飼料糖水平對大黃魚生長和糖代謝的影響
2017-04-12 09:49:13邢淑娟孫瑞健馬俊韋海明徐瑋周慧慧張彥嬌張文兵麥康森
水生生物學報 2017年2期
邢淑娟孫瑞健馬 俊韋海明徐 瑋周慧慧張彥嬌張文兵麥康森
(1. 中國海洋大學水產學院, 水產動物營養與飼料農業部重點實驗室, 海水養殖教育部重點實驗室, 青島 266003; 2. 通威股份有限公司技術中心, 成都 610041)
飼料糖水平對大黃魚生長和糖代謝的影響
邢淑娟1孫瑞健2馬 俊1韋海明1徐 瑋1周慧慧1張彥嬌1張文兵1麥康森1
(1. 中國海洋大學水產學院, 水產動物營養與飼料農業部重點實驗室, 海水養殖教育部重點實驗室, 青島 266003; 2. 通威股份有限公司技術中心, 成都 610041)
以初始體重為(137.5±0.4) g的大黃魚Larimichthys crocea 為實驗對象, 在海水浮式網箱中進行為期8周的攝食生長實驗, 研究飼料中糖水平對其生長、飼料利用、血液生化指標和糖代謝酶活力等的影響, 以確定大黃魚的飼料糖需求量。實驗飼料按等氮(粗蛋白質45%)等能(18 kJ/g)設計, 糖含量分別為1.75%、6.67%、13.64%、21.15%、26.69%和32.25%。結果表明隨著飼料糖水平的升高, 大黃魚特定生長率(SGR)先升高后降低, 當糖含量為26.69%時, SGR達最大值, 顯著高于糖含量為1.75%、6.67%、13.64%和32.25%處理組(P<0.05)。飼料效率(FER)和蛋白質效率(PER)均在糖含量為13.64%—21.15%時顯著高于其他處理組(P<0.05)。隨飼料中糖水平的升高, 全魚粗脂肪含量顯著降低, 在糖含量為32.25%時降至最低(10.56%), 顯著低于其他處理組(P<0.05)。肝體比和肝糖原含量均隨飼料糖水平的升高而顯著升高(P<0.05), 在糖含量為32.25%時達到最大值, 顯著高于糖含量為1.75%和6.67%處理組(P<0.05)。隨飼料糖水平的升高, 血漿甘油三酯和膽固醇水平均顯著降低(P<0.05), 而血糖水平不受飼料糖含量的影響(P>0.05)。大黃魚血清溶菌酶、脂蛋白脂酶和肝脂酶活性均隨飼料糖水平的升高顯著降低(P<0.05), 而腸淀粉酶活性表現為先升高后降低, 在糖含量為26.69%時, 酶活力達到最大值。隨飼料糖水平的升高, 大黃魚肝臟己糖激酶活性先上升后下降, 在糖含量為21.15%時達到最大值, 顯著高于糖含量為32.25%處理組(P<0.05), 而丙酮酸激酶活力在糖水平為32.25%時達到最大值, 顯著高于糖含量為1.75%和6.67%處理組(P<0.05)。用二次多項回歸模型擬合特定生長率和飼料糖水平的關系, 得到大黃魚飼料中最適糖含量為22.7%。
大黃魚; Larimichthys crocea; 糖; 營養; 飼料
在飼料中適量的糖和脂肪不僅可以節約飼料蛋白質, 提高飼料利用率, 還可以降低蛋白質代謝產物對養殖水體的污染[1—4]。然而, 飼料非蛋白源能量物質選擇不當或不適宜的添加量都會對魚類的生長、代謝和飼料效率產生不利影響[5,6]。
飼料脂肪除了提供魚體必需脂肪酸外, 還為魚體生理活動提供能量。攝入過量的脂肪不利于魚類的生長和代謝[6—8], 多余的脂肪還將以體脂肪的形式儲存, 影響魚肉品質[9]。與脂肪相比, 糖類具有價格低廉, 易儲存, 不易被氧化等優點。在飼料中以適宜的糖類物質替代脂肪, 可以直接為魚體提供能量, 減少攝入多余脂肪, 提高魚肉品質[10]。過高或過低的飼料糖含量, 會導致魚類生長緩慢和代謝紊亂[11—16]。適量的飼料糖水平可促進魚類生長, 提高飼料效率, 增強魚體免疫力, 并降低養殖成本, 提高養殖效益[6,17—21]。
大黃魚(Larimichthys crocea)是我國黃海南部、東海、臺灣海峽以及南海北部海水養殖的名貴經濟魚類。到目前為止, 有關大黃魚營養生理的研究已有相關報道[22—24]。并且, 在我們先前的研究中確定了較小規格(7.6—26.0 g)大黃魚幼魚對飼料糖的需求量為19%—21%, 但較大規格(體重>100 g)大黃魚的飼料糖需求量尚未見報道。本研究擬通過分析飼料中不同糖水平對大黃魚生長、消化、代謝和免疫反應等的影響, 確定較大規格大黃魚幼魚配合飼料中適宜的糖含量, 為大黃魚不同生長階段的精準飼料配方的設計提供基礎數據。
1 材料與方法
1.1 實驗飼料
以魚肉濃縮蛋白(來源于鱈)、酪蛋白和明膠為蛋白源, 魚油為脂肪源, 并以小麥淀粉梯度替代飼料中的魚油, 使飼料糖水平依次為1.75%、6.67%、13.64%、21.15%、26.69%和32.25%, 分別記為Diet 1—Diet 6 (表 1)。原料經過粉碎后過320 μm篩網,將粉碎好的原料按飼料配方逐級放大充分混勻后,加入魚油并手工搓勻, 隨后加入30%的水混合揉勻成糜團狀, 經F(II)-26 型雙螺桿擠條機(華南理工大學, 廣州)加工制成直徑為2.5 mm×3.0 mm的顆粒飼料。飼料在恒溫60℃的烘箱中烘干12h后, 再放于-20℃的冰箱保存待用。
1.2 實驗魚和養殖實驗
養殖實驗在浙江省寧波市象山縣鳳凰礁育苗場養殖基地進行, 實驗用大黃魚為當年人工培育的同一批苗種(由寧波市象山縣鳳凰礁育苗場提供)。正式實驗前, 實驗魚暫養于3 m×3 m ×3 m的浮式網箱中, 以實驗對照組飼料 (Diet 1) 飽食投喂2周, 使之逐漸適應實驗飼料和養殖環境。
實驗開始前, 停止投喂24h后挑選出體格健壯、規格一致的大黃魚進行稱重分組[初始體重: (137.5±0.35) g], 每個網箱 (1.5 m×1.5 m×2.0 m) 放養45尾, 每種實驗飼料隨機投喂3個網箱的實驗魚。每天飽食投喂2次(05:00和17:00), 養殖實驗持續8周, 每天記錄投餌量, 如有死亡則計數并稱重。實驗期間水溫為27—30℃, 鹽度為25‰—28‰, 溶氧約為7 mg/L。

表 1 實驗飼料配方及成分分析(%干重)Tab. 1 Formulation and proximate composition of the experimental diets (% dry matter)
1.3 樣品收集
8周養殖實驗結束, 實驗魚饑餓24h后以丁香酚(1∶10000)麻醉, 然后逐尾稱重和計數。每個網箱隨機取5尾實驗魚保存于-20℃冰箱, 用于常規分析。
每個網箱中隨機取5尾實驗魚, 分別測量體長、肝臟重、內臟重; 最后每個網箱隨機取8尾實驗魚, 麻醉后由尾靜脈取血, 并于4000×g離心10min (4℃)分離血清后立即置于-80℃保存待用。對完成取血的實驗魚, 迅速解剖取其肝臟、腸道和肌肉,液氮速凍后保存于-80℃冰箱用于酶活性的測定。
1.4 樣品分析測定方法
常規分析魚體水分、粗蛋白、粗脂肪及灰分測定參照AOAC (1995)方法。原料、飼料和魚體樣品在105℃烘箱中烘干至恒重, 得到各自水分含量; 用凱氏定氮儀(Kjeltec TM8400, FOSS, Sweden)測定粗蛋白; 索氏抽提儀(Buchi 36680, Switzerland) 測定粗脂肪; 在馬弗爐中灼燒(550℃, 16h)測定灰分; 飼料能量采用氧彈儀(Par 6100, Moline, IL, USA)測定; 還原糖含量采用3, 5-二硝基水楊酸法測定[25]。
糖脂代謝物相關指標血糖、血漿總膽固醇、血漿甘油三酯水平由青島大學附屬醫院測定。肝糖原、肌糖原、血清高/低密度脂蛋白膽固醇含量(HLD-C和LDL-C)、血漿總脂酶(脂蛋白脂酶LPL和肝脂酶HL)和血清溶菌酶均采用比色法檢測(試劑盒A043、A112-2、A113-2、A067和A050-1, 南京建成生物工程研究所, 中國)。其中, 血清總脂酶活性定義為: 每毫升每小時在反應系統中所產生的1微摩爾(μmol)的游離脂肪酸FFA為1個酶活性單位(FFAμmol/mL血清×小時)。溶菌酶活性定義為: 在25℃, pH 6.2條件下, 于450 nm處每分鐘使吸光度值降低0.001為一個酶活力單位。
消化酶活性消化酶液的制備: 準確稱取組織重量, 按樣品重的9倍加入預冷的磷酸緩沖液(0.01 mol/L, pH 7.4, 稀釋1/10), 高速組織勻漿機冰浴勻漿, 勻漿液在4℃下 10000×g離心30min, 取上清液進行消化酶活力測定。
肝臟和腸道的胰蛋白酶、淀粉酶活性均采用比色法檢測(試劑盒A080-2和C016, 南京建成生物工程研究所, 中國)。其中, 胰蛋白酶活性定義為:在pH 8.0, 37℃條件下, 每毫克蛋白中含有的胰蛋白酶每分鐘使吸光度變化0.003即一個酶活力單位。淀粉酶活力定義為: 組織中每毫克蛋白在37℃與底物作用30min, 水解10 mg淀粉即為一個淀粉酶活力單位。參考Bradford[26]的考馬斯亮蘭法測定肝胰臟勻漿液中蛋白質的含量, 以牛血清白蛋白作為標準蛋白。
糖代謝酶活性勻漿緩沖液的制備參照Polakof等[27]的方法, 其中含有50 mmol/L Tris (pH 7.6), 5 mmol/L EDTA, 2 mmol/L 1, 4-二硫蘇糖醇以及1%蛋白酶抑制劑混合物(Sigma Chemical, P-2714)。勻漿液在4℃下9000×g離心10min取上清后再10000×g離心20min, -80℃保存備用。
肝臟磷酸果糖激酶PFK和磷酸烯醇式丙酮酸羧激酶PEPCK活性測定參照Polakof等[27]的方法, 果糖-1, 6-二磷酸酶FBPase活性測定參照Susana Sangiao-Alvarellos等[28]的方法。參考Bradford[26]的考馬斯亮蘭法測定肝胰臟勻漿液中蛋白質的含量, 以牛血清白蛋白作為標準蛋白。
肝臟己糖激酶HK、丙酮酸激酶PK采用比色法檢測(試劑盒A077-1和A076-1, 南京建成生物工程研究所, 中國)。HK活性定義為: 在37℃, pH7.6的條件下, 每克組織蛋白在本反應體系中每分鐘生成1 mmol/L的NADP定義為一個酶活力單位。PK活性定義為: 在37℃, pH7.6的條件下, 每克組織蛋白每分鐘將1 μmol的PEP轉變成丙酮酸為一個酶活力單位。
1.5 計算公式和統計分析
存活率(Survival rate, SR, %)=100×終末尾數/初始尾數
特定生長率(Specific growth rate, SGR, %/d)= 100×[Ln(魚體終重)-Ln(魚體初重)]/實驗天數
飼料效率(Feed efficiency ratio, FER)=100×(魚體終重-魚體初重)/攝食量
蛋白質效率(Protein efficiency ratio, PER)= 100× (魚體終重-魚體初重)/飼料蛋白攝食量
肥滿度(CF, %)=100×魚體終體重/魚體長3
臟體比(Viserosomatic index, VSI, %)=100×內臟重/體重
肝體比(Hepatosomatic index, HSI, %)=100×肝臟重/體重
數據用平均值±標準誤(means±SEM)表示, 采用單因素方差分析(One-way ANOVA), 顯著性水平確定為0.05。當方差分析顯示處理間結果差異顯著時(P<0.05), 使用Tukey多重檢驗進行比較。數據分析采用的軟件為SPSS 17.0。以SGR為評價指標, 用二次曲線模型[29]來確定大黃魚的飼料糖需求量。
每天最少5升液體飼料,最多6升/天;開食料對瘤胃上皮的發育有重要作用,喂6升奶比喂8升奶能促使犢牛多采食1磅/天的開食料。
2 結果
2.1 飼料糖水平對大黃魚生長性能和飼料利用的影響
經過8周的養殖實驗, 大黃魚存活率均在95.56%以上, 且在各處理組間無顯著差異(P>0.05) (表 2)。
隨著飼料糖水平的升高, 各處理組特定生長率SGR 先升高后降低, 在糖含量為26.69%時達到最大(0.37±0.01) %/d, 顯著高于糖含量為1.75%、6.67%、13.64%和32.25%處理組 (P<0.05)。用二次曲線回歸模型擬合SGR和飼料糖水平的關系, 得到大黃魚飼料中糖的最適含量為22.7% (圖 1)。飼料效率FER和蛋白質效率PER與特定生長率SGR有相似的變化趨勢, 兩者均在糖含量為21.15%時達最大值,顯著高于糖含量為1.75%、6.67%和32.25%處理組(P <0.05)。
2.2 飼料糖水平對大黃魚體組成的影響
隨飼料糖水平的升高, 全魚粗脂肪含量顯著降低(P<0.05), 在糖含量為32.25%時降至最低(10.56%),顯著低于其他各處理組(P<0.05)。全魚水分(66.84%—68.97%)、粗蛋白(17.82%—18.24%)和灰分(3.45%—3.58%)均未受飼料糖水平的顯著影響 (P>0.05)(表3)。
2.3 飼料糖水平對大黃魚形體指標、肝糖原和肌糖原含量的影響
飼料糖水平對大黃魚肝體比HSI和肝糖原含量影響顯著(P<0.05), 隨飼料中糖含量的增加, 兩者均顯著升高且在糖含量為32.25%時達到最大值, 顯著高于糖含量為1.75%和6.67%處理組。大黃魚臟體比VSI (1.71%—2.36%)、肥滿度CF (1.55%—1.67%)、肌糖原含量(0.39—0.62 mg/g)在各處理間均無顯著性差異(P>0.05)(表 4)。
2.4 飼料糖水平對大黃魚血液指標的影響
大黃魚血漿總膽固醇、血漿甘油三酯水平均受飼料糖水平的顯著影響(P<0.05)(表 5)。血漿總膽固醇和甘油三酯水平隨飼料糖含量的增加均顯著降低, 在糖含量為32.25%時降至最小值, 顯著低于糖含量為1.75%處理組(P<0.05)。然而飼料糖水平對大黃魚血糖(3.32—4.48 mmol/L)、血清高/低密度脂蛋白膽固醇(1.63—2.13 mmol/L和1.17—1.92 mmol/L)水平均未產生顯著影響 (P>0.05)。血清脂蛋白脂酶、肝脂酶和溶菌酶活力均隨飼料糖含量的升高顯著降低(P<0.05), 且在糖含量為32.25%處均降至最低, 顯著低于糖含量為1.75%處理組(表5)。
2.5 飼料糖水平對大黃魚消化酶活力的影響
大黃魚腸道淀粉酶活力隨飼料糖水平的升高表現為先上升后下降的趨勢, 在糖含量為26.69%時活力達到最大值(0.47 U/mg pro.), 顯著高于糖含量為1.75%、6.67%、13.64%和32.25%處理組(P<0.05) (表 6)。然而, 飼料糖水平并未對魚體肝臟淀粉酶和胰蛋白酶、腸道胰蛋白酶活力產生顯著影響(P>0.05)。
2.6 飼料糖水平對大黃魚肝臟糖代謝酶活力的影響
肝臟糖酵解酶: 己糖激酶HK和丙酮酸激酶PK活力隨飼料糖含量的升高均顯著上升(P<0.05)。其中, HK活力在糖含量為21.15%時達到最大值(8.11 U/g prot.), 之后逐漸降低, 在糖含量為32.25%時, HK活力降至最低(4.6 U/g prot.), 顯著低于糖含量為21.15%處理組(P<0.05)。PK活性在糖含量為32.25%時達到最大值(30.28 U/g prot.), 顯著高于糖含量為1.75%和6.67%處理組(P<0.05)。肝臟磷酸果糖激酶PFK (3.75—9.14 U/g prot.)活性不受飼料糖水平的影響, 各組間無顯著差異(P>0.05)。

表 2 飼料糖水平對大黃魚生長效應和飼料利用的影響Tab. 2 Effects of dietary carbohydrate on growth performance and feed utilization of large yellow croaker

圖 1 大黃魚的特定生長率(SGR)與飼料糖水平的關系Fig. 1 Relationship between dietary carbohydrate levels and the specific growth rate (SGR) of large yellow croaker Larimichthys crocea fed with the experimental diets for 8 weeks
肝臟糖異生酶: 果糖-1, 6-二磷酸酶FBPase (11.92—17.68 U/g prot.)和磷酸烯醇式丙酮酸羧激酶PEPCK (9.95—12.23 U/g prot.)活性均不受飼料糖水平的顯著影響(P>0.05)(表 7)。
3 討論
本研究結果顯示, 在飼料中適量的添加糖顯著提高了大黃魚的生長和飼料利用。在糖含量為21.15%和26.69%時(糖脂比分別為2.29和4.41), SGR達到最大值, 顯著高于其他各處理組, 飼料糖含量的進一步增加反而顯著降低了SGR, 表明飼料中糖含量過少或過量均導致魚體生長受到抑制。同時, 當飼料中糖含量過高或過低時(糖脂比為9.92和0.10), 大黃魚FER和PER顯著低于其他處理組, 表明等氮等能飼料中, 可用適量的糖替代脂肪, 充分發揮魚體內脂肪和糖代謝的協同效應[21], 從而有利于提高魚類生長和飼料利用率[11]。飼料中適量糖的促生長和對蛋白質的節約效應可能是因為葡萄糖是組織中的優先氧化底物并且能抑制魚體內糖異生途徑, 從而有效地減少了氨基酸進入氧化途徑[30]。同樣, 在對許氏平鲉[13]、長吻[9]、尖齒胡子鯰[31]、蟾胡鯰[6]和眼斑擬石首魚[32]等的研究中也有類似的發現。然而, 也有研究表明, 等氮等能飼料中糖水平不會對魚體的生長產生顯著影響[5,10,33—35]。不同的實驗結果可能是由于實驗魚的種類、生長階段、飼料糖源、糖脂比例、實驗條件和周期等的差異造成。用二次多項回歸模型擬合SGR和飼料糖水平的關系, 得到飼料中糖的最適含量為22.7%。這一結果與尖吻鱸20%[21]、長吻23.4%[9]、虹鱒18%[36]、軍曹魚21.1%[14]、許氏平鲉19.8%[13]等魚類的飼料中糖的最適含量相似。
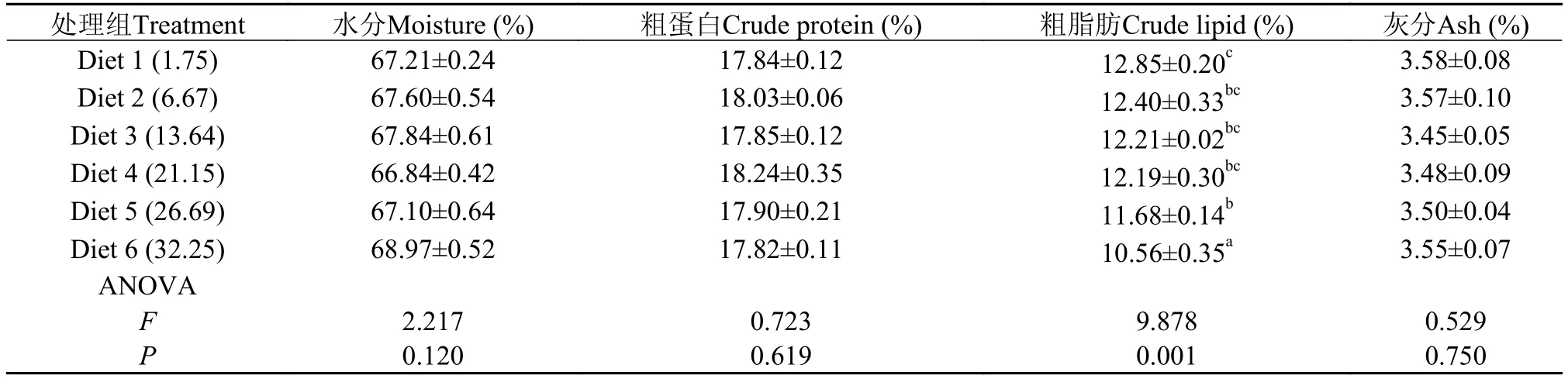
表 3 飼料糖水平對大黃魚體組成的影響Tab. 3 Effects of dietary carbohydrate levels on the body compositions of large yellow croaker

表 4 飼料糖水平對大黃魚形體指數、肝糖原和肌糖原含量的影響Tab. 4 Effects of dietary carbohydrate levels on body index, liver glycogen and muscle glycogen contents of large yellow croaker

表 5 飼料糖水平對大黃魚血液指標的影響Tab. 5 Effects of dietary carbohydrate levels on plasma parameters of large yellow croaker

表 6 飼料糖水平對大黃魚肝臟和腸消化酶活力的影響Tab. 6 Effects of dietary carbohydrate levels on digestive enzyme activities in liver and intestinal tract of large yellow croaker

表 7 飼料糖水平對大黃魚肝臟糖酵解和糖異生酶活性的影響Tab. 7 Effects of dietary carbohydrate levels on the activities of glycolysis and gluconeogenesis enzyme in the liver of large yellow croaker
在本研究中各處理組飼料能量相等, 為保證在等氮的前提下飼料中糖含量的梯度, 各實驗飼料間脂肪和纖維素含量差異較大。有研究表明, 飼料中纖維素含量增加會導致魚體對飼料中其他營養素的利用率降低[37]。然而, Wiesmann和Pfeffer[38]研究發現飼料中的纖維素并不影響虹鱒對飼料蛋白質和能量的利用, 并且在魚類營養學研究中微晶纖維素作為填充劑被廣泛采用[39,40]。此外, 飼料中微晶纖維素含量對魚體產生的影響還與魚的種類和食性有關[17]。在飼料中添加40%的微晶纖維素并沒有對斑點叉尾(Ictalurus punctatus)和羅非魚(Tilapia zillii)的生長產生顯著性影響[5,41]。由此可見, 本研究中微晶纖維素對大黃魚產生的影響還需要進一步研究確定。
在本研究中, 對于高脂低糖飼料組, 由于脂肪過高導致魚攝食減少[31], 從而減少了蛋白質和其他營養素的攝入, 最終導致生長緩慢, 飼料效率和蛋白質效率顯著降低。對于高糖低脂飼料組, 飼料提供的必需脂肪酸含量可能不足以滿足魚體的最佳生長需要, 同時高糖飼料導致魚體血糖迅速升高,產生飽足感從而抑制攝食, 最終影響其生長[6,11]。已有研究表明, 魚類特別是肉食性魚類對高糖飼料耐受性較差, 高糖飼料的攝入將導致體內代謝紊亂,進而抑制其生長[42]。此外, 不論高脂低糖還是高糖低脂飼料組, 微晶纖維素含量過高或過低均會影響大黃魚胃排空時間和實驗飼料的適口性, 從而影響魚類正常攝食。總之, 大黃魚攝食高脂低糖或高糖低脂飼料均對其生長、代謝和飼料利用產生不利影響。在本研究中, 高糖低脂組(Diet 6)的SGR顯著高于高脂低糖組(Diet 1), 由此推測在飼料蛋白質和能量含量一定的情況下, 相對于飼料脂肪, 大黃魚能在一定范圍內更好地利用飼料中的糖作為能量來源。當然, 該推測還需要進一步的研究。
飼料中任何非蛋白質能量的來源或含量的不平衡均會影響魚類生長、營養物質利用和體脂肪沉積[43]。在本研究中, 無論是高脂低糖組(糖脂比為0.10)還是高糖低脂組(糖脂比為9.92), 大黃魚SGR、PER和魚體粗脂肪含量均顯著降低。已有研究表明, 飼料糖脂比能顯著影響糖類對蛋白質的節約效應, 適宜的飼料糖脂比CHO/L能夠充分發揮糖和脂肪的協同效應[15], 從而促進魚類生長和飼料利用[11]。在本研究中, 飼料中糖脂比為1.11和2.29時, 大黃魚FER和PER均顯著高于糖脂比為0.10和9.92時的值。本研究結果顯示, 大黃魚飼料最適糖脂比為2.29—4.41, 這與在虹鱒(CHO/L 2.45)、長吻(CHO/L 1.98)、蟾胡鯰(CHO/L 3.389)、尖齒胡子鯰(CO/L 1.7—3.4)上的研究結果相近[6,9,31,36]。
飼料中的脂肪含量是影響魚體脂肪含量的最重要因素[44], 魚體脂肪含量會隨著攝入脂肪量的增加而上升[10]。Hemre和Kahrs[45]采用同位素標記葡萄糖研究大西洋鱈體內脂肪重新合成, 發現只有少量的的糖作為碳源合成脂肪, 飼料中的糖類促進機體脂肪重新合成主要通過增加細胞質內的還原型NADPH的量, 而不是通過為脂肪的合成提供碳架。在本研究中, 在飼料糖含量增加的同時, 飼料脂肪含量在降低(飼料糖脂比含量升高), 全魚粗脂肪含量顯著減少。這表明相對于糖類, 飼料脂肪更容易轉化成魚體脂肪并沉積。全魚粗脂肪和飼料中脂肪含量的正相關性與在長吻[9]、星斑川鰈[33]、尖齒胡子鯰[31]、尖吻鱸[10]以及蟾胡鯰[6]等魚類中的研究結果一致。
肝糖原是一種應急能源儲備。當出現應激時,肝糖原很容易被合成存儲或分解利用, 從而造成肝糖原的升高或降低[46]。Brauge等[28]研究表明, 隨著飼料糖含量的增加, 由14C標記的葡萄糖轉化成肝臟脂肪的含量也隨之增加, 攝食高糖飼料會誘導虹鱒脂肪合成, 并增加肝臟脂肪的沉積。在本研究中,大黃魚的肝體比HSI和肝糖原含量隨飼料糖含量的增加而顯著升高, 其中的原因可能是大黃魚能將飼料中過量的糖轉化成糖原儲存在肝臟中, 進而導致肝體比的增大。在對長吻[9]、南方鲇[47]、歐州鱸[48,49]、星斑川鰈[33]、金頭鯛[50,51]和虹鱒[52,53]等魚類的研究中也得到了相似結論。然而在對尖齒胡子鯰[31]和雜交條紋鱸[54]的研究發現, 飼料糖含量并不影響魚體肝糖原含量和肝體比。在對塞內加爾鰨[35]和許氏平鲉[13]的研究中發現飼料糖含量的增加反而使魚體肝糖原含量和肝體比顯著降低。不同的實驗結果除了來自魚種類的差異外, 不同的生長階段、飼料配方、飼料糖水平設計范圍、實驗條件和周期等也造成了結果的差異。
肝臟腫大和肝糖原的過度累積可能導致肝臟解毒能力下降等免疫抑制反應[55]。在本研究中, 高糖飼料顯著降低了大黃魚血清溶菌酶的活性, 可能是由于肝糖原和肝臟脂肪的過度累積在一定程度上造成的免疫抑制反應。已有研究表明, 高血糖水平和高肝體比導致虹鱒對水中銅離子的耐受力降低[56]。然而, 肝糖原的累積可在一定程度上緩解魚類攝食高糖飼料后導致的持續高血糖反應[57], 同時減少丙酮酸、乳酸和乙酰輔酶等血糖代謝中間產物對魚體造成的影響[58,59]。在本研究中, 隨著飼料糖水平的升高, 大黃魚血糖水平并沒有發生顯著變化, 可能與大量的肝糖原累積有關, 在一定程度上緩解了高糖飼料對魚體生理代謝平衡產生的影響。
血漿甘油三酯和膽固醇是魚類血脂的重要組成成分, 具有供給能量、合成某些酶、維生素和激素等作用, 是維持正常生命活動所必需的物質[60]。血漿甘油三酯一般由飼料攝取的脂肪分解以及肝臟中轉運出來的極低密度脂蛋白VLDL水解產生。在本研究中大黃魚血漿甘油三酯水平隨飼料脂肪含量的降低而顯著下降, 可能是因為攝入的脂肪減少, 同時脂蛋白脂酶LPL活力的顯著降低導致從肝臟轉運出來的VLDL水解減弱, 最終導致了血漿甘油三酯水平的顯著下降, 這也從一定程度上說明大黃魚體內的脂肪沉積主要是來自飼料脂肪的攝入而不是脂肪合成。在高脂低糖實驗組中, 魚體為了應對高脂飼料而加強了對脂肪的運輸, 表現為血漿膽固醇水平也顯著升高。在對大菱鲆[61]、羅非魚[62]、團頭魴[63]和長吻[9]等的研究中也發現了相似的結論。
大量研究表明, 魚類能夠根據攝食不同的飼料來調節自身消化酶的分泌, 但是這種調節能力隨魚種類不同而異[64]。一般來說, 雜食性和草食性魚類的調節能力遠高于肉食性魚類[63]。在本研究中, 隨飼料糖含量的升高, 大黃魚腸道淀粉酶活力顯著提高, 表明大黃魚對飼料中的糖類具有一定的適應能力。這與在鯉[65]、胡鯰[66]和尖齒胡子鯰[67]的研究結果一致。然而, 飼料糖含量的進一步升高(26.69%—32.25%, 糖脂比由4.41升高至9.92)顯著降低了大黃魚腸道淀粉酶活性, 可能是由于當飼料糖含量為26.69%(糖脂比為4.41)時, 淀粉酶活性已接近飽和,過多糖的攝入反而抑制了其活性。此外, 可能還與飼料脂肪含量降低、脂肪酶活性變化有關, 這有待進一步研究。
本研究結果顯示, 飼料糖含量的升高顯著提高了大黃魚肝臟丙酮酸激酶PK活力, 這在一定程度上促進了葡萄糖酵解, 從而緩解了魚體高血糖癥狀。同樣地, 有關歐洲鱸[68]、金頭鯛[69]和河鱸[70]的研究表明高糖飼料能顯著提高肝臟PK活力。然而,一些研究發現飼料糖水平對魚類的PK活力沒有產生顯著影響[35,53,71,72]。Panserat等[73]用高糖飼料喂養虹鱒, 其PK的mRNA表達量無顯著變化。因此,魚類PK活力的調節可能是由基因轉錄后的修飾作用完成, 如PK酶原的磷酸化或去磷酸化作用。因此, 飼料糖水平對魚類肝臟PK活力的影響仍需進一步研究。
在本研究中大黃魚肝臟已糖激酶HK活力在飼料糖含量為21.15% (糖脂比為2.29)時達到最大值(8.11 U/g prot), 之后隨糖含量的進一步升高, HK活性反而顯著降低, 在糖含量為32.25% (糖脂比為9.92)時降至最低。HK活力的降低可能是因為其活性受到其產物(葡萄糖-6-磷酸)的負反饋抑制[74], 這在一定程度上可視為大黃魚對高糖飼料的一種負反饋調節。在對鱸的研究中也發現飼料糖水平能顯著提高肝臟HK活力[70]。然而, 在對虹鱒[75,76]、金頭鯛[71]和歐洲鱸[68,77]的研究中并未發現飼料糖水平對其肝臟HK活力產生顯著影響。
4 結論
在本研究中, 飼料糖含量在21.15%—26.67% (糖脂比2.29—4.41)時, 大黃魚取得了最高的飼料效率、最佳的SGR和生理狀態。用二次曲線回歸模型擬合特定生長率和飼料糖水平的關系, 得到大黃魚(130 g)飼料中糖的最適含量為22.7%。
[1]Fernández F, Miquel A G, Córdoba M, et al. Effects of diets with distinct protein-to-carbohydrate ratios on nutrient digestibility, growth performance, body composition and liver intermediary enzyme activities in gilthead sea bream (Sparus aurata, L. ) fingerlings [J]. Journal of Experimental Marine Biology and Ecology, 2007, 343(1): 1—10
[2]Stone D, Allan G L, Anderson A J. Carbohydrate utilization by juvenile silver perch, Bidyanus bidyanus (Mitchell). III. The protein-sparing effect of wheat starchbased carbohydrates [J]. Aquaculture Research, 2003, 34(2): 123—134
[3]Dias J, Alvarez M J, Diez A, et al. Regulation of hepatic lipogenesis by dietary protein/energy in juvenile European seabass (Dicentrarchus labrax) [J]. Aquaculture, 1998, 161(1): 169—186
[4]Peres H, Goncalves P, Oliva-Teles A. Glucose tolerance in gilthead seabream (Sparus aurata) and European seabass (Dicentrarchus labrax) [J]. Aquaculture, 1999, 179(1): 415—423
[5]El-Sayed A M, Garling Jr D L. Carbohydrate-to-lipid ratios in diets for Tilapia zillii fingerlings [J]. Aquaculture, 1988, 73(1): 157—163
[6]Jafri A K. Effect of dietary carbohydrate-to-lipid ratio on growth and body composition of walking catfish (Clarias batrachus) [J]. Aquaculture, 1998, 161(1): 159—168
[7]Kj?r M A, Todor?evi? M, Torstensen B E, et al. Dietary n-3 HUFA affects mitochondrial fatty acid β-oxidation capacity and susceptibility to oxidative stress in Atlantic salmon [J]. Lipids, 2008, 43(9): 813—827
[8]Tan Q, Xie S, Zhu X, et al. Effect of dietary carbohydrate-to-lipid ratios on growth and feed utilization in Chinese longsnout catfish (Leiocassis longirostris Günther) [J]. Journal of Applied Ichthyology, 2007, 23(5): 605—610
[9]Hanley F. Effects of feeding supplementary diets containing varying levels of lipid on growth, food conversion, and body composition of Nile tilapia, Oreochromis niloticus (L. ) [J]. Aquaculture, 1991, 93(4): 323—334
[10]Nankervis L, Matthews S J, Appleford P. Effect of dietary non-protein energy source on growth, nutrient retention and circulating insulin-like growth factor I and triiodothyronine levels in juvenile barramundi, Lates calcarifer [J]. Aquaculture, 2000, 191(4): 323—335
[11]Hemre G I, Mommsen T P, Krogdahl ?. Carbohydrates in fish nutrition: effects on growth, glucose metabolism and hepatic enzymes [J]. Aquaculture Nutrition, 2002, 8(3): 175—194
[12]Cowey C B, Walton M J. Intermediary metabolism [J]. Fish Nutrition, 1989, 2: 259—329
[13]Moon T W. Glucose intolerance in teleost fish: fact or fic-tion [J]? Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 2001, 129(2): 243—249
[14]Wilson R P. Utilization of dietary carbohydrate by fish [J]. Aquaculture, 1994, 124(1): 67—80
[15]Brauge C, Corraze G, Médale F. Effects of dietary levels of carbohydrate and lipid on glucose oxidation and lipogenesis from glucose in rainbow trout, Oncorhynchus mykiss, reared in freshwater or in seawater [J]. Comparative Biochemistry and Physiology Part A: Physiology, 1995, 111(1): 117—124
[16]Catacutan M R, Coloso R M. Growth of juvenile Asian seabass, Lates calcarifer, fed varying carbohydrate and lipid levels [J]. Aquaculture, 1997, 149(1): 137—144
[17]Gao W, Liu Y J, Tian L X, et al. Effect of dietary carbohydrate-to-lipid ratios on growth performance, body composition, nutrient utilization and hepatic enzymes activities of herbivorous grass carp (Ctenopharyngodon idella) [J]. Aquaculture Nutrition, 2010, 16(3): 327—333
[18]Vielma J, Koskela J, Ruohonen K, et al. Optimal diet composition for European whitefish (Coregonus lavaretus): carbohydrate stress and immune parameter responses [J]. Aquaculture, 2003, 225(1): 3—16
[19]Lee S M, Kim K D. Effects of dietary carbohydrate to lipid ratios on growth and body composition of juvenile and grower rockfish, Sebastes schlegeli [J]. Aquaculture Research, 2009, 40(16): 1830—1837
[20]Ren M, Ai Q, Mai K, et al. Effect of dietary carbohydrate level on growth performance, body composition, apparent digestibility coefficient and digestive enzyme activities of juvenile cobia, Rachycentron canadum L [J]. Aquaculture Research, 2011, 42(10): 1467—1475
[21]Jafri A K. Growth rate, feed conversion, and body composition of Catla catla, Labeo rohita, and Cirrhinus mrigala fry fed diets of various carbohydrate-to-lipid ratios [J]. Journal of the World Aquaculture Society, 1998, 29(1): 84—91
[22]Duan Q, Mai K, Zhong H, et al. Studies on the nutrition of the large yellow croaker, Pseudosciaena crocea R. I: growth response to graded levels of dietary protein and lipid [J]. Aquaculture Research, 2001, 32(s1): 46—52
[23]Ma H, Cahu C, Zambonino J, et al. Activities of selected digestive enzymes during larval development of large yellow croaker (Pseudosciaena crocea) [J]. Aquaculture, 2005, 245(1): 239—248
[24]Mai K, Zhang C, Ai Q, et al. Dietary phosphorus requirement of large yellow croaker, Pseudosciaena crocea R [J]. Aquaculture, 2006, 251(2): 346—353
[25]Yu S, Olsen C E, Marcussen J. Methods for the assay of 1, 5-anhydro-D-fructose and α-1, 4-glucan lyase [J]. Carbohydrate Research, 1997, 305(1): 73—82
[26]Bradford M M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding [J]. Analytical Biochemistry, 1976, 72(1): 248—254
[27]Polakof S, Miguez J M, Soengas J L. Dietary carbohydrates induce changes in glucosensing capacity and food intake of rainbow trout [J]. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology, 2008, 295(2): R478-R489
[28]Sangiao-Alvarellos S, Laiz-Carrión R, Guzmán J M, et al. Acclimation of S. aurata to various salinities alters energy metabolism of osmoregulatory and nonosmoregulatory organs [J]. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology, 2003, 285(4): R897-R907
[29]Zeitoun I H, Ullrey D E, Magee W T, et al. Quantifying nutrient requirements of fish [J]. Journal of the Fisheries Board of Canada, 1976, 33(1): 167—172
[30]Sánchez muros M J, García rejón L, Lupianez J A, et al. Long-term nutritional effects on the primary liver and kidney metabolism in rainbow trout (Oncorhynchus mykiss). II. Adaptive response of glucose 6-phosphate dehydrogenase activity to high-carbohydrate/low-protein and high-fat/non-carbohydrate diets [J]. Aquaculture Nutrition, 1996, 2(4): 193—200
[31]Ali M Z, Jauncey K. Optimal dietary carbohydrate to lipid ratio in African catfish Clarias gariepinus (Burchell 1822) [J]. Aquaculture International, 2004, 12(2) : 169—180
[32]Ellis S C, Reigh R C. Effects of dietary lipid and carbohydrate levels on growth and body composition of juvenile red drum, Sciaenops ocellatus [J]. Aquaculture, 1991, 97(4): 383—394
[33]LEE S M, Lee J H. Effect of dietary glucose, dextrin and starch on growth and body composition of juvenile starry flounder Platichthys stellatus [J]. Fisheries Science, 2004, 70(1): 53—58
[34]Mohapatra M, Sahu N P, Chaudhari A. Utilization of gelatinized carbohydrate in diets of Labeo rohita fry [J]. Aquaculture Nutrition, 2003, 9(3): 189—196
[35]Dias J, Rueda Jasso R, Panserat S, et al. Effect of dietary carbohydrate-to-lipid ratios on growth, lipid deposition and metabolic hepatic enzymes in juvenile Senegalese sole (Solea senegalensis, Kaup) [J]. Aquaculture Research, 2004, 35(12): 1122—1130
[36]Yamamoto T, Konishi K, Shima T, et al. Influence of dietary fat and carbohydrate levels on growth and body composition of rainbow trout Oncorhynchus mykiss under self-feeding conditions [J]. Fisheries Science, 2001, 67(2): 221—227
[37]Kirchgessener M, Kürzinger H, Schwarz F J. Digestibility of crude nutrients in different feeds and estimation of their energy content for carp (Cyprinus carpio L. ) [J]. Aquaculture, 1986, 58(3—4): 185—194
[38]Wiesmann D, Pfeffer E. Influence of indigestible carbo-hydrates on the efficiency of utilization of dietary energy and protein in growing rainbow trout (Salmo gairdnerii, R) [J]. Archives of Animal Nutrition, 1986, 36(12): 1145—1149
[39]Mishra K, Samantaray K. Interacting effects of dietary lipid level and temperature on growth, body composition and fatty acidprofile of rohu, Labeo rohita (Hamilton) [J]. Aquaculture Nutrition, 2004, 10(6): 359—369
[40]Raven P A, Devlin R H, Higgs D A. Influence of dietary digestible energy content on growth, protein and energy utilization and body composition of growth hormone transgenic and nontransgenic coho salmon (Oncorhynchus kisutch) [J]. Aquaculture, 2006, 254(1—4): 730—747
[41]Garling Jr D L, Wilson R P. Effects of dietary carbohydrate-to-lipid ratios on growth and body composition of fingerling channel catfish [J]. The Progressive Fish-Culturist, 1977, 39(1—4): 43—47
[42]Masayuki furuichi, Yasuo yone. Change of blood sugar and plasma insulin levels of fishes in glucose tolerance test [J]. Bulletin of the Japanese Society of Scientific Fisheries, 1981, 47(6): 761—764
[43]Garling Jr D L, Wilson R P. Optimum dietary protein to energy ratio for channel catfish fingerlings, Ictalurus punctatus [J]. The Journal of Nutrition, 1976, 106(9): 1368—1375
[44]Sargent J, Henderson R J, Tocher D R. The lipids [A]. In: Fish Nutrition. In: Halver J E (Eds. ), Fish Nutrition [M]. New York: Academic Press. 1989, 154—209
[45]Hemre G I, Kahrs F.14C-glucose injection in Atlantic cod, Gadus morhua, metabolic responses and excretion via the gill membrane [J]. Aquaculture Nutrition, 1997, 3(1): 3—8
[46]Soengas J L, Aldegunde M, Andrés M D. Gradual transfer to sea water of rainbow trout: effects on liver carbohydrate metabolism [J]. Journal of Fish Biology, 1995, 47(3): 466—478
[47]Luo Y, Xie X. Effects of body lipid content on the resting metabolic rate and postprandial metabolic response in the southern catfish Silurus meridionalis [J]. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 2009, 154(4): 547—550
[48]Moreira I S, Peres H, Couto A, et al. Temperature and dietary carbohydrate level effects on performance and metabolic utilisation of diets in European sea bass (Dicentrarchus labrax) juveniles [J]. Aquaculture, 2008, 274(1): 153—160
[49]Peres H, Oliva-Teles A. Utilization of raw and gelatinized starch by European sea bass (Dicentrarchus labrax) juveniles [J]. Aquaculture, 2002, 205(3): 287—299
[50]Bou M, Todor?evi? M, Fontanillas R, et al. Adipose tissue and liver metabolic responses to different levels of dietary carbohydrates in gilthead sea bream (Sparus aurata) [J]. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 2014, 175(1): 72—81
[51]Couto A, Enes P, Peres H, et al. Effect of water temperature and dietary starch on growth and metabolic utilization of diets in gilthead sea bream (Sparus aurata) juveniles [J]. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 2008, 151(1): 45—50
[52]Capilla E, Médale F, Navarro I, et al. Muscle insulin binding and plasma levels in relation to liver glucokinase activity, glucose metabolism and dietary carbohydrates in rainbow trout [J]. Regulatory Peptides, 2003, 110(2): 123—132
[53]Hilton J W, Atkinson J L. Response of rainbow trout (Salmo gairdneri) to increased levels of available carbohydrate in practical trout diets [J]. British Journal of Nutrition, 1982, 47(3): 597—607
[54]Nematipour G R, Brown M L, Gatlin D M. Effects of dietary carbohydrate: lipid ratio on growth and body composition of hybrid striped bass [J]. Journal of the World Aquaculture Society, 1992, 23(2): 128—132
[55]Cai C F. Study on the utilization of dietary carbohydrate by Myloparyngodon piecus Richrdson and Carassius auratus and their mechanism of metabolism [D]. Thesis for Doctor of Science. East China Normal University, Shanghai. 2004 [蔡春芳. 青魚和鯽對飼料糖的利用及其代謝機制的研究. 博士學位論文, 華東師范大學, 上海. 2004]
[56]Dixon D G, Hilton J W. Influence of available dietary carbohydrate content on tolerance of waterborne copper by rainbow trout, Salmo gairdneri Richardson [J]. Journal of Fish Biology, 1981, 19(5): 509—518
[57]Lin X Z, Luo Y P, Xie X J. Effect of dietary carbohydrate level on glycolytic enzymes and serum glucose concentrations in the juvenile southern catfish after feeding [J]. Acta Hydrobiologica Sinica, 2006, 30(3): 304—310 [林小植, 羅毅平, 謝小軍. 飼料碳水化合物水平對南方鲇幼魚餐后糖酵解酶活性及血糖濃度的影響.水生生物學報, 2006, 30(3): 304—310]
[58]Nakano K, Tagawa M, Takemura A, et al. Temporal changes in liver carbohydrate metabolism associated with seawater transfer in Oreochromis mossambicus [J]. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 1998, 119(4): 721—728
[59]Luo Y P. Ecophysiological responses in a carnivorous fish, the southern catfish (Silurus meridionalis Chen) to nutritional stress of dietary carbohydrate [D]. Thesis for Doctor of Science. Southwest University, Chongqing. 2007 [羅毅平. 肉食性魚類南方鲇對飼料碳水化合物營養脅迫的生理生態學反應. 博士學位論文, 西南大學,重慶. 2007]
[60]Likimani T A, Wilson R P. Effects of diet on lipogenicenzyme activities in channel catfish hepatic and adipose tissue [J]. The Journal of Nutrition, 1982, 112(1): 112—117
[61]Regost C, Arzel J, Cardinal M, et al. Dietary lipid level, hepatic lipogenesis and flesh quality in turbot (Psetta maxima) [J]. Aquaculture, 2001, 193(3): 291—309
[62]Shimeno S, Ming D, Takeda M. Metabolic response to dietary carbohydrate to lipid ratios in Oreochromis niloticus [J]. Nippon Suisan Gakkaishi, 1993, 59(5): 827—833
[63]Li X, Jiang Y, Liu W, et al. Protein-sparing effect of dietary lipid in practical diets for blunt snout bream (Megalobrama amblycephala) fingerlings: effects on digestive and metabolic responses [J]. Fish Physiology and Biochemistry, 2012, 38(2): 529—541
[64]Moro G V, Camilo R Y, Moraes G, et al. Dietary nonprotein energy sources: growth, digestive enzyme activities and nutrient utilization by the catfish jundiá, Rhamdia quelen [J]. Aquaculture Research, 2010, 41(3): 394—400
[65]Kawai S, Ikeda S. Studies on digestive enzymes of fishes. 2. Effect of dietary change on activities of digestive enzymes in carp intestine [J]. Bulletin of the Japanese Society of Scientific Fisheries, 1972, 38(3): 265
[66]Mukhopadhyay P K. Studies on the enzymatic activities related to varied pattern of diets in the air-breathing cat fish, Clarias batrachus (Linn. ) [J]. Hydrobiologia, 1977, 52(2): 235—237
[67]Ali M Z, Jauncey K. Effect of dietary lipid to carbohydrate ratios on body composition, digestive enzyme activities and blood plasma components in African catfish Clarias gariepinus (Burchell, 1822) [J]. Journal of Aquaculture in the Tropics, 2005, 20: 57—70
[68]Enes P, Panserat S, Kaushik S, et al. Effect of normal and waxy maize starch on growth, food utilization and hepatic glucose metabolism in European sea bass (Dicentrarchus labrax) juveniles [J]. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 2006, 143(1): 89—96
[69]Fernández F, Miquel A G, Córdoba M, et al. Effects of diets with distinct protein-to-carbohydrate ratios on nutrient digestibility, growth performance, body composition and liver intermediary enzyme activities in gilthead sea bream (Sparus aurata, L. ) fingerlings [J]. Journal of Experimental Marine Biology and Ecology, 2007, 343(1): 1—10
[70]Borrebaek B, Christophersen B. Hepatic glucose phosphorylating activities in perch (Perca fluviatilis) after different dietary treatments [J]. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 2000, 125(3): 387—393
[71]Enes P, Panserat S, Kaushik S, et al. Hepatic glucokinase and glucose-6-phosphatase responses to dietary glucose and starch in gilthead sea bream (Sparus aurata) juveniles reared at two temperatures [J]. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 2008, 149(1): 80—86
[72]Suárez M D, Sanz A, Bazoco J, et al. Metabolic effects of changes in the dietary protein: carbohydrate ratio in eel (Angilla anguilla) and trout (Oncorhynchus mykiss) [J]. Aquaculture International, 2002, 10(2): 143—156
[73]Panserat S, Plagnes-Juan E, Kaushik S. Nutritional regulation and tissue specificity of gene expression for proteins involved in hepatic glucose metabolism in rainbow trout (Oncorhynchus mykiss) [J]. Journal of Experimental Biology, 2001, 204(13): 2351—2360
[74]Pen-Hsing Tung, Shi-Yen Shiau. Effects of meal frequency on growth performance of hybrid tilapia, Oreochromis niloticus×O. aureus, fed different carbohydrate diets [J]. Aquaculture, 1991, 92: 343—350
[75]Cowey C B, Knox D, Walton M J, et al. The regulation of gluconeogenesis by diet and insulin in rainbow trout (Salmo gairdneri) [J]. British Journal of Nutrition, 1977, 38(3): 463—470
[76]Kirchner S, Seixas P, Kaushik S, et al. Effects of low protein intake on extra-hepatic gluconeogenic enzyme expression and peripheral glucose phosphorylation in rainbow trout (Oncorhynchus mykiss) [J]. Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology, 2005, 140(2): 333—340
[77]Enes P, Panserat S, Kaushik S, et al. Rapid metabolic adaptation in European sea bass (Dicentrarchus labrax) juveniles fed different carbohydrate sources after heat shock stress [J]. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 2006, 145(1): 73—81
EFFECTS OF DIETARY CARBOHYDRATE ON GROWTH PERFORMANCE AND GLYCOMETABOLISM OF LARGE YELLOW CROAKER LARIMICHTHYS CROCEA
XING Shu-Juan1, SUN Rui-Jian2, MA Jun1, WEI Hai-Ming1, XU Wei1, ZHOU Hui-Hui1, ZHANG Yan-Jiao1, ZHANG Wen-Bing1and MAI Kang-Sen1
(1. Key Laboratory of Mariculture (Ministry of Education), Key Laboratory of Aquaculture Nutrition and Feeds (Ministry of Agriculture), Ocean University of China, Qingdao 266003, China; 2. Tongwei Technology Center Limited by Share Ltd., Chengdu 610041, China)
To evaluate the effects of dietary carbohydrate on growth performance, feed utilization, plasma parameters, and glycometabolism enzyme activities of large yellow croaker Larimichthys crocea, triplicate groups of fish [the initial body weight: (137.5±0.35) g] were randomly fed six isonitrogenous (45% crude protein) and isoenergetic (18 kJ/g gross energy) diets containing graded levels of carbohydrate (1.75%, 6.67%, 13.64%, 21.15%, 26.69% and 32.25%) in floating sea cages for an 8-week feeding trial. Results showed that the highest specific growth rate (SGR) was by 26.69% of dietary carbohydrate. The highest dietary carbohydrate content (32.25%) significantly decreased the SGR (P<0.05). Both feed efficiency ratio (FER) and protein efficiency ratio (PER) of large yellow croaker fed diets with 13.64% and 21.15% of carbohydrate were significantly higher than those of other groups (P<0.05). The whole-body lipid content significantly decreased the increased dietary carbohydrate contents with the lowest lipid content 32.25% of carbohydrate group (P<0.05). Hepatosomatic index (HSI) and liver glycogen content significantly increased with the increased dietary carbohydrate contents (P<0.05). Plasma total cholesterol and triglyceride levels significantly decreased with the increased dietary carbohydrate level (P<0.05), but plasma glucose did not impact by dietary carbohydrate level (P>0.05). The increased dietary carbohydrate significantly depressed activities of serum lysozyme, lipoprotein lipase and hepatic lipase (P<0.05). The activities of intestinal tract amylase and hepatic hexokinase (HK) increased significantly and then decreased with the highest value at 26.69% and 21.15% dietary carbohydrate contents, respectively (P<0.05). Dietary carbohydrate significantly up-regulated activities of pyruvate kinase (PK) with the peak level by 32.25% carbohydrate group (P<0.05). These results indicated the requirement of dietary carbohydrate for large yellow croaker was 22.7%.
Large yellow croaker; Larimichthys crocea; Carbohydrate; Nutrition; Feed
S963
A
1000-3207(2017)02-0265-12
10.7541/2017.33
2016-03-02;
2016-08-14
國家重點基礎研究發展計劃(“973”計劃)項目(2014CB138600); 廣東恒興水產動物營養與飼料院士工作站項目(2014B09090514)資助 [Supported by the National Basic Research Program (“973” Program, 2014CB138600); Academican Workstation for Aquatic Animal Nutrition and Feed in Guangdong Evergreen (2014B09090514)]
邢淑娟(1989—), 女, 貴州銅仁人; 碩士; 研究方向為水產動物營養與飼料。E-mail: shujuanxing0504@163.com
張文兵, 教授; E-mail: wzhang@ouc.edu.cn
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
火花(2019年12期)2019-12-26 01:00:28
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
人大建設(2019年12期)2019-05-21 02:55:32
汽車工程學報(2017年2期)2017-07-05 08:13:02
學苑創造·A版(2015年11期)2016-01-14 09:03:27
