浙江飲用水源地浮游動物體積多樣性及水質響應
2017-04-12 09:49:45徐杭英于海燕俞建韓明春徐智煥李共國黃
水生生物學報 2017年2期
關鍵詞:營養
徐杭英于海燕俞 建韓明春徐智煥李共國黃 藝
(1. 浙江省環境監測中心, 杭州 310000; 2. 浙江萬里學院, 浙江省生物工程重中之重學科, 寧波 315100; 3. 北京大學環境科學與工程學院, 北京 100871)
浙江飲用水源地浮游動物體積多樣性及水質響應
徐杭英1,3于海燕1俞 建1韓明春1徐智煥2李共國2黃 藝3
(1. 浙江省環境監測中心, 杭州 310000; 2. 浙江萬里學院, 浙江省生物工程重中之重學科, 寧波 315100; 3. 北京大學環境科學與工程學院, 北京 100871)
為了解浙江飲用水源地浮游動物體積多樣性及對水體綜合營養狀態(TLIc值)的響應特征, 對16個飲用水源地(H1-H8為河網型, K1-K8為水庫型)展開了水質參數、浮游動物群落生物體積及其多樣性指數的季節調查。結果表明, 生物體積密度的季節變化以1月(0.681 mL/L)>4月(0.443 mL/L)>10月(0.252 mL/L)>7月(0.229 mL/ L), 各季的第一優勢種分別為湯匙華哲水蚤(Sinocalanus dorrii)(1月)、晶囊輪蟲(Asplanchna sp.)(4月)、粗壯溫劍水蚤(Thermocyclops dybowskii)(7月)和長額象鼻溞(Bosmina longirostris)(10月)。河網浮游動物生物體積以輪蟲(44.6%)和枝角類(23.4%)為主, 水庫則以哲水蚤(39.5%)和輪蟲(29.0%)為主。夏秋季浮游動物體積多樣性指數(P<0.01)和物種多樣性指數(P<0.05)與lg(浮游動物/浮游植物生物量比率)[lg (ZB∶PB)]之間均有顯著的相關性, 水庫中僅浮游甲殼動物體積多樣性指數與lg (ZB∶PB)之間有極其緊密的相關性(R2=0.77)。與物種多樣性指數響應環境效應相比, 甲殼動物的體積多樣性指數(P<0.01)能更好地指示夏秋季河網的TLIc值, 這歸因于甲殼動物個體數(ind./L)轉換成了生物體積密度(mL/L)過程中有效信息的放大。
河網; 水庫; 浮游動物; 體積多樣性指數; TLIc值
生物多樣性的生態系統功能一直是生態學研究的熱點, 由于微型生物群落對環境的敏感變化是有機體連接許多生態系統的基本特征之一, 利用浮游動物生物多樣性評價水體環境質量引起了一些研究者的興趣[1,2], 在營養水平較高的水域, 浮游動物群落多樣性與水體營養水平呈現相反的變化趨勢[3—5], 在營養水平較低的水域, 它們兩者之間則呈現相同的變化趨勢[6]。實際上, 浮游動物群落會受到許多環境因子如溫度、光周期、溶解營養物、水流速快慢等的影響, 這種群落內正常的季節性或日變化與人類活動引起的污染效果還難以區分[7]。因此, 利用浮游動物群落多樣性評價水質還存在諸多限制因子。近年來, 生物體積已被廣泛認為是一個決定生態系統群落結構的關鍵因素, 因為它會影響系統內食物網的結構、穩定性和動態, 浮游動物體積多樣性與水體中懸浮物的過濾效率、抑制藻類生長和水體透明度等密切相關[8—10]。因此, 研究浮游動物體積多樣性及其對水體營養狀態的響應特征, 對促進水質生態學監測工作具有重要意義。
浙江飲用水水庫占全省51%城鄉飲用水源地,供水人口達62%, 但庫區污染源和人類活動已對水庫水源地水質安全構成威脅, 溪、江、河流型飲用水源地的水質更易受到富營養化和突發污染事件的影響[11]。為了解浙江飲用水源地浮游動物體積多樣性及對水質的響應, 本文于2011年1月至10月,對全省兩類水體16個飲用水源地展開了水質理化參數、浮游動物群落優勢種和體積多樣性的季節調查, 探討群落體積多樣性與水體營養水平之間的關系, 以期為浮游動物群落監測水質奠定基礎。
1 材料與方法
1.1 站位布設與采樣方法
根據浙江省飲用水源地主要由大江、大溪、河流和水庫等組成特點, 將由大江、大溪、河流等流速較大的水源地歸為河網型, 選取8個采樣站(H1—H8); 水流速較緩的水庫(型)也選取8個采樣站(K1—K8), 16個浮游動物采樣站分布見圖 1。于2011年的1月、4月、7月和10月, 由全省各地方監測站定點、定時(上午9:00—11:00時)分別采樣與送樣, 采樣和計數按《淡水浮游生物研究方法》[12]進行, 定量樣品用2.5 L采水器分別于該樣站上、下水層, 各采10 L水樣, 用25號篩絹制成的浮游生物網(網孔孔徑64 μm)過濾濃縮, 當即用5%甲醛溶液固定, 帶回實驗室鏡檢。同時測定水體pH (玻璃電極法GB/T6920-1986)、DO (電化學探頭法HJ506-2009)、TN (堿性過硫酸鉀消解紫外分光光度法HJ636-2012)、TP (鉬酸銨分光光度法GB/T11893-1989)、(納氏試劑分光光度法HJ535-2009)和Chl.a (丙酮紫外分光光度法比色法)等水質參數。
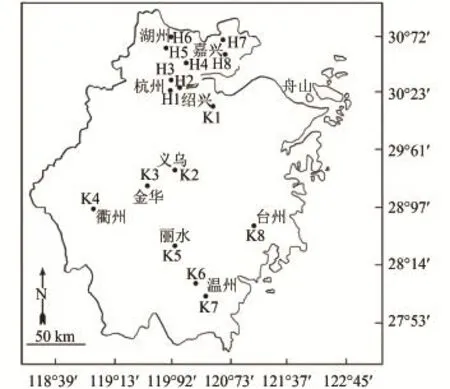
圖 1 浙江飲用水源地浮游動物采樣點分布Fig. 1 Localization of sampling stations for zooplankton from the drinking water sources in Zhejiang
1.2 標本鑒定與計算方法
輪蟲、枝角類和橈足類分別按王家楫[13]、蔣燮治和堵南山[14]、沈嘉瑞[15]鑒定。浮游動物的體長、寬度和厚度采用(顯微鏡)目鏡測微尺測量, 生物體積主要參照章宗涉和黃祥飛[12]、陳雪梅[16]的體積法和體長-體重回歸方程法計算。橈足幼體和成體數量按該種橈足類成體或幼體出現頻率乘以樣本總數計算。群落物種多樣性指數按Shannon-Wiener方法[3]統計; 體積多樣性(Size diversity)指數計算是基于Shannon-Wiener多樣性的表達方式[9,10],即將每種浮游動物的不同體積作為連續變量, 而并非僅僅考慮某一物種的種群數量。為便于計算, 將浮游動物體長的連續變量以每增長200 μm作為一個體積等級分別統計其體積密度, 然后將群落內各種浮游動物不同體積等級的密度分布集合起來, 統計成該群落的體積多樣性。同時, 以Chl.a濃度代替浮游植物生物量[17], 計算浮游生物的營養結構[lg(浮游動物/浮游植物生物量比率)][9], 記作lg (ZB∶PB); 計算水體綜合營養狀態指數(Comprehensive of trophic level index, TLIc)[18]。
1.3 統計分析
以lg (ZB: PB)、TLIc為因變量, 以群落物種或體積多樣性指數為自變量的相關回歸分析應用DPS數據處理系統[19]完成。
2 結果
2.1 飲用水源地理化參數
在16個飲用水源地中, 以河網型的H7、H8站TN、TP和含量為最高(表 1), 均顯著高于其他各站, 其中, 尤以河網各站的TP含量顯著高于水庫各站(P<0.05); 而H8站的DO含量顯著低于水庫各站(P<0.05)。pH以H5、H6和K8站較高, Chl.a含量以K7站最高。河網水體綜合營養指數(TLIc)顯著高于水庫(P<0.05)。相關回歸分析表明: 飲用水源地TLIc值分別與TP、和TN含量均呈極顯著的正相關性, 與DO含量呈極顯著的負相關性(P<0.01)。
2.2 浮游動物生物體積的組成及其季節變化
浮游動物體積密度的組成, 1月以哲水蚤0.391 mL/ L(占57.1%)為主, 主要為水庫中的湯匙華哲水蚤(Sinocalanus dorrii)大量繁殖, K4、K6和K7站分別達2.411、0.589和1.256 mL/L(圖 2); 4月則以輪蟲0.346 mL/L(占78.1%)為主, H2、K5站萼花臂尾輪蟲(Brachionus calyciflorus)體積密度分別達1.394和0.655 mL/L, K1站晶囊輪蟲(Asplanchna sp.)生物體積密度為1.350 mL/L; 7月份的枝角類和劍水蚤體積密度分別達0.083(占36.2%)和0.075 mL/L(占32.8%), 河網以H4站廣布中劍水蚤(Mesocyclops leuckarti)(0.188 mL/L)、H6站長額象鼻溞(Bosmina longirostris)(0.288 mL/L)較高, 水庫以K5站的臺灣溫劍水蚤(Thermocyclops taihokuensis)(0.182 mL/L)和K6站的角突臂尾輪蟲(Brachionus angularis) (0.251 mL/L)的體積密度較高; 10月份的劍水蚤和枝角類體積密度分別達0.116 (占46.0%)和0.081 mL/ L (占32.1%), 其中H5站的廣布中劍水蚤和長額象鼻溞的體積密度分別為0.232和0.420 mL/L, H8站的粗壯溫劍水蚤(Thermocyclops dybowskii)、廣布中劍水蚤和微型裸腹溞(Moina micrura)的體積密度分別為0.499、0.299和0.235 mL/L, 水庫以K4站的粗壯溫劍水蚤(0.141 mL/L)較高。

表 1 2011年飲用水源地水質參數(平均值±標準差)Tab. 1 Water quality parameters in the sampling stations of the drinking water sources during 2011 (mean±SD)
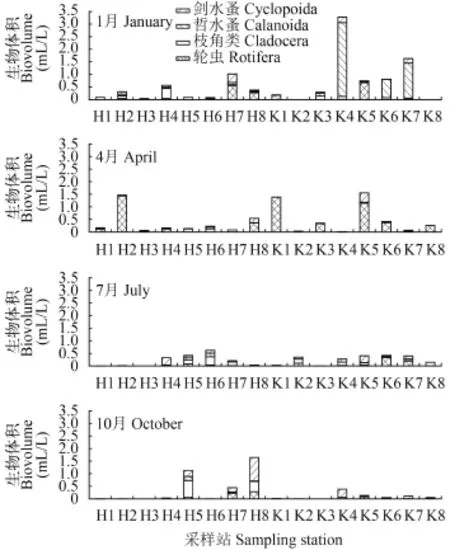
圖 2 各類浮游動物生物體積的季節變化Fig. 2 The seasonal biovolume of each group of zooplankton in the drinking water sources
總浮游動物體積密度的季節變化, 以1月(0.681 mL/L)>4月(0.443 mL/L)>10月(0.252 mL/L)>7月(0.229 mL/L)。水庫1月份浮游動物平均體積密度最高, 達1.169 mL/L; 10月份最低, 僅0.108 mL/L。
2.3 浮游動物體積多樣性及其與lg (ZB∶PB)之間的關系
飲用水源地浮游動物體積多樣性指數平均值1.88, 河網(2.02)>水庫(1.74), 除H8 (2.87)站顯著大于K8 (1.15)(P<0.05)站外, 其余各站之間無顯著差異;甲殼動物體積多樣性指數平均值1.66, 河網(1.78)>水庫(1.55), 除H8 (2.38)站顯著大于K3 (0.84)(P<0.05)站外, 其余各站之間無顯著差異。輪蟲體積多樣性指數平均值1.10, 河網(1.21)>水庫(1.00), 除H7 (2.13)站顯著大于H1—H3、K1 (0.35—0.72)(P<0.05)站外, 其余各站之間無顯著差異。相關分析表明, 夏秋季浮游動物多樣性指數與lg (ZB∶PB)之間呈極顯著(體積多樣性指數, P<0.01)或顯著(物種多樣性指數, P<0.05)的正相關性, 并以甲殼動物體積多樣性指數與lg (ZB∶PB)之間相關性最緊密, 相關系數達0.803 (圖 3); 而冬春季兩者之間則無明顯的相關性。
水庫甲殼動物體積多樣性指數與lg (ZB∶PB)之間相關性比河網更緊密, 相關系數達0.877, 而河網輪蟲體積多樣性指數與lg (ZB∶PB)之間的相關性比水庫更緊密, 相關系數達0.763(圖 4)。
2.4 浮游動物體積多樣性與環境之間的關系
河網夏秋季水體綜合營養指數(TLIc)與浮游動物體積多樣性指數(n=16, r=0.737, P<0.01)和物種多樣性指數(n=16, r=0.635, P<0.05)之間均呈顯著的正相關性。其中, TLIc值與甲殼動物的體積多樣性指數(P<0.01)之間相關性比物種多樣性指數(P<0.05)之間的相關性更緊密(圖 5), 而輪蟲物種多樣性指數與TLIc值之間的相關性更好。水庫夏秋季TLIc值與該兩種浮游動物多樣性指數之間均無相關性(P>0.05)。

圖 3 夏秋季飲用水源地浮游動物多樣性指數與浮游動/植物生物量比率之間的關系Fig. 3 Relationships between biomass ratio of lg (ZB∶PB) and size or species diversities of zooplankton in the drinking water sources during summer and autumn
3 討論
3.1 浮游動物生物體積特征
在貧營養水體中, 浮游動物生物體積通常由體型大的哲水蚤和枝角類占優勢, 而在富營養水體中則由較小的劍水蚤和輪蟲占優勢[20], 飲用水源地河網浮游動物生物量以輪蟲(44.6%)和枝角類(23.4%)為主, 水庫則以哲水蚤(39.5%)和輪蟲(29.0%)為主。各季節的第一優勢種, 1月為湯匙華哲水蚤(水庫), 4月晶囊輪蟲(水庫), 7月長額象鼻溞(河網), 10月粗壯溫劍水蚤(河網)。另外, 飲用水源地一些具富營養化指示功能的數量優勢種, 如萼花臂尾輪蟲、短尾秀體溞(Diaphanosoma brachyurum)、網紋溞(Ceriodaphnia)、長額象鼻溞、裸腹溞(Moina)等[21], 也是浮游動物生物體積(特別是河網)的重要組成者(圖 2), 它們與淺的棲息水體、高濃度的磷和人類活動密切相關[22], 受有毒微囊藻的影響也較小[23]。
另外, 浮游動物體積密度的季節變化還受到了魚類捕食強度的影響, 如K3站(部分水域)2010年底首次開展漁業資源增養殖放流, 投放約9000 kg濾食性的花白鰱后, 在還是魚苗的冬春季, 浮游動物的體積密度較大(0.375 mL/L); 隨著魚類的生長, 捕食強度不斷增大, 夏秋季浮游動物的體積密度僅0.003 mL/L, 哲水蚤種群已消失。
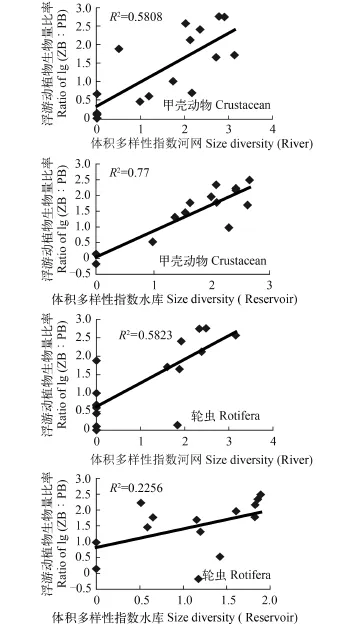
圖 4 夏秋季河網、水庫浮游動物體積多樣性與浮游動/植物生物量比率之間的關系Fig. 4 Relationships between biomass ratio of lg (ZB∶PB) and size diversities of zooplankton in the rivers and reservoirs during summer-autumn

圖 5 河網夏秋季浮游動物多樣性與水體綜合營養狀態指數之間的關系Fig. 5 Relationships between water body TLIc values and size diversities or species diversities of zooplankton in the rivers during summer-autumn
3.2 浮游動物體積多樣性對浮游生物營養結構的影響
在溫帶低營養水平湖泊中, 甲殼動物對葉綠素a牧食率隨著溞屬(Daphian)和枝角類生物量的增大而增大[24]。Ye等[9]假設了浮游動物的每個體積等級都有適合自己攝食的浮游植物最佳尺寸, 并可通過優化的浮游動物/浮游植物生物量比率, 即lg (ZB∶PB)作為“抑藻強度”描述之, 提高浮游動物平均體長或浮游動物體積多樣性(兩者之間呈顯著的正相關性)均可增強對浮游植物的抑制強度, 從而使浮游動物體積多樣性成為決定東海“自上而下”控制浮游植物的最重要因素。在計算體積多樣性指數過程中, 個體數(ind./L)被轉換成了體積密度(mL/L),這放大了大型浮游動物(如溞類、橈足類)“過濾效能”的有效信息, 并使夏秋季飲用水源地浮游動物的“抑藻強度”以體積多樣性指數的指示性較物種多樣性指數更好(圖 3)。河網的輪蟲、甲殼浮游動物體積多樣性與“抑藻強度”均有顯著的正相關性,而水庫僅表現為浮游甲殼動物(圖 4), 這可能與后者在水庫生存竟爭上更勝于輪蟲[25], 以及進入水庫(流域面積小)的外源有機物較河網(流域面積大)少[26], 甲殼動物的營養只能以牧食鏈為主等有關。
3.3 浮游動物體積多樣性指數對水體富營養化的響應特征
河網TLIc值顯著高于水庫(表 1), 但該兩種飲用水源地的浮游動物體積多樣性指數差異并不顯著。可見, 在不同類型的水體中, 不宜用浮游動物體積多樣性指數來指示水質。在水質較好的千島湖輪蟲、枝角類多樣性與營養水平呈現相同的變化趨勢[6], 認為千島湖的營養水平還沒有達到影響浮游動物群落多樣性的程度。同樣的情形也發生在飲用水源地, 河網夏秋季的TLIc值隨著輪蟲、甲殼動物多樣性指數的增大而增大(圖 5)。在生態系統中碎屑食物鏈對物質和能量流動中起著重要的作用, 林群等[27]研究發現, 長江口及毗鄰水域生態系統中通過碎屑鏈傳遞的能流達47%, 并在三峽工程蓄水3年后下降到41%。寧加佳等[26]發現一個由兩條較大河流形成的水庫顆粒有機物與浮游動物的碳、氮穩定同位素之間有相似的變化趨勢, 認為雨季浮游動物的食物來源主要來自外源有機物。因此, 流域面積大的河網水能通過降雨(特別是夏秋季)接納更多來自流域內的有機物, 致使水體中的有機物以外源性為主, 甲殼動物和輪蟲的營養獲得除牧食食物鏈外, 還有較多的有機碎屑鏈補充,在溫度較高的夏秋季, 浮游動物生長繁殖快, 對水體中營養物質的利用度更高, 這使浮游動物體積多樣性與TLIc值呈現顯著的正相關性。在鏡檢過程中, 河網采樣站發現有較多的有機碎屑。而水庫由于其流域面積遠小于河網型, 外源有機物進入少,且溫帶低營養水平中甲殼動物對Chl.a的牧食率較低, 可利用藻類(體長<30 μm)牧食率隨著Chl.a濃度的增大而下降[23]。因此, 水庫浮游動物的食物顯得較為貧乏, 這可能是夏秋季水庫浮游動物體積多樣性指數與TLIc值無顯著相關性的主要原因。
與物種多樣性相比較, 體積多樣性包涵了物種不同體積分布的信息, 對于體型大、生長過程中又經歷了多個不同發育期的浮游甲殼動物, 通過統計體積多樣性更能反映其對水體營養物質的利用、轉化與積累, 從而貯存更多的信息。這可能是夏秋季河網水體TLIc值與甲殼動物體積多樣性指數(P<0.01)之間的相關性較物種多樣性指數(P<0.05)更緊密的原因(圖 5), 而個體小的輪蟲在統計體積多樣性的過程中, 會因體積數值小而舍棄部分物種信息,導致輪蟲體積多樣性與水體TLIc值的相關性(P>0.05)反而不如物種多樣性(P<0.05)。可見, 浮游動物營養物來源和多樣性指數計算方式的不同, 決定了群落對飲用水源地營養水平的響應特征。
綜上所述, 不同營養水體、季節變化條件下浮游動物生物多樣性存在較大的差異。夏秋季河網和水庫中浮游生物營養結構主要受到浮游動物體積多樣性的影響, 響應夏秋季河網水體富營養最敏感的是甲殼動物體積多樣性指數。冬春季飲用水源地、夏秋季水庫的水體營養狀態對浮游動物生物多樣性的影響均不顯著, 可推測其營養水平較低、且波動不大, 但還需要進一步的論證。因此, 應尋求一種更為靈敏的生物多樣性指數或其他拓展性指標, 以適應于更寬闊生態范圍內的水質監測。
[1]Vandysh O I. Zooplankton as an indicator of the state of lake ecosystems polluted with mining wastewater in the Kola Peninsula [J]. Russian Journal of Ecology, 2004, 35(2): 110—116
[2]Paturej E, Go?dziejewska A. Zooplankton-based assessment of the trophic state of three coastal lakes - ?ebsko, Gardno, and Jamno [J]. The Bulletin of the Fisheries Institute, 2005, 166(3): 7—25
[3]Li G G, Bao W H , Xu S L, et al. Seasonal change of zooplankton communities and its relationship with aquatic environments in the Yongjiang River, Ningbo [J]. Acta Hydrobiologica Sinica, 2015, 39(1): 1—12 [李共國, 包薇紅, 徐石林, 等. 甬江干流浮游動物群落結構季節動態與水環境的關系. 水生生物學報, 2015, 39(1): 1—12]
[4]Zeng Y, Fu X E, Miao M S, et al. Water quality assessment of Wenyuhe River based on the cross-correlation analysis on the diversity of macro-zooplankton and water parameters [J]. Asian Journal of Ecotoxicology, 2012, 2(7): 162—170 [曾陽, 付秀娥, 苗明升, 等. 基于大型浮游動物多樣性與水質參數相關性分析的溫榆河水質評價. 生態毒理學報, 2012, 2(7): 162—170]
[5]Ji Y, Zhang J, Ma X, et al. Space-time distribution characteristics of zooplankton and water quality assessment in the Fuhe River [J]. Journal of Henan Agricultural Sciences, 2013, 42(3): 66—70 [計勇, 張潔, 麻夏, 等. 撫河干流浮游動物生物多樣性調查及水質評價. 河南農業科學, 2013, 42(3): 66—70]
[6]Li G G, Yu Z M. Community structure of zooplankton in Lake Qiandaohu [J]. Acta Ecologica Sinica, 2002, 22(2): 156—162 [李共國, 虞左明. 千島湖浮游動物群落結構研究. 生態學報, 2002, 22(2): 156—162]
[7]Guo P Y, Lin Y Z. Application of micro-biological monitoring of water pollution [J]. Shandong Environment, 1998, 82(1): 19—20 [郭沛涌, 林育真. 應用微型生物監測水質污染, 山東環境, 1998, 82(1): 19—20]
[8]Ordonez J, Armengol J, Moreno-Ostos E, et al. On non-Eltonian methods of hunting Cladocera, or impacts of the introduction of planktivorous fish on zooplankton composition and clear-water phase occurrence in a Mediterranean reservoir [J]. Hydrobiologia, 2010, 653(1): 119—129
[9]Ye L, Chang C Y, Carmen G C, et al. Increasing zooplankton size diversity enhances the strength of top-down control on phytoplankton through diet niche partitioning [J]. Journal of Animal Ecology, 2013, 82(5): 1052—1061
[10]Quintana X D, Arim M, Badosa A, et al. Predation and competition effects on the size diversity of aquatic communities [J]. Aquatic Sciences, 2015, 77(1): 45—57
[11]Wang D Y, Zhang Y. Statistical analysis on drinking water source and supply system contamination threats and incidents for urban areas in China during 2006 [J]. Journal of Safety and Environment, 2007, 7(6): 150—155 [王東宇, 張勇. 2006年中國城市飲用水源突發污染事件統計及分析. 安全與環境學報, 2007, 7(6): 150—155]
[12]Zhang Z S, Huang X F. Methods for Study on Freshwater Plankton [M]. Beijing: Science Press. 1991, 358—371 [章宗涉, 黃祥飛. 淡水浮游生物研究方法. 北京: 科學出版社. 1991, 358—371]
[13]Wang J J. Fauna Sinica of Chinese Freshwater Rotifera [M]. Beijing: Science Press. 1961, 69—254 [王家楫. 中國淡水輪蟲志. 北京: 科學出版社. 1961, 69—254]
[14]Jiang X Z, Du N S. Fauna Sinica, Crustacea, Freshwater Cladoceran [M]. Beijing: Science Press. 1979, 86—171 [蔣燮治, 堵南山. 中國動物志(淡水枝角類). 北京: 科學出版社. 1979, 86—171]
[15]Fauna Editorial Committee, Academia Sinica. Fauna Sinica, Crustacea, Freshwater Copepoda [M]. Beijing: Science Press. 1979, 60—420 [中國科學院動物研究所甲殼動物研究組編. 中國動物志(淡水橈足類) . 北京: 科學出版社. 1979, 60—420]
[16]Chen X M. Biomass calculation of freshwater Copepoda [J]. Acta Hydrobiologica Sinica, 1981, 7(3): 397—408 [陳雪梅. 淡水橈足類生物量的測算. 水生生物學集刊, 1981, 7(3): 397—408]
[17]Brucet S, Boix D, Lopez-Flores R, et al. Size and species diversity of zooplankton communities in fluctuating Mediterranean salt marshes [J]. Estuarine, Coastal and Shelf Science, 2006, 67(3): 424—432
[18]Wang M C, Liu X Q, Zhang J H. Evaluate method and classification standard on lake eutrophication [J]. Environmental Monitoring in China, 2002, 18(5): 47—49 [王明翠, 劉雪芹, 張建輝. 湖泊富營養化評價方法及分級標準. 中國環境監測, 2002, 18(5): 47—49]
[19]Tang Q Y, Feng M G. DPS Data Processing System-Experimental Design, Statistical Analysis and Data Mining [M]. Beijing: Science Press. 2002, 636—644 [唐啟義, 馮明光. DPS數據處理系統-實驗設計、統計分析及數據挖掘. 北京: 科學出版社. 2002, 636—644]
[20]Wang X H, Wang T, Lin Q Q, et al. Specific composition and structure of meta-zooplankton community in a pumbped storage reservoir, South China [J]. Acta Ecologica Sinica, 2009, 29(1): 456—465 [王曉輝, 望甜, 林秋奇, 等. 華南地區典型抽水型水庫后生浮游動物群落的種類組成與結構. 生態學報, 2009, 29(1): 456—465]
[21]Xu H Y, Yu H Y, Han M C, et al. Characteristics of zooplankton community and their association with environmental factors in the drinking water sources, Zhejiang Province, China [J]. Acta Ecologica Sinica, 2015, 35(21): 7219—7228 [徐杭英, 于海燕, 韓明春, 等. 浙江飲用水源地浮游動物群落特征及環境響應. 生態學報, 2015, 35(21): 7219—7228]
[22]Van Egeren S J, Dodson S I, Torke B, et al. The relative significance of environmental and anthropogenic factors affecting zooplankton community structure in Southeast Wisconsin Till Plain lakes [J]. Hydrobiologia, 2011, 668(1): 137—146
[23]Zhu J Y, Lu K H, Pan J H. Progress in the effect of Microcystis bloom on freshwater zooplankton rotifer and cladoceran [J]. Journal of Fishery Sciences of China, 2008, 15(2): 367—375 [朱津永, 陸開宏, 潘潔慧. 微囊藻水華對淡水浮游動物輪蟲和枝角類影響的研究進展.中國水產科學, 2008, 15(2): 367—375]
[24]Symons C C, Arnott S E, Sweetman J N. Grazing rates of crustacean zooplankton communities on intact phytoplankton communities in Canadian Subarctic lakes and ponds [J]. Hydrobiologia, 2012, 694(1): 131—141
[25]Luis M B, Leo C F, Luiz F M. Zooplankton assemblage concordance patterns in Brazilian reservoirs [J]. Hydrobiologia, 2008, 598(1): 247—255
[26]Ning J J, Liu H, Gu B H, et al. Carbon and nitrogen stable isotope characteristics of particulate organic matter and zooplankton in Liuxihe Reservoir [J]. Acta Ecologica Sinica, 2012, 32(5): 1502—1509 [寧加佳, 劉輝, 古濱河,等. 流溪河水庫顆粒有機物及浮游動物碳、氮穩定同位素特征. 生態學報, 2012, 32(5): 1502—1509]
[27]Lin Q, Jin X S, Guo X W, et al. Study on the structure and energy flow of theYangtze River Estuary and adjacent waters ecosystem based on ecopath model [J]. Journal of Hydroecology, 2009, 2(2): 28—36 [林群, 金顯仕,郭學武, 等. 基于Ecopath模型的長江口及毗鄰水域生態系統結構和能量流動研究. 水生態學雜志, 2009, 2(2): 28—36]
A PRELIMINARY STUDY ON SIZE DIVERSITY OF ZOOPLANKTON AND THEIR ASSOCIATION WITH TROPHIC LEVEL IN THE DRINKING WATER SOURCES, ZHEJIANG PROVINCE
XU Hang-Ying1,3, YU Hai-Yan1, YU Jian1, HAN Ming-Chun1, XU Zhi-Huan2, LI Gong-Guo2and HUANG Yi3
(1. Zhejiang Environment Monitoring Center, Hangzhou 310000, China; 2. Zhejiang Provincial Top Key Discipline of Biological Engineering, Zhejiang Wanli University, Ningbo 315100, China; 3. College of Environmental Sciences and Engineering, Peking University, Beijing 100871, China)
In order to understand the size diversities of the zooplankton community and response characteristics of the comprehensive water bodies by trophic level index (TLIc value), a seasonal investigation was conducted on zooplankton size diversities from two types of 16 drinking water sources (H1-H8 for rivers, K1-K8 for reservoirs) from January to October 2011 in Zhejiang Province. The results showed that seasonal variations in the biovolume density of zooplankton from high to low were January (0.681 mL/L), April (0.443 mL/L), October (0.252 mL/L), and July (0.229 mL/L). The first dominant species of January, April, July, and October were Sinocalanus dorrii, Asplanchna sp., Thermocyclops dybowskii, and Bosmina longirostris respectively. Zooplankton were mainly Rotifera (44.6%) and Cladocera (23.4%) in the rivers by their biological volume, while Calanoida (39.5%) and Rotifera (29.0%) in the reservoirs. There were significant relationships between lg 10 transformed zooplankton/phytoplankton biomass ratio [lg (ZB∶PB)] and size diversities (P<0.01) or species diversities (P<0.05) of zooplankton in the drinking water sources during summer and autumn, and crustacean zooplankton size diversity index was significantly positive correlated with lg (ZB∶PB) (R2=0.77) in the reservoirs. Crustaceans size diversity (P<0.01) were more reliable to TLIc value of the rivers than the species diversity during summer and autumn, supported by the useful information amplification from the numbers of crustaceans (ind./L) converting into biovolume density (mL/L).
Rivers; Reservoirs; Zooplankton; Size diversity index; TLIc value
Q145+.2
A
1000-3207(2017)02-0407-07
10.7541/2017.50
2015-08-03;
2016-06-28
浙江省自然科學基金青年基金(Q13C030015); 浙江省重中之重學科開放基金(KF2016003)資助 [Supported by the Zhejiang Province Science Foundation for Youths (Q13C030015); the Open Foundation of Zhejiang Provincial Top Key Discipline (KF2016003)]
徐杭英(1985—), 女, 浙江杭州人; 碩士研究生, 工程師; 主要從事水域環境保護研究。E-mail: xinxinxhy@163.com
李共國, E-mail: ligongguo@tom.com; 黃藝, E-mail: yhuang@pku.edu.cn
猜你喜歡
中老年保健(2022年5期)2022-11-25 14:16:14
今日農業(2021年11期)2021-08-13 08:53:34
今日農業(2021年4期)2021-06-09 06:59:58
今日農業(2020年24期)2020-03-17 08:58:18
中國生殖健康(2019年8期)2019-01-07 01:18:24
幸福(2018年33期)2018-12-05 05:22:46
廣東第二課堂·小學(2017年2期)2017-02-20 15:10:08
廣東第二課堂·小學(2017年1期)2017-02-20 14:47:45
少兒科學周刊·兒童版(2015年12期)2016-05-16 03:38:08
小學閱讀指南·低年級版(2016年8期)2016-05-14 13:24:45
