nrf2/ARE信號通路激活機制,用轉染了ARE的HepG2C8細胞作為實驗工具,檢測NCTD對Nrf2/ARE信號通路的激活作用。結果顯示,在12 h內,NCTD 30,60和120 μmol·L-1有明顯激活Nrf2/ARE信號通路的作用,且激活效率有一定濃度依賴性。已知細胞內ROS可以作為第二信使激活Nrf2/ARE抗氧化信號通路[23],在本研究觀察時間內雖然未檢測到NCTD對HepG2細胞內ROS水平的影響,NCTD對HepG2細胞內Nrf2/ ARE信號通路的激活仍有可能與更早期的ROS產生有關。有研究表明,Nrf2/ARE信號通路與腫瘤細胞的耐藥性密切相關[24],Nrf2/ARE抗氧化信號通路的激活可以抵抗細胞內ROS的過量生成,若ROS與NCTD的抗腫瘤作用相關,Nrf2/ARE信號通路抑制劑與NCTD聯合用藥可能會增強NCTD的抗腫瘤作用,需進一步開展聯合用藥及效果評估研究。另外,大量研究表明,細胞的轉化及惡性腫瘤的發生與轉移與細胞內ROS的產生有關[25],NCTD可以激活Nrf2/ARE信號通路,為NCTD作為一種癌癥預防藥物的研發提供了思路。
[1]Pan CY,Gu XG.Research advance in effective mechanisms of norcantharidin in anti-liver cancer[J].J Mod Oncol(現代腫瘤醫學),2015,23(4):575-577.
[2]Li ZY,Ma QT,Zhang Y,Wang XC,Liu YL. Gemcitabine and cisplatin combined with norcan?tharidin sodium for the treatment of advanced NSCLC[J].Chin J Cancer Prev Treat(中華腫瘤防治雜志),2014,21(4):293-295.
[3]Peng C,Li Z,Niu Z,Niu W,Xu Z,Gao H,et al. Norcantharidin suppresses colon cancer cell epithelialmesenchymal transition by inhibiting the αvβ6-ERKEts1 signaling pathway[J].Sci Rep,2016,6:20500.
[4]Hsieh CH,Chao KS,Liao HF,Chen YJ.Nocantharidin,derivative of cantharidin,for cancer stem cells[J].Evid Based Complement Alternat Med,2013,2013:838651.
[5]Li XP,Jing W,Sun JJ,Liu ZY,Zhang JT,Sun W,etal.A potential small-molecule synthetic anti?lymphangiogenic agent norcantharidin inhibits tumor growth and lymphangiogenesis of human colonic adenocarcinomas through blocking VEGF-A,-C,-D/ VEGFR-2,-3″multi-points priming″mechanisms in vitro and in vivo[J].BMC Cancer,2015,15:527.
[6]Wu Y,Cao CM,Wang HQ,Li ZM,Sun ZX.Nor?cantharidin downregulates programmed cell death 4 expression in human gastric cancer cells[J]. Chin J Pharmacol Toxicol(中國藥理學與毒理學雜志),2013,27(4):622-628.
[7]Cui BD,Wang M,Sun ZX.The inhibition of norcantharidin on human lung adenocarcinoma A549 cells[J].Carcinog Teratog Mutag(癌變·畸變·突變),2015,27(1):21-25.
[8]Li ZM,Sun ZX.The effect and the synergistic mechanism of irinotecan combined with norcan?tharidin in human gastric cancer cell line BGC-823[J].Prog Biochem Biophys(生物化學與生物物理進展),2015,42(2):169-181.
[9]Lin SS,Huang HP,Yang JS,Wu JY,Hsia TC,Lin CC,et al.DNA damage and endoplasmic retic?ulum stress mediated curcumin-induced cell cycle arrest and apoptosis in human lung carcinoma A-549 cells through the activation caspases cascadeand mitochondrial-dependent pathway[J].Cancer Lett,2008,272(1):77-90.
[10]Liao YH,Tang Y,Qian NS,Dou KF.Oxidative stress and cell apoptosis[J].J Xinxiang Med Univ(新鄉醫學院學報),2011,28(1):110-113.
[11]Wang N,Ma HP,Qi XZ,Meng P,Jia ZP.Pro?gression of Nrf2-ARE signaling pathway in protection of oxidative stress injury of the body[J].Med Pharm J Chin PLA(解放軍醫藥雜志),2015,27(12):21-27.
[12]Hagos GK,Abdul-Hay SO,Sohn J,Edirisinghe PD,Chandrasena RE,Wang Z,et al.Anti-inflammatory,antiproliferative,and cytoprotective activity of NO chimera nitrates of use in cancer chemoprevention[J].Mol Pharmacol,2008,74(5):1381-1391.
[13]Xi MY,Jia JM,Sun HP,Sun ZY,Jiang JW,Wang YJ,et al.3-Aroylmethylene-2,3,6,7-tetra?hydro-1H-pyrazino[2,1-a]isoquinolin-4(11bH)-ones as potent Nrf2/ARE inducers in human cancer cells and AOM-DSS treated mice[J].J Med Chem,2013,56(20):7925-7938.
[14]Huang QT,Chen JH,Hang LL,Liu SS,Zhong M. Activation of PAR-1/NADPH oxidase/ROS signaling pathways is crucial for the thrombin-induced sFlt-1 production in extravillous trophoblasts:possible involvement in the pathogenesis of preeclampsia[J].Cell Physiol Biochem,2015,35(4):1654-1662.
[15]Liu D,Shi P,Yin X,Chen Z,Zhang X.Effect of norcantharidin on the human breast cancer Bcap-37 cells[J].Connect Tissue Res,2012,53(6):508-512.
[16]Maiese K.Environmental stimulus package:poten?tial for a rising oxidative deficit[J].Oxid Med Cell Longev,2009,2(4):179-180.
[17]Jorgensen ED,Zhao H,Traganos F,Albino AP,Darzynkiewicz Z.DNA damage response induced by exposure of human lung adenocarcinoma cells to smoke from tobacco-and nicotine-free cigarettes[J].Cell Cycle,2010,9(11):2170-2176.
[18]Cai R,Bu JG,Xu JJ,Zheng JF,Zhang JR.Recent progress in the molecular carcinogenic mechanism of heavy metal exposure[J].Med Recapitulate(醫學綜述),2014,20(10):1786-1789.
[19]Shen B,He PJ,Shao CL.Norcantharidin induced DU145 cell apoptosis through ROS-mediated mito?chondrial dysfunction and energy depletion[J]. PLoS One,2013,8(12):e84610.
[20]Chang C,Zhu YQ,Mei JJ,Liu SQ,Luo J.Involve?ment of mitochondrial pathway in NCTD-induced cytotoxicity in human hepG2 cells[J].J Exp Clin Cancer Res,2010,29(1):145.
[21]Hong XF,Li BS,Sun ZX.Inhibitory effect of nor?cantharidin on tubulin polymerization in vitro[J]. Chin J Pharmacol Toxicol(中國藥理學與毒理學雜志),2012,26(5):630-634.
[22]Li RJ,Hong XF,Ji YY,Sun ZX.Norcantharidin induced mitotic arrest and apoptosis in human ovarian cancer SK-OV-3 cells[J].Chin J Pharmacol Toxicol(中國藥理學與毒理學雜志),2013,27(2):180-186.
[23]Chen N,Hu R,Guo QL.Role of Nrf2 in chemopre?vention of cancer[J].Pharm Biotech(藥物生物技術),2010,17(5):452-456.
[24]Jaramillo MC,Zhang DD.The emerging role of the Nrf2-Keap1 signaling pathway in cancer[J].Genes Dev,2013,27(20):2179-2191.
[25]Xiong SS,Shi YY,Shi HP.Research status of reactive oxygen species and tumor[J].Chin J Caner Prev Treat(中華腫瘤防治雜志),2014,21(13):1045-1048.
Early events of norcantharidin reactive oxygen species and NF-E2-related factor 2/antioxidant response element pathway of HepG2 cells
CHEN Jing*,CUI Bao-di*,SUN Zhen-xiao
(College of Chinese Materia Medica,Beijing University of Chinese Medicine,Beijing 100102,China)
OBJECTIVETo investigate the early events of norcantharidin(NCTD)induced cell apoptosis and cell cycle arrest,the variation of reactive oxygen species(ROS)and the NF-E2-relate?dactor 2/antioxidant response element(Nrf2/ARE)pathway in human HepG2 cells.METHODSThe cyto?toxicity was measured by MTT assay.Apoptosis and cell cycle was analyzed by flow cytometry.The intra toxicity ROS production was evaluated by flow cytometry analysis with DCFH-DA probe and the effect of NCTD on Nrf2/ARE pathway was detected by luciferase assay in HepG2C8 cells under the same condition. The mRNA expression of heme oxygenase-1(HO-1)and NAD(P)H:quinone oxidoreductase 1(NQO1) antioxidase gene in Nrf2/ARE pathway downstream was evaluated by quantitative real-time PCR.RESULTSNo significant cytotoxicity was detected after HepG2 cells were treated with NCTD 30,60 and 120 μmol·L-1for 3 and 6 h,but cellular viability was inhibited significantly by NCTD 30,60 and 120 μmol·L-1for 24,48 and 72 h(P<0.01).Cell apoptosis and G2/M phase arrest occurred after HepG2 cells were treated with NCTD 60 μmol·L-1for 12,24 and 48 h.The percentage of apoptosis increased from(4.00±1.98)%to(12.10±1.70)%for 12 h,from(4.05±0.21)%to(31.8±6.50)%for 24 h,and from (3.90±0.85)%to(33.30±1.41)%for 48 h,respectively.The percentage of G2/M phase increased from (16.51±1.58)%to(40.89±0.18)%for 12 h,from(16.99±1.32)%to(55.29±3.99)%for 24 h,and from (14.45±0.59)%to(50.66±5.88)%for 48 h,respectively.Compared with cell control group,the percentage of G1phase had a significant decrease in the group with NCTD treated at different time points(P<0.01).No significant change in ROS in HepG2 cells was detected after the treatment with NCTD 30,60 and 120 μmol·L-1for 3,6 and 12 h.Nrf2/ARE pathway in HepG2C8 cells was activated by NCTD 30,60 and 120 μmol·L-1for 6 and 12 h.mRNA expression of HO-1 and NQO1 had a signifi?cant activation in HepG2 cells after treatment with NCTD 30,60 and 120 μmol·L-1for 6 and 12 h(P<0.05).CONCLUSIONNCTD can activate Nrf2/ARE pathway in the early stage in HepG2 cells,which may inhibit the intracellular ROS production in the early stage.Activation of ROS may not be the main event in NCTD induced HepG2 cell apoptosis and G2/M phase arrest.
norcantharidin;apoptosis;cell cycle;Nrf2-ARE pathway;reactive oxygen species; heme oxygenase-1;NAD(P)H:quinone oxidoreductase 1
SUN Zhen-xiao,E-mail:sunzxcn@hotmail.com,Tel:(010)84738646
R285.5,R979.1
A
1000-3002-(2017)01-0094-07
10.3867/j.issn.1000-3002.2017.01.012
2016-05-03接受日期:2016-09-18)
(本文編輯:喬 虹)
國家自然科學基金項目(81473418);國家林業局野生動植物保護項目(2012-2016)
陳 靜,女,碩士研究生,主要從事中藥分子細胞藥理學與毒理學研究,E-mail:476494894@qq.com,Tel:(010)84738646;崔寶弟,女,碩士研究生,主要從事中藥及其有效成分抗腫瘤分子細胞藥理學研究,Tel:(010)84738646,E-mail:1293772869@qq.com
孫震曉,E-mail:sunzxcn@hotmail.com,Tel:(010)84738646
*共同第一作者。
Foundation item:The project supported by National Natural Science Foundation of China(81473418);and Wildlife Protection Project of the State Forestry Administration(2012-2016)
*Co-first author.
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
鴨綠江(2021年35期)2021-04-19 12:24:18
當代陜西(2021年2期)2021-03-29 07:41:24
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
天津醫科大學學報(2019年3期)2019-08-13 06:53:08
電子制作(2018年11期)2018-08-04 03:25:42
中國塑料(2016年3期)2016-06-15 20:30:00
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
中國當代醫藥(2015年16期)2015-03-01 02:03:11
