短季棉生育期性狀的遺傳分析
2017-04-13 06:56:47田鋒范術麗魏恒玲王寒濤趙樹琪龐朝友胡守林喻樹迅
新疆農業科學 2017年2期
田鋒,范術麗,魏恒玲,王寒濤,趙樹琪,3,龐朝友,胡守林,喻樹迅
(1.塔里木大學植物科學學院,新疆阿拉爾 843300;2.中國農業科學院棉花研究所/棉花生物學國家重點實驗室,河南安陽 455000;3.黃岡市農業科學院,湖北黃岡 438000)
短季棉生育期性狀的遺傳分析
田鋒1,2,范術麗2,魏恒玲2,王寒濤2,趙樹琪2,3,龐朝友2,胡守林1,喻樹迅2
(1.塔里木大學植物科學學院,新疆阿拉爾 843300;2.中國農業科學院棉花研究所/棉花生物學國家重點實驗室,河南安陽 455000;3.黃岡市農業科學院,湖北黃岡 438000)
【目的】研究短季棉生育期的遺傳規律,為棉花早熟性狀QTL定位和分子標記輔助育種提供理論基礎。【方法】利用四個陸地棉品種構建兩個雜交群體,采用主基因+多基因聯合世代分析,對棉花早熟性相關的播種-開花(Flowering time,FT),花鈴期(Flowering and boll-setting period, FBP),全生育期(Whole growing period,WGP)這三個性狀進行遺傳分析。【結果】兩個組合,除組合Ⅱ中F1的花鈴期和F2的播種-開花期偏向晚熟親本外,其他表型性狀均偏向早熟親本,但總體偏早熟親本;組合Ⅰ及組合Ⅱ的相同性狀均符合同一個遺傳模型,且均受兩對主基因控制。【結論】三個表型性狀是由主基因和多個微效基因共同作用,且主要由主基因控制;三個表型性狀的遺傳力高,且加性效應大。
短季棉;生育期;主基因+多基因;遺傳分析
0 引 言
【研究意義】棉花一直是我國重要的經濟作物,棉花種植業的發展關系到紡織工業的興衰,同時也涉及到我國近一億棉農及兩千多萬紡織工人的就業、再就業問題[1-2]。人多地少,糧棉爭地的問題一直存在,這使棉花種植面積一再縮減,導致國內棉花產量無法滿足內需,還需要向外大量進口[3]。緩解糧棉爭地矛盾,解決我國棉花供需以及糧食需求的一個有效方法就是提高糧棉復種指數,實現糧棉由一熟向兩熟發展[4]。因此,選育生育期較短,品種優良的短季棉品種勢在必行[5]。棉花早熟相關性狀是復雜的數量性狀,受到多個數量性狀位點和環境的共同影響,了解棉花關于早熟的遺傳規律,對培育短季棉品種具有的指導意義。【前人研究進展】前人根據數量遺傳學的方法,從整體上分析了某一性狀的多個微效基因的遺傳效應,但無法解析單一基因的遺傳效應[6-7]。在相關棉花早熟性的遺傳分析中,范術麗[8]利用兩個組合的四個世代群體,對棉花早熟相關性狀主基因-多基因的遺傳規律進行了分析,揭示了早熟相關性狀主基因存在的普遍性。董娜[7]根據六個世代,利用了主基因-多基因混合遺傳模型分析方法,說明早熟相關性狀的基因表達很容易受到環境的影響。【本研究切入點】喻樹迅等[9]在對所推廣的早熟棉品種進行的遺傳相關分析中提出,生育期可以作為早熟性的指示性狀。艾先濤等[10]認為,通過品種遺傳多樣性的分析可以減少育種過程中的盲目性與重復性。利用多世代聯合遺傳分析,對棉花早熟性狀進行分析,確定控制早熟性基因的作用方式。【擬解決的關鍵問題】研究采用主基因-多基因混合遺傳模型對組合Ⅰ(中棉所60×中棉所50)和組合Ⅱ(中棉所41×中棉所74)的四個世代群體P1、P2、F1、F2,對棉花生育期三個早熟相關性狀的遺傳規律進行分析,明確棉花早熟性狀的遺傳模型及基因作用方式,估測出兩組合的主基因遺傳效應和遺傳力,為棉花早熟性狀QTL定位及分子標記輔助育種提供理論基礎[10]。
1 材料與方法
1.1 材 料
選用早熟棉品種和中熟棉品種為親本構建兩個分離群體(組合Ⅰ:中棉所60×中棉所50;組合Ⅱ:中棉所41×中棉所74)。2014年在安陽配置兩個雜交組合,獲得F1代種子,同年將獲得的F1代種子在海南進行加代,自交得到F2代種子。2015年5月25日在河南安陽種植兩個雜交組合的親本及其F1各3行,種植組合Ⅰ的F2分離群體414株,種植組合Ⅱ的F2分離群體468株。
1.2 方 法
1.2.1 性狀調查
對兩雜交組合的親本、F1及F2群體進行單株調查。親本及F1代植株各調查30個單株,其F2分離群體單株全部調查。調查性狀包括:播種-開花(Flowering time,FT)、花鈴期(Flowering and boll-setting period, FBP)、全生育期(Whole growing period,WGP)3個表型性狀。
1.2.2 遺傳模型及參數估計
采用章元明等[11]提出的利用P1、P2、F1、F2四個世代的主基因+多基因混合遺傳分析法對單株表型數據進行分析。根據所計算出的各遺傳模型AIC值(Akaike's Information Criterion,AIC)來確定候選基因型,進一步對候選基因模型進行均勻性檢驗、Smirnov檢驗以及Kolmogorov檢驗,最終根據檢驗結果選定最適模型,并對估算出的各項遺傳參數進行分析。
2 結果與分析
2.1 表型性狀特性
研究表明,組合Ⅰ中的母本相對于父本為晚熟品種,并且兩個親本間播種-開花時間相差11.57 d,花鈴期相差13 d,全生育期相差24.57 d;組合Ⅱ的母本為晚熟品種,父本為早熟品種,調查數據顯示,兩個親本間播種-開花相差7.44 d,花鈴期相差8.16 d ,全生育期相差15.6 d;兩個組合親本之間的差異有統計學意義。從表型數據可以看出,組合Ⅰ中F1、F2表型偏向早熟親本中棉所50,由此可以肯定早熟親本的早熟性在后代群體中部分呈顯性遺傳;組合Ⅱ中除F1的花鈴期和F2的播種-開花期偏向晚熟親本中棉所41,其它性狀的都偏向早熟親本,對兩個組合的總體而言,群體表型偏向早熟親本,在后代群體中,早熟性部分呈顯性遺傳。表1
表1 各世代表型特征
2.2 遺傳模型的選擇和適合性檢測
采用四世代聯合分析的方法,根據兩個組合的播種-開花、花鈴期、全生育期的田間表型數據估算出24種遺傳模型的AIC值,并根據最小AIC值的原則在每個指標中選取兩個候選模型。結果顯示兩個組合的播種-開花、全生育期、花鈴期的候選模型全都為E和E-1。通過對以上候選的遺傳模型進行適合性檢驗,結合檢驗結果確定最適遺傳模型。表2
適合性檢驗結果所示,根據兩個組合不同生育期性狀模型的統計量達到的顯著水平及最小AIC的原則,確定組合Ⅰ、組合Ⅱ不同生育期的最適遺傳模型。兩個雜交組合的播種-開花及全生育期的最適遺傳模型均符合模型E(兩對加性-顯性-上位性主基因+加性-顯性-上位性多基因模型);花鈴期的最適遺傳模型也都符合模型E-1(即兩對加性-顯性-上位性主基因+加性-顯性多基因模型)。根據最適模型可以看出,組合Ⅰ及組合Ⅱ各生育期的最適遺傳模型是相一致的,兩個雜交組合在生育期的遺傳性狀穩定,這三個生育期性狀可以作為早熟性的指示性狀。表3
2.3 遺傳參數的估計
利用最小二階乘法分別估計組合Ⅰ及組合Ⅱ播種-開花、花鈴期、全生育期最適遺傳模型的一階參數和二階參數,分析兩個雜交組合各個生育時期的遺傳規律。表4
2.3.1 播種-開花主-多基因的遺傳分析
通過最優遺傳模型E(兩對加性-顯性-上位性主基因+加性-顯性-上位性多基因混合遺傳模型)對兩個組合播種-開花的性狀進行遺傳參數的估計,研究表明,組合Ⅰ、組合Ⅱ控制開花的兩對主基因的加性效應分別為7.174、5.189和3.474、1.402,顯性效應分別為4.684、2.709和-1.172、-1.393,兩個雜交組合的第一對主基因的顯性程度高于第二對主基因的顯性程度;兩對主基因間的上位效應分別為-2.709和-0.884;兩個雜交組合控制播種-開花的主基因的遺傳率分別為63.007%和64.289%,多基因的遺傳率分別為36.993%和35.702%,主基因的遺傳率高于多基因的遺傳率,組合Ⅰ、組合Ⅱ播種-開花性狀主要是由主基因控制。表4
表2 四世代不同模型的AIC值
Table 2 The AIC values of different genetic models
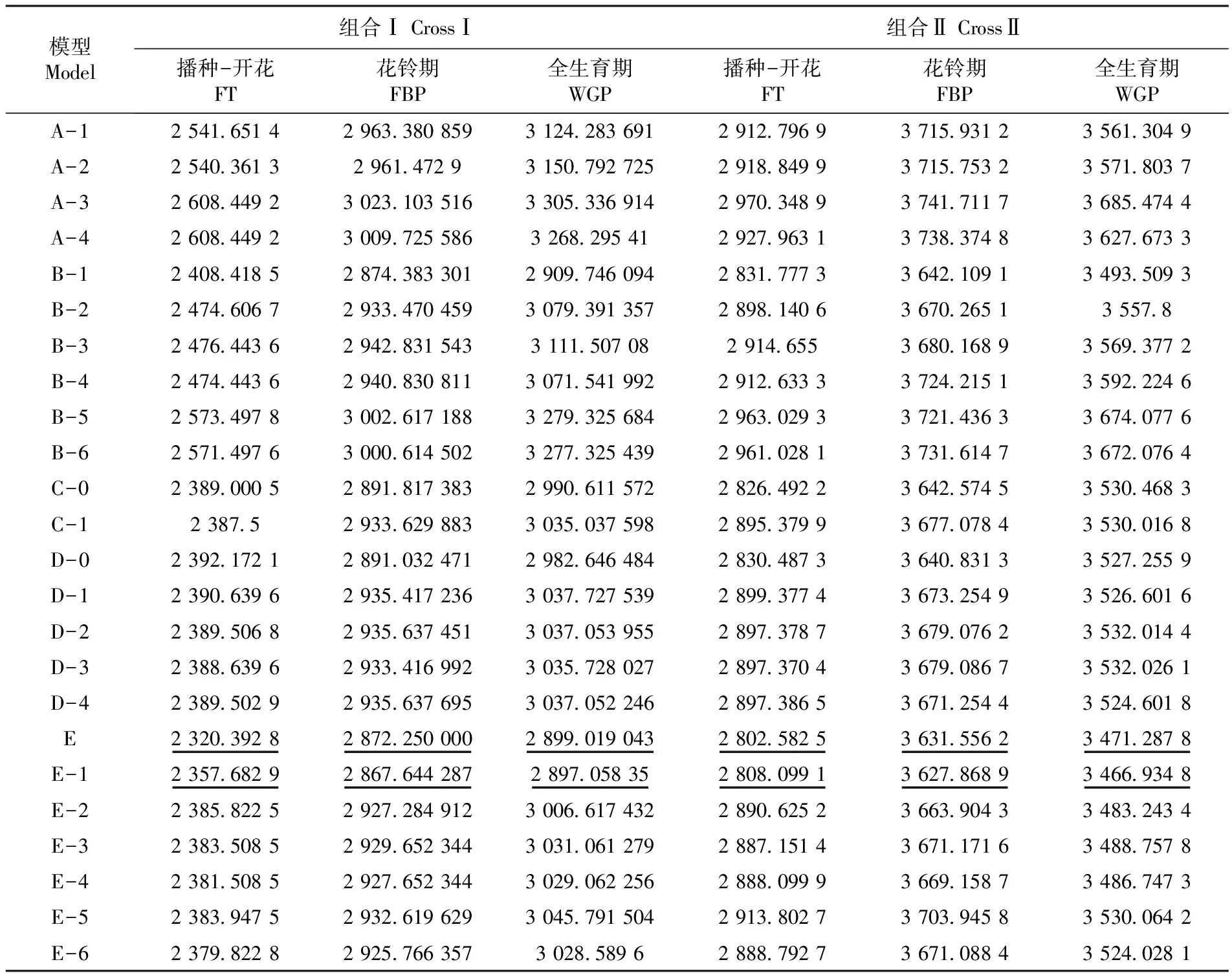
模型Model組合ⅠCrossⅠ組合ⅡCrossⅡ播種-開花FT花鈴期FBP全生育期WGP播種-開花FT花鈴期FBP全生育期WGPA-12541651429633808593124283691291279693715931235613049A-225403613296147293150792725291884993715753235718037A-32608449230231035163305336914297034893741711736854744A-4260844923009725586326829541292796313738374836276733B-12408418528743833012909746094283177733642109134935093B-22474606729334704593079391357289814063670265135578B-324764436294283154331115070829146553680168935693772B-42474443629408308113071541992291263333724215135922246B-52573497830026171883279325684296302933721436336740776B-62571497630006145023277325439296102813731614736720764C-02389000528918173832990611572282649223642574535304683C-12387529336298833035037598289537993677078435300168D-02392172128910324712982646484283048733640831335272559D-12390639629354172363037727539289937743673254935266016D-22389506829356374513037053955289737873679076235320144D-32388639629334169923035728027289737043679086735320261D-42389502929356376953037052246289738653671254435246018E2320392828722500002899019043280258253631556234712878E-1235768292867644287289705835280809913627868934669348E-22385822529272849123006617432289062523663904334832434E-32383508529296523443031061279288715143671171634887578E-42381508529276523443029062256288809993669158734867473E-52383947529326196293045791504291380273703945835300642E-623798228292576635730285896288879273671088435240281
注:“__”表示候選模型
Note: “__”Means candidate model
表3 早熟性狀遺傳模型適合性檢驗
Table 3 Tests for goodness-of-fit about genetic models of earliness traits
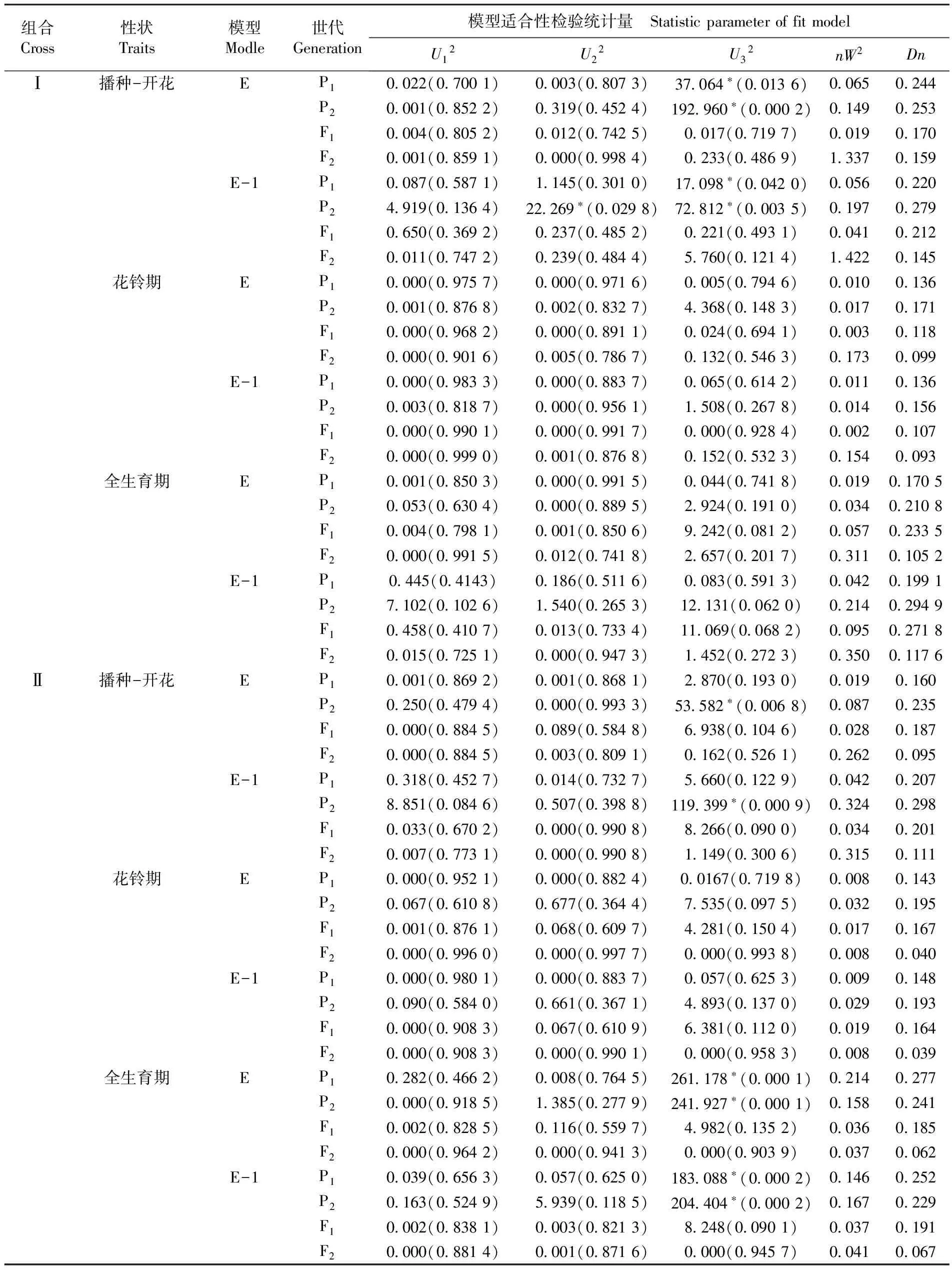
組合Cross性狀Traits模型Modle世代Generation模型適合性檢驗統計量 StatisticparameteroffitmodelU12U22U32nW2DnⅠ播種-開花EP10022(07001)0003(08073)37064?(00136)00650244P20001(08522)0319(04524)192960?(00002)01490253F10004(08052)0012(07425)0017(07197)00190170F20001(08591)0000(09984)0233(04869)13370159E-1P10087(05871)1145(03010)17098?(00420)00560220P24919(01364)22269?(00298)72812?(00035)01970279F10650(03692)0237(04852)0221(04931)00410212F20011(07472)0239(04844)5760(01214)14220145花鈴期EP10000(09757)0000(09716)0005(07946)00100136P20001(08768)0002(08327)4368(01483)00170171F10000(09682)0000(08911)0024(06941)00030118F20000(09016)0005(07867)0132(05463)01730099E-1P10000(09833)0000(08837)0065(06142)00110136P20003(08187)0000(09561)1508(02678)00140156F10000(09901)0000(09917)0000(09284)00020107F20000(09990)0001(08768)0152(05323)01540093全生育期EP10001(08503)0000(09915)0044(07418)001901705P20053(06304)0000(08895)2924(01910)003402108F10004(07981)0001(08506)9242(00812)005702335F20000(09915)0012(07418)2657(02017)031101052E-1P10445(04143)0186(05116)0083(05913)004201991P27102(01026)1540(02653)12131(00620)021402949F10458(04107)0013(07334)11069(00682)009502718F20015(07251)0000(09473)1452(02723)035001176Ⅱ播種-開花EP10001(08692)0001(08681)2870(01930)00190160P20250(04794)0000(09933)53582?(00068)00870235F10000(08845)0089(05848)6938(01046)00280187F20000(08845)0003(08091)0162(05261)02620095E-1P10318(04527)0014(07327)5660(01229)00420207P28851(00846)0507(03988)119399?(00009)03240298F10033(06702)0000(09908)8266(00900)00340201F20007(07731)0000(09908)1149(03006)03150111花鈴期EP10000(09521)0000(08824)00167(07198)00080143P20067(06108)0677(03644)7535(00975)00320195F10001(08761)0068(06097)4281(01504)00170167F20000(09960)0000(09977)0000(09938)00080040E-1P10000(09801)0000(08837)0057(06253)00090148P20090(05840)0661(03671)4893(01370)00290193F10000(09083)0067(06109)6381(01120)00190164F20000(09083)0000(09901)0000(09583)00080039全生育期EP10282(04662)0008(07645)261178?(00001)02140277P20000(09185)1385(02779)241927?(00001)01580241F10002(08285)0116(05597)4982(01352)00360185F20000(09642)0000(09413)0000(09039)00370062E-1P10039(06563)0057(06250)183088?(00002)01460252P20163(05249)5939(01185)204404?(00002)01670229F10002(08381)0003(08213)8248(00901)00370191F20000(08814)0001(08716)0000(09457)00410067
注:*:差異顯著(P<0.05);U12、U22、U32:均勻性檢驗;nW2:Smirnov檢驗;Dn:Kolmogorov 檢驗
Note:*: Significant difference at 0.05 levels;U12,U22andU32: were statistics of uniformity test;nW2was the statistic of Smirnov test;Dnwas the statistic of Kolmogorov test
2.3.2 花鈴期主-多基因的遺傳分析
通過最優遺傳模型E-1(兩對加性-顯性-上位性主基因+加性-顯性多基因混合遺傳模型)對花鈴期這一性狀進行遺傳參數估計,結果可以看出,組合Ⅰ、組合Ⅱ控制花鈴期的兩對主基因的加性效應分別為3.671、0.511和7.473、2.613;顯性效應分別為-0.121、-4.480和-4.149、-4.658,兩個雜交組合控制花鈴期主基因的顯性效應均為負值,控制花鈴期性狀的兩對主基因呈負向不完全顯性;組合Ⅰ控制花鈴期的多基因的加性效應和顯性效應分別為2.318、3.535,組合Ⅱ控制花鈴期的多基因的加性效應和顯性效應分別為-6.003、7.168;兩個雜交組合控制花鈴期主基因的遺傳率分別為67.675%、86.405%;多基因的遺傳率分別為32.325%、13.595%,主基因遺傳率相比多基因遺傳率要高,花鈴期性狀主要是由主基因來控制。表4
2.3.3 全生育期主-多基因的遺傳分析
通過最優遺傳模型E-1(兩對加性-顯性-上位性主基因+加性-顯性多基因混合遺傳模型)對全生育期性狀進行遺傳參數估計,研究表明,組合Ⅰ、組合Ⅱ控制全生育期的兩對主基因的加性效應分別為6.365、3.513和8.460、4.450;顯性效應分別為-0.283、-4.091和-1.000、1.298;組合Ⅰ、組合Ⅱ的兩對主基因間均存在上位性效應,上位性效應分別為3.128和0.932;組合Ⅰ多基因的加性效應和顯性效應分別為2.406、1.687,組合Ⅱ多基因的加性效應和顯性效應分別為-5.109、-1.227;兩個雜交組合主基因的遺傳率分別為60.875%和70.837%;多基因的遺傳率分別為39.125%和29.163%,主基因遺傳率較高,全生育期性狀是由主基因控制。表4
表4 棉花早熟性狀遺傳參數估計
Table 4 The estimates of genetic parameters of fit model of earliness characters
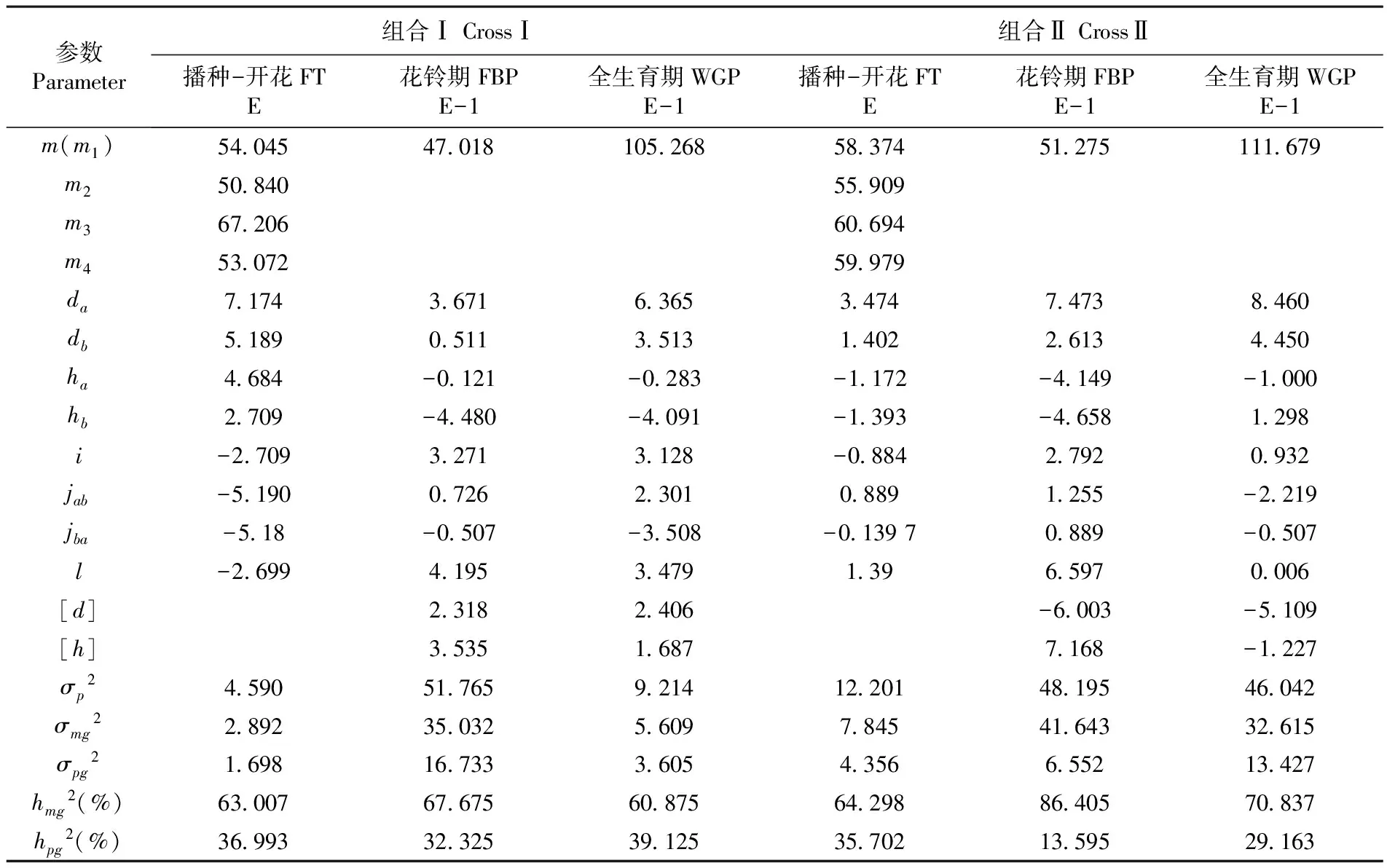
參數Parameter組合ⅠCrossⅠ組合ⅡCrossⅡ播種-開花FTE花鈴期FBPE-1全生育期WGPE-1播種-開花FTE花鈴期FBPE-1全生育期WGPE-1m(m1)54045470181052685837451275111679m25084055909m36720660694m45307259979da717436716365347474738460db518905113513140226134450ha4684-0121-0283-1172-4149-1000hb2709-4480-4091-1393-46581298i-270932713128-088427920932jab-51900726230108891255-2219jba-518-0507-3508-013970889-0507l-26994195347913965970006[d]23182406-6003-5109[h]353516877168-1227σp24590517659214122014819546042σmg2289235032560978454164332615σpg216981673336054356655213427hmg2(%)630076767560875642988640570837hpg2(%)369933232539125357021359529163
注:m、m1、m2、m3、m4:中親值;da、ha:主基因a的加性效應和顯性效應;db、hb:主基因b的加性效應和顯性效應:[d]、[h]:多基因加性效應和顯性效應;i、l:兩個主基因的加×加效應和顯×顯效應;jab:加性(a)×顯性(b)效應;jba:加性(b)×顯性(a)效應;σp2:表型方差;σmg2:主基因方差;σpg2:多基因方差;hmg2:主基因遺傳率;hpg2:多基因遺傳率性
Note:m,m1,m2,m3,m4:Mid-parent value;da、ha: Additive effect and dominant effect of the first pair major gene;db、hb: Additive effect and dominant effect of the second pair major gene; [d]、[h]: Additive effective value and dominant effective value of polygene;i、l: Additive effect plus additive effect and dominant effect plus dominant effect of the two major genes;jab: Additive effect plus dominant effect of the two major genes;jba: Dominant effect plus additive effect of the two major genes;σp2: Phenotypic variance;σmg2: Major gene variance;σpg2: Polygenic variance;hmg2: Major gene heritability;hpg2: Polygene heritability
3 討 論
棉花的早熟性是復雜的數量性狀極易受到環境因素干擾[12-13],研究利用兩個雜交組合的四個世代群體對棉花生育期的三個性狀進行的聯合分析,可以使試驗的精準度有所增加,也可以使主基因遺傳參數方面估計值的精度有所提高,確保了分析結果的準確度和可靠性[14-15]。在前人的研究結果中總結發現,數量性狀是一種混合遺傳模型,是由主基因和多基因共同來控制的,因此,控制數量性狀的基因在其遺傳效應大小上存在較大的差異[16]。棉花的早熟性是由多基因控制的復雜的數量性狀,同時存在加性效應與顯性效應[17-19]。播種-開花、花鈴期、全生育期等性狀是短季棉早熟的重要指標性狀,Godoy[20]和吳吉祥等[21]的研究中指出生育期、始花期、鈴期和果枝始節都是以加性效應為主,而顯性效應則在次之;在研究中,組合Ⅰ各性狀第一對主基因的加性效應分別為4.174、3.671和6.365,顯性效應分別為4.684、-0.121和-0.283,各性狀第一對主基因的加性效應均大于其顯性效應;第二對主基因的加性效應分別為5.189、0.511、3.513,顯性效應分別為2.709、-4.480、-4.091,加性效應大于顯性效應。組合Ⅱ各性狀的第一對主基因和第二對主基因的加性效應分別為3.474、7.473、8.460和1.402、2.613、4.450,顯性效應分別為-1.172、-4.149、-1.000和-1.393、-4.658、1.298,加性效應大于顯性效應,可見兩組合各時期主基因的加性效應均大于顯性效應,結果與其相符,且董娜[7]的研究結果也是如此。而周有耀等[22]則認為早熟性狀的遺傳應為顯性效應;范術麗[8]在對播種-開花,全生育期和開花-吐絮研究結果的數據中表示,播種-開花和全生育期的加性效應分別為-6.257和-9.697 2,顯性效應分別為-3.451 0和-6.527 58,加性效應小于顯性效應,開花-吐絮的第一對主基因的加性效應為-12.443,顯性效應為-11.751,加性效應小于顯性效應;第二對主基因的加性效應和顯性效應分別為12.488和-6.474,加性效應大于顯性效應,兩人結果基本一致。董承光[15],宋美珍等[23]均認為這些結果與研究的遺傳材料、環境條件及當時年份有關。研究結果中花鈴期的遺傳模型和范術麗[8]的研究結果相符均為E-1模型,且主基因遺傳率均大于多基因遺傳率,說明花鈴期是以主基因控制的可以在多環境下穩定遺傳的性狀,可以作為短季棉育種選擇的重要標志。
4 結 論
研究以生育期差異顯著的四個陸地棉品種為親本,聯合其構建的F1、F2為材料,利用多世代混合遺傳分析的方法,明確了兩個雜交組合播種-開花性狀的遺傳同時符合E模型,花鈴期和全生育期性狀均符合E-1模型,播種-開花,花鈴期,全生育期三個性狀可以穩定遺傳,且都受兩對主基因和多基因控制。組合Ⅰ播種-開花、花鈴期、全生育期的主基因的遺傳率分別為63.007%、67.675%、60.875%,多基因的遺傳率分別為36.993%、32.325%、39.125%;組合Ⅱ播種-開花、花鈴期、全生育期的主基因遺傳率分別為64.298%、86.405%、70.837%,多基因遺傳率分別為35.702%、13.595%、29.163%,數據顯示,兩個雜交組合控制生育期性狀的主基因的遺傳力要高于多基因的遺傳率,兩個雜交組合的三個生育期性狀主要由主基因控制,同時還受到微效多基因的調控。聯合兩個雜交組合的表型數據來看,組合Ⅰ分離后代的三個表型數據都偏向早熟親本,組合Ⅱ分離后代中除F1的花鈴期和F2的播種-開花偏向晚熟,其他分離后代的三個表型數據均偏向早熟,兩個雜交組合的表型總體偏向早熟;且兩個雜交組合播種-開花、花鈴期、全生育期的主基因的加性效應均要高于顯性效應。遺傳力高,加性效應大的生育期性狀可以做為選育早熟棉品種的重要指標[20],并且,存在主基因遺傳的生育期性狀可以作為數量性狀分子標記定位的主攻方向。
References)
[1] 喻樹迅,張雷,馮文娟.快樂植棉-中國棉花生產的發展方向[J].棉花學報,2015,(3):283-290.
YU Shu-xun, ZHANG Lei, FENG Wen-juan. (2015). Easy and Enjoyable Cotton Cultivation: Developments in China's Cotton Production [J].CottonScience, (3):283-290.(in Chinese)
[2] 喻樹迅,張雷,馮文娟.棉花生產規模化、機械化、信息化、智能化和社會服務化發展戰略研究[J].中國工程科學,2016,(1):137-148.
YU Shu-xun, ZHANG Lei, FENG Wen-juan. (2016). Study on Strategy of Large Scale, Mechanization, Informationization, Intelligence and Social Services for Cotton Production [J].EngineeringSciences, (1):137-148. (in Chinese)
[3] 喻樹迅.我國棉花生產現狀與發展趨勢[J].中國工程科學,2013,(4):9-13.
YU Shu-xun. (2013). Present situation and development trend of cotton production in China [J].EngineeringSciences, 15(4): 9-13. (in Chinese)
[4] 楊偉華,王延琴,周大云,等.植物修復重金屬污染土壤研究進展[J].湖南農業科學,2014,(18):41-44.
YANG Wei-hua, WANG Yan-qin, ZHOU Da-yun, et al. (2014). Research Progress in Remediation of Heavy-Metal Contaminated Soils by Planting Cotton [J].HunanAgriculturalSciences, (18):41-44. (in Chinese)
[5] 喻樹迅,范術麗.我國棉花遺傳育種進展與展望[J].棉花學報,2003,15(2):120-124.
YU Shu-xun, FAN Shu-li. (2003). The Evolutions and Prospect of Cotton Genetics and Breeding in China [J].CottonScience, 15(2):120-124. (in Chinese)
[6] 袁有祿,張天真,郭旺珍,等.棉花高品質纖維性狀的主基因與多基因遺傳分析[J].遺傳學報,2002,29(9):827-834.
YUAN You-lu, ZHANG Tian-zhen, GUO Wang-zhen, et al. (2002). Major-polygene effect analysis of super quality fiber properties in upland cotton (GossypiumhirsutumL.)[J].ActaGeneticaSinica, 29(9): 827-834. (in Chinese)
[7] 董娜.短季棉早熟及相關性狀的混合遺傳與QTL定位研究[D].新鄉:河南科技學院碩士論文,2010.
DONG Na. (2010).MixedInheritanceandQTLLocationStudyonEarlinessanditsReiatedTraitsofShortSeasonCotton[D]. Master Dissertation. Henan Institute of Science and Technology, Xinxiang. (in Chinese)
[8] 范術麗.短季棉早熟性相關性狀的遺傳及QTL 定位研究[D].北京:中國農業科學院博士論文, 2004.
FAN Shu-li. (2004).StudyoninheritanceofearlinessanditsrelativetraitsofshortseasoncottonandQTLsmapping[D].PhD Dissertation. The Chinese Academy of Agricultural Sciences, Beijing. (in Chinese)
[9] 喻樹迅,黃禎茂.短季棉品種早熟性構成因素的遺傳分析[J].中國農業科學,1990,23(6):48-54.
YU Shu-xun, HUANG Zhen-mao.(1990). Inheritance analysis on earliness components of short season cotton varieties in G.hirsutum[J].ScientiaAgriculturaSinica, 23(6): 48-54. (in Chinese)
[10] 艾先濤,李雪源,王俊鐸,等.北疆陸地棉育成品種表型性狀遺傳多樣性分析[J].分子植物育種,2011,9(1):113-122.
Ai Xian-tao, Li Xue-yuan, Wang Jun-duo, et al. Genetic Diversity on Agronomic Phenotypes in Upland Cotton Varieties of North Xinjiang Area[J].MolecularPlantBreeding, 2011,9(1):113-122. (in Chinese)
[11] 章元明,蓋均鎰,張孟臣.利用P1、F1、P2和F2或F2∶3世代聯合的數量性狀分離分析[J].西南農業大學學報,2000,22(1):6-9.
ZHANG Yuan-ming, GAI Jun-yi, ZHANG Meng-chen. (2000). Jointly segregating analysis of P1P2F1and F2or F2∶3families [J].JournalofSouthwestAgriculturalUniversity, 22(1): 6-9. (in Chinese)
[12] 林忠旭,馮常輝,郭小平,等. 陸地棉產量、纖維品質相關性狀主效 QTL 和上位性互作分析[J]. 中國農業科學,2009,42(9):3 036-3 047.
Lin Zhong-xu, Feng Chang-hui, Guo Xiao-ping, et al. (2009). Genetic Analysis of Major QTLs and Epistasis Interaction for Yield and Fiber Quality in Upland Cotton[J].ScientiaAgriculturaSinica, 42(9):3,036-3,047. (in Chinese)
[13] 柳賓.棉花早熟性、產量和纖維品質性狀的遺傳分析和QTL定位[D]. 山東:山東農業大學,2010.
Liu Bin. (2010).GeneticAnalysisandQTLMappingforMaturity,YieldandFiberQualityTraitsofCotton[D]. Shandong: Shandong Agricultural University.(in Chinese)
[14] 殷劍美,武耀廷,朱協飛.陸地棉產量與品質性狀的主基因與多基因遺傳分析[J].棉花學報,2003,15(2): 67-72.
YIN Jian-mei, WU Yao-ting, ZHU Xie-fei, et al. (2003). Genetic analysis of yield traits and fiber qualities by using major gene plus polygene mixed inheritance model in upland cotton (G.hirsutumL.)[J].CottonScience, 15(2):67-72. (in Chinese)
[15] 董承光,王娟,周小鳳,等.新疆早熟陸地棉早熟性狀的遺傳分析[J].西北農業學報,2014,23(12):96-101.
DONG Cheng-guang, WANG Juan, ZHOU Xiao-feng, et al. (2014). Inheritance of earliness traits in Xinjiang early maturityupland cotton (GossypiumhirsutumL.)[J].ActaAgriculturaeBoreali-OccidentalisSinica, 23(12):96-101. (in Chinese)
[16] 杜雄明,汪若海,劉國強,等.棉花纖維相關性狀的主基因-多基因混合遺傳分析[J].棉花學報,1999,11(2):73-78.
DU Xiong-ming, WANG Ruo-hai, LIU Guo-qiang, et al. (1999). Inheritance Analysis of the Characters Related with Fiber by Using Mixed Major Gene and Polygene Model [J].CottonScience, 11(2):73-78. (in Chinese)
[17] 范術麗,喻樹迅,原日紅,等.短季棉早熟性的遺傳效應及其與環境互作研究[J].西北植物學報,2006,26(11):2 270-2 275.
FAN Shu-li, YU Shu-xun, YUAN Ri-hong, et al. (2006). Genetic effects and environmental interactions of early maturity in short-season cotton [J].ActaBotanicaBoreali-OccidentaliaSinica, 26(11): 2,270-2,275. (in Chinese)
[18] 詹有俊,楊濤,孫建船,等.特早熟陸地棉的遺傳效應及雜種優勢分析[J].農業現代化研究,2012,33(4):943-947.
Zhan You-jun, Yang Tao, Sun Jian-chuan, et al. Analysis of Genetic Effect and Heterosis of Special-early MatureGossypiumhirsutumL. [J].Researchofagriculturalmodernization, 2012,33(4):943-947. (in Chinese)
[19] 詹有俊,楊濤,孫建船,等.特早熟陸地棉熟性產量品質的遺傳相關分析[J].農業現代化研究,2013,34(1):118-121.Zhan You-jun, Yang Tao, Sun Jian-chuan, et al. (2013). Genetic Correlation Analysis of Early Maturity Yield and Quality in Special-early MatureGossypiumhirsutumL. [J].Researchofagriculturalmodernization, 34(1):118-121. (in Chinese)
[20] Godoy, A. S., & Palomo, G. A. (1999). Genetic analysis of earliness in upland cotton (Gossypiumhirsutuml.). ii. yield and lint percentage.Euphytica, 105(2):161-166.
[21] 吳吉祥,朱軍,季道藩,等.陸地棉產量性狀的遺傳效應及其與環境互作的分析[J].遺傳,1995,17(5):1-4.
WU Ji-xiang, ZHU Jun, JI Dao-fan, et al. (1995). Analysis of Genetic Effect × Environment Interactions for Yield Traits in Upland Cotton [J].Hereditas, 17(5):1-4. (in Chinese)
[22] 周有耀.棉花早熟性與纖維品質性狀關系的研究[J].中國棉花,1990,17(5):13-15.
ZHOU You-yao. (1990). Study on the relationship between cotton maturity and fiber quality traits [J].ChinaCotton, 17(5):13-15. (in Chinese)
[23] 宋美珍,喻樹迅,范術麗,等.短季棉主要農藝性狀的遺傳分析[J].棉花學報,2005,17(2):94-98.
SONG Mei-zhen, YU Shu-xun, FAN Shu-li, et al. (2005). Genetic analysis of main agronomic traits in short-season upland cotton (Gossypium hirsutum L.) [J].CottonScience, 17(2): 94-98. (in Chinese)
[24] 艾尼江,朱新霞,管榮展,等.棉花生育期的主位點組遺傳分析[J].中國農業科學,2010,43(20):4 140-4 148.
AI Ni-jiang, ZHU Xin-xia, GUAN Rong-zhan, et al. (2010). Genetic Analysis of Major Locus Group Constitutions of Growth Stages in Upland Cotton [J].ScientiaAgriculturaSinica, 43(20):4,140-4,148. (in Chinese)
Supported by:The National Key Technology R&D Program "The project of drought resistant, saline-alkaline tolerant cotton cultivar screening and breeding " (2014BAD03B01)
Genetic Analysis of Growth Traits in Short Season Cotton
TIAN feng1, 2,FAN Shu-li2,WEI Heng-ling2,WANG Han-tao2,ZHAO Shu-qi2,3,PANG Chao-you2,HU Shou-lin1,YU Shu-xun2
(1.CollegeofPlantScience,TarimUniversity,AlarXinjiang843300,China; 2.InstituteofCottonResearchofCAAS/StateKeyLaboratoryofCottonBiology,AnyangHenan455000,China;3.HuanggangAcademyofAgriculturalSciences,HuanggangHubei438000,China)
【Objective】 To study the genetic law of short-season cotton growth period.【Method】Our study utilized four upland cotton cultivars to construct two crosses and analyzed three traits (Flowering time FT, Flowering and boll-setting period FBP, Whole growing period WGP) of growth period using the joint segregation analysis method of mixed major gene plus polygene inheritance model.【Result】The results from phenotypic analysis showed that: the other phenotypic traits were in favor of the early-maturing parent and the partial early-maturing parent, except for the combination of F1in the blooming stage and F2sowing-flowering stage. The genetic analysis results showed that: the same traits of the combination I and II were in accordance with the same genetic model, and were controlled by two pairs of major genes.【Conclusion】It can be seen from the heritability of major genes and polygenes, the three phenotypic traits were mainly controlled by the major genes and the minor genes, the heritability of the three phenotypic traits is high, and the additive effect is large. This study could provide a theoretical basis for QTL mapping and molecular marker assisted breeding of cotton early maturing traits.
short season cultivars; growth stages; major gene plus polygene inheritance; genetic analysis
2016-11-07
國家科技支撐計劃“抗旱、耐鹽堿棉花品種篩選及新品種培育”(2014BAD03B01)
田鋒(1990-),男,新疆阿克蘇人,碩士研究生,研究方向為棉花遺傳育種,(E-mail)tian_feng12@sina.com
龐朝友(1979-),男,河北邢臺人,副研究員,研究方向為棉花遺傳育種,(E-mail)chypang@163.com 胡守林(1968-),男,新疆阿克蘇人,教授,研究方向為棉花育種,(E-mail)179390139@qq.com
S562.1;S503.2
A
1001-4330(2017)02-0197-09
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
核科學與工程(2021年4期)2022-01-12 06:30:26
今日農業(2020年19期)2020-12-14 14:16:52
小學生必讀(中年級版)(2020年9期)2020-12-04 02:07:22
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
中學物理·高中(2016年12期)2017-04-22 11:53:03
光學精密工程(2016年6期)2016-11-07 09:07:19
核科學與工程(2015年4期)2015-09-26 11:59:03
小櫻桃·童年閱讀(2014年11期)2014-12-01 22:21:30
