高寒地區燕麥 (Avena sativa L.) 人工草地生物量分配對施肥和混播措施的響應
2017-04-14 05:28:24劉文輝張英俊師尙禮孫建魏小星
植物營養與肥料學報 2017年2期
劉文輝,張英俊,師尙禮,孫建,魏小星
(1 甘肅農業大學草業學院,甘肅蘭州 730070;2 青藏高原優良牧草種質資源研究省級重點實驗室 / 青海省畜牧獸醫科學院,青海西寧 810016;3 中國農業大學草地研究所,北京 100094;4 中國科學院地理科學與資源研究所,北京 100101)
高寒地區燕麥 (Avena sativa L.) 人工草地生物量分配對施肥和混播措施的響應
劉文輝1,2,張英俊1,3*,師尙禮1,孫建4,魏小星2
(1 甘肅農業大學草業學院,甘肅蘭州 730070;2 青藏高原優良牧草種質資源研究省級重點實驗室 / 青海省畜牧獸醫科學院,青海西寧 810016;3 中國農業大學草地研究所,北京 100094;4 中國科學院地理科學與資源研究所,北京 100101)
【目的】了解高寒地區燕麥人工草地生物量分配格局對燕麥品種、施肥措施和混播水平的響應機制,為高寒地區人工草地建植提供依據。 【方法】采用“三因素四水平”正交試驗設計,包括 4 個燕麥品種,分別為青燕 1 號、林納、青海 444、青海甜燕麥;4 種施肥措施,分別為不施任何肥料 (CK0)、尿素 75 kg/hm2+ 磷酸二銨 150 kg/hm2(NP)、有機肥 1500 kg/hm2(OM) 和尿素 37.5 kg/hm2+ 磷酸二銨 75 kg/hm2+ 有機肥 750 kg/hm2(NP + OM);4 個箭筈豌豆混播水平,分別為 0、45、60、75 kg/hm2,在燕麥拔節期、抽穗期、開花期和乳熟期分別對各處理燕麥和箭筈豌豆各器官生物量進行了測定。 【結果】燕麥人工草地地上生物量以青燕 1 號 (85.57%)、尿素 + 磷酸二銨 + 有機肥處理 (84.91%) 和 0 kg/hm2混播水平 (85.81%) 時分配比例最高;地下生物量以林納(18.04%)、不施肥 (16.02%) 和 30 kg/hm2混播水平 (16.29%) 時分配比例最高。莖、葉、穗和根生物量分配比例分別為 44%~48%、19%~24%、17%~22% 和 14%~18%。莖、葉、穗和地下生物量分配隨季節分別呈增加、先降后增、增加和下降的變化趨勢;在各因素影響下,燕麥生長前期以葉生長為主,中、后期以莖生長為主,而箭筈豌豆在生長前期以根生長為主,中、后期以葉生長為主。燕麥生物量分配在整個生育期處于優勢地位,地上生物量分配始終高于地下,表現為異速生長關系。莖和地下生物量分配在生長前期和后期分別受施肥措施和品種的影響;葉生物量分配在整個生育期受混播水平的影響;燕麥穗生物量分配主要受品種和混播水平的影響。 【結論】品種、施肥措施和箭筈豌豆混播水平均對高寒地區燕麥人工草地生物量分配具有顯著影響,選用青燕 1 號品種,單播,配施尿素 37.5 kg/hm2、磷酸二銨 75 kg/hm2、有機肥 750 kg/hm2時地上生物量分配最高;選用林納,并混播 30 kg/hm2的箭筈豌豆,采用不施肥措施時地下生物量分配最高。
高寒地區;燕麥品種;施肥;混播;生物量分配
植物響應環境條件的變化是通過調節各器官的生物量分配,以最大化地獲取光、營養和水等受限資源。生物量分配格局的可塑性貫穿植物的整個生活史,決定著植物在不同環境中獲取資源的能力[1]。植物在整個生長發育過程中,不同功能器官對資源的利用始終存在著競爭,植物只有通過優化資源分配以適應環境變化,才能維持其生存。
建立優質、高產的人工草地是解決青藏高原高寒地區草畜季節性供求矛盾、保護生態環境和促進草地畜牧業可持續發展的關鍵措施之一。青藏高原地區一年生人工草地建設主要以燕麥 (Avena sativa L.)單播、燕麥與箭筈豌豆 (Vicia sativa L.) 混播為主。禾本科和豆科牧草的混播草地以較高的生產力、較好的營養搭配和經濟有效的氮素來源而長期受到人們的重視。混播草地的生長動態,尤其是生物量的積累和分配動態變化是草地群落內競爭力大小的具體參考指標。在燕麥人工草地的研究方面,國內外科研人員從燕麥單播[2]、燕麥與箭筈豌豆混播的品種選擇[3]、混播組合[4]、施肥水平[5]、光能轉化效率[6]、生產性能[7]以及營養組成[5]等方面進行的深入而細致的理論研究和生產實踐[6],有效促進了燕麥人工草地初級生產力的提高。以往的大量研究多集中在單一因素 (品種、肥料、密度) 確定最優化分配格局,忽略了植物生長過程中各器官間生物量的分配格局對草地生態和生產方面的影響。本研究以青藏高原燕麥人工草地建植過程中選用的 4 個燕麥品種為研究對象,采用不同的施肥措施和箭筈豌豆混播比例,建立燕麥與箭筈豌豆混播人工草地,從草地生態和生產方面研究植物各器官資源分配的動態變化,以了解植物生物分配格局對品種、施肥措施和混播水平的響應機制。
1 材料與方法
1.1 試驗地概況
試驗于 2014 年在青海省海北州西海鎮 (36°59.36′E、100°52.848′N) 進行。海拔 3156 m,氣候寒冷潮濕,無絕對無霜期,年平均氣溫 0.5℃,年降水量 369.1 mm,且多集中在 7、8、9 三個月,年蒸發量為 1400 mm,全年日照時數在 2980 h,平均無霜期為 93 d 左右。土壤為栗鈣土,pH 值 8.43,全氮 (N) 1.56 g/kg,全磷 (P2O5) 1.39 g/kg,全鉀 (K2O) 22.06 g/kg,堿解氮88.77 mg/kg,速效磷 2.2 mg/kg,速效鉀 168.2 mg/kg,有機質 32.48 g/kg。
1.2 試驗材料與方法
本試驗為品種、施肥和箭筈豌豆混播水平三因素四水平正交試驗設計[L16(43),表 1],共 16 個處理,完全隨機區組排列,3 次重復。燕麥品種 4 個,分別為青燕 1 號 (A1)、林納 (A2)、青海 444 (A3) 和青海甜燕麥 (A4);施肥措施 4 個,分別為不施任何肥料 (B1)、尿素 75 kg/hm2+ 磷酸二銨 150 kg/hm2(B2)、有機肥 1500 kg/hm2(B3) 和尿素 37.5 kg/hm2+ 磷酸二銨 75 kg/hm2+ 有機肥 750 kg/hm2(B4);箭筈豌豆混播水平 4 個,分別為 0 kg/hm2(C1)、45 kg/hm2(C2)、60 kg/hm2(C3) 和 75 kg/hm2(C4)。試驗小區面積為20m2(4 m × 5 m),小區間隔 0.5 m。試驗選用肥料為尿素,磷酸二銨 (N 16%、P2O546%),有機肥 (有機質 > 40%、N + P2O5+ K2O 25%)。
燕麥播量按 600 × 104plant/hm2保苗數計算,根據種子用價計算得青燕 1 號、林納、青海 444 和青海 甜 燕 麥 的 實 際 播 量 分 別 為154.5 kg/hm2、150.0 kg/hm2、183.0 kg/hm2和 216.0 kg/hm2,撒 播 , 燕 麥 與 箭 筈 豌 豆 5 月 14 日 同 時 播 種 , 播深 3~4 cm。肥料在播種前一次性施入。出苗后,人工除雜草 1 次。分別于燕麥拔節期 (6 月 15 日)、抽穗期 (7 月 15 日)、開花期 (8 月 15 日) 和乳熟期 (9 月15 日) 在各試驗小區選取 1 m × 1 m 的樣方(3 次重復),連同地下部分挖出,燕麥和箭筈豌豆按根、莖、葉、穗 (燕麥) 分開,帶回實驗室,洗凈地下生物量,置于 85℃ 烘箱中烘 3 h 后分別稱重。
1.3 數據處理
采用 Excel 2003 對數據進行初步整理,用 SPSS for Windows 11.5 進行方差分析,采用 Duncan 法在0.05 水平上進行多重比較。用 Sigmaplot 10.0 進行繪圖。
相關生長是以冪函數的形式描述生物個體大小和其它屬性間的關系[8],可用 LogY = a LogX + b 表示(其中 Y 為地上生物量,X 為地下生物量,a 為異速生長指數,b 為異速生長常數)。根據 a 值的不同 , 地 上 和 地 下 生 物 量 生 長 關 系 存 在3 種 形式:a = 1.0,表現為等速生長,即地上和地下生物量增加速率相等;a > 1.0,表現為地上生物量積累速率高于地下;a < 1.0,表示地下生物量積累速率高于地上。利用標準主軸法 (standardized major axis,SMA)計算異速生長方程的指數 a,和 95% 置信區間 (CI)及決定系數 (R2)[9]。

表1 正交試驗試驗設計方案 L16(43)Table 1 The orthogonal experiment design L16(43)
2 結果與分析
2.1 生物量構件分配模式
2.1.1品種、施肥措施和混播水平對各器官生物量分配的影響 表 2 顯示,品種因素影響下,地上總生物量 分 配 比 例 以 青 燕 1 號 (85.57%) 和 青 海 甜 燕 麥(85.62%) 最高,地下生物量以林納 (18.04%) 和青海444 (17.29%) 分配比例最高;施肥措施影響下,地上以尿素 + 二銨 + 有機肥 (84.91%) 和尿素 + 二銨(84.42%) 最高,地下以不施肥 (17.14%) 和有機肥(16.02%) 最高;混播因素影響下,地上以 0 kg/hm2最高 (85.81%),地下以 30 kg/hm2最高 (16.29%)。在3 個因素影響下,莖、葉、穗和根生物量分配比例分別為 44%~48%、19%~24%、17%~22% 和14%~18%。
2.1.2生物量分配季節動態 表 3 顯示,在 3 個因素影響下,燕麥人工草地總莖生物量分配隨生育期的推 進 而 顯 著 增 加 , 至 開 花 - 乳 熟 期 最 大(約 為44%~48%),其中燕麥和箭筈豌豆莖生物量分配比例分別在開花期最大 (41.43%~41.63%)和乳熟期最大 (7.23%~10.48%)。總葉、燕麥葉生物量分配比例在拔節期最大,平均分別達 49.56%~50.67% 和46.77%~47.53%。總葉生物量分配比例顯著下降至開花期后,在乳熟期有所增加,而燕麥葉生物量分配比例顯著下降至乳熟期,箭筈豌豆葉生物量分配比例隨生育期的推進顯著增加,至乳熟期最大(8.12%~11.67%)。穗分配比例則從開花期-乳熟期呈顯著增加;總根、燕麥根和箭筈豌豆根生物量分配比例均在拔節期最大,分別為 36.10%~37.02%、4.42%~6.27% 和 41.02%~43.20%,從拔節到乳熟期呈顯著下降趨勢。
2.1.3生物量分配策略 從分配策略來看 (表 3),在 3個因素影響下,拔節期、抽穗期、開花期、乳熟期4個時期燕麥生物量分配比例分別為葉 > 根 > 莖、莖 > 葉 > 根、莖 > 穗 > 葉 > 根、莖 > 穗 > 葉 > 根;拔節期以葉和根生物量為主,抽穗期后,莖生物量所占比例最大。4 個時期箭筈豌豆生物量分配比例分別為根 > 葉 > 莖、葉 > 莖 > 根、葉 > 莖 > 根、葉 >莖 > 根;拔節期箭筈豌豆根生物量分配最大,隨后葉生物量占據主導地位。4 個時期總生物量分配比例分別為葉 > 根 > 莖、莖 > 葉 > 根、莖 > 葉 > 穗 >根、莖 > 葉 > 穗 > 根;總生物量分配比例在拔節期和抽穗期與燕麥生物量分配比例一致,開花期和乳熟期莖生物量占主導地位,其次為葉和穗生物量。

表2 不同措施對開花期生物量構件分配比例的影響 (%)Table 2 Effect of different treatments on the biomass allocation ratio at the flowering stage

表3 不同措施對生物量構件分配比例的季節變化 (%)Table 3 Effect of different treatments on the biomass allocation ratio in the growth period
2.2 三個因素對生物量分配的差異性分析
表 4 顯示,3 個因素對地上和地下總生物量分配的 差 異 性 均 達 到 顯 著(P <0.05) 或 極 顯 著 水 平(P < 0.01)。拔節期對地上和地下總生物量的影響大小為施肥措施 > 品種 > 混播水平;而其余時期均表現為品種 > 混播水平 > 施肥措施。
總莖、燕麥莖、箭筈豌豆莖生物量分配在拔節期施肥措施對其影響最大,其余時期品種對總莖和箭筈豌豆莖影響最大;抽穗期品種對燕麥莖影響最大,開花-乳熟期混播水平對燕麥莖影響最大。葉生物量分配在整個生育期均表現為混播水平對其的影響最大,混播水平是影響葉生物量分配的關鍵因素。穗生物量分配在開花期受品種影響最大,在乳熟期受混播水平影響最大。
地下總生物量分配在拔節期受施肥措施影響最大,其它時期受品種影響最大;燕麥根生物量分配在乳熟期以混播水平對其的影響最大,其它時期以品種影響最大;箭筈豌豆根生物量分配在拔節和乳熟期受品種影響最大,而在抽穗和開花期受施肥措施水平影響最大。
2.3 相關生長關系分析
2.3.1施肥措施下的相關生長關系 圖 1 顯示,不施肥處理在開花期、尿素+磷酸二銨+有機肥處理在抽穗期和乳熟期,燕麥人工草地地下與地上生物量間均表現為等速生長 (P > 0.05),各施肥處理在其余時期表現為異速生長 (P < 0.05)。拔節期不施肥、尿素+磷酸二銨和有機肥三個處理下的斜率小于 1,表明地下生物量積累速率高于地上;抽穗期、開花期和乳熟期各施肥處理的斜率大于 1,表明地上生物量積累速率大于地下,這與上述研究結果一致。
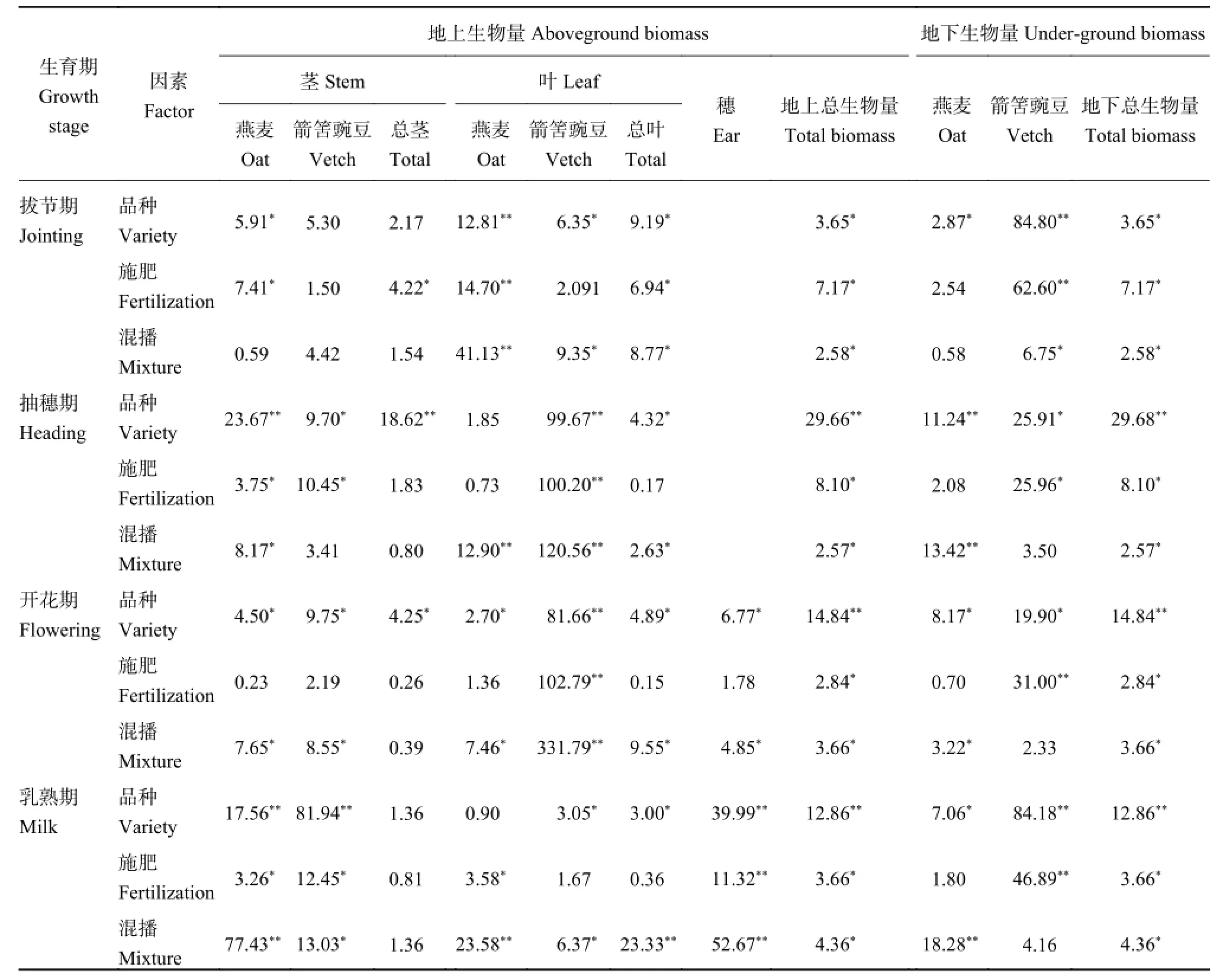
表4 不同措施對地上、地下生物量分配的差異性分析Table 4 Effect of different measures on the aboveground and underground biomass allocation
2.3.2混播處理下的相關生長關系 圖 2 顯示,開花期箭筈豌豆混播 45 kg/hm2和拔節、抽穗、開花期混播 75 kg/hm2時,燕麥人工草地地下與地上生物量間表現為等速生長 (P > 0.05),而其余各時期各混播水平下則表現為異速生長 (P < 0.05)。拔節期混播 45 kg/hm2和 75 kg/hm2的斜率小于 1,表明地下生物量積累速率高于地上,抽穗期、開花期和乳熟期各混播處理的斜率大于 1,表明地上生物量積累速率大于地下,這與上述研究結果一致。
3 討論
3.1 3 種措施對生物量分配的影響
目前許多學者在玉米[10]、燕麥[11]等生物量分配方面的研究均提出,品種對生物量分配具有顯著影響,本研究也得出了一致的結論。4 個燕麥品種中青燕 1 號和青海甜燕麥具有較高的地上生物量分配比例,而林納和青海 444 具有較高的地下生物量分配比例,這與品種的遺傳性狀有關。青燕 1 號通過增加莖生物量分配,青海甜燕麥通過葉和穗生物量分配的增加實現較高的地上生物量分配。

圖1 不同施肥措施下地上、地下生物量生長關系Fig. 1 The relationship between aboveground and underground biomass under different fertilizations[注(Note):U—尿素 Urea;NP—磷酸二銨 NH4H2PO4;OM—有機肥 Organic manure.]
施肥通過增加土壤養分含量來影響植物對肥料的吸收、利用和轉化。趙宏魁等[12]研究指出,施肥對提高燕麥人工草地地上生物量的分配效果高于地下;寇明科等[13]指出,有機和無機肥配施可有效提高燕麥與箭筈豌豆混播人工草地地上生物量。本研究發現,施肥措施顯著影響燕麥人工草地地上、地下生物量分配,地上生物量分配以尿素 + 磷酸二銨 +有機肥處理和尿素 + 磷酸二銨處理最高;地下生物量分配以不施肥和單施有機肥處理最高。施肥通過增加地上部分的生物量,進而減少地下根系分配比例[14];但在低養分下,植物相對更多地分配到根系以增強必要的養分補充[15]。
密度是影響燕麥人工草地生物量分配的重要因素[16]。本研究發現,箭筈豌豆混播水平顯著影響燕麥人工草地各器官生物量的分配,地上生物量分配以單播燕麥最高,地下生物量分配以 30 kg/hm2混播水平最高。燕麥單播時,減少了與箭筈豌豆在光照、溫度、水分等方面的競爭,燕麥地上各器官所占比例較高,隨著混播比例的增加,燕麥與箭筈豌豆競爭加劇,抑制了燕麥的生長,從而減少了地上生物量的分配。
3.2 生物量分配策略

圖2 不同混播措施下地上、地下生物量生長關系Fig. 2 The relationship between aboveground and underground biomass under different mixture levels
燕麥人工草地在品種、施肥和混播措施的影響下,為更好地適應環境,實現自身適合度最大化,最終保持植物的持續生存和繁衍,各器官的資源分配方面都存在“此消彼長”的關系[17]。本研究中,生長前期,燕麥以葉生長為主,中、后期以莖生長為主,箭筈豌豆生長前期以根生長為主,中、后期以葉生長為主。由于燕麥所占比重大于箭筈豌豆,燕麥和箭筈豌豆各器官生物量分配最終導致總生物量分配表現為生長前期以葉為主,中、后期以莖為主。根系在各器官生物量分配中始終處于最低。
根據 Donald 的理論,在發生競爭的種群中,占有更多有限資源的植物會以其它植物的犧牲為代價[18]。本研究發現,不同燕麥品種、肥料配比和箭筈豌豆混播水平下建植的混播草地中,燕麥在混播群落中始 終 處 于 優 勢 地 位 , 這 與 張 雪 洲 等[19]的 研 究 結 果 一致。Weiner 和 Thomas[20]將競爭分為單側競爭和雙側競爭。當光資源受限時,植物分配更多的資源給莖和葉的生長,表現為“不對稱”的單側競爭;而營養受限時,植物往往增大根的產量,表現為“對稱”的雙側競爭[21]。本研究中,燕麥、箭筈豌豆和總生物量地上部分生長高于地下部分的生長,地上部分對資源的競爭始終處于主導地位,表明在整個生育期植物一直處于對光資源的競爭上,燕麥混播群落不受營養的限制,這與宋清華等[22]、張靜等[16]、雷占蘭等[23]的研究結果一致。
3.3 地上和地下生物量相關生長關系
本研究發現,不同施肥措施下,以不施肥處理在開花期、尿素 + 磷酸二銨 + 有機肥處理在抽穗期和乳熟期燕麥人工草地地上、地下生物量表現為等速生長,其余時期均表現為異速生長,且地上生物量積累速率大于地下;不同混播措施下,以開花期 45 kg/hm2混 播 水 平 和 拔 節 期 、 抽 穗 期 、 開 花期 75 kg/hm2混播水平下地上、地下生物量表現為等速生長,其余時期為異速生長,且在拔節期表現為地下生物量高于地上,其余時期地上生物量積累高于地下。
3.4 三個因素對生物量分配的差異
很多研究[24-26]提出,生物量構建分配模式不僅受土壤營養、水熱條件等環境因子的影響,在一定程度上還受水肥管理等措施的影響。本研究發現,品種、施肥措施和混播水平均能顯著影響燕麥人工草地各器官的生物量分配,且 3 個因素對生物量分配影響的大小不同。施肥措施和品種主要影響生長前期和后期的生物量分配。其中莖生物量分配在整個生長前期受施肥措施影響最大,中、后期受品種影響最大;混播水平是影響葉和穗生物量分配的關鍵因素;地下生物量分配在生長前期和后期分別受施肥措施和品種影響最大。本研究采用正交試驗設計的方法討論了品種、施肥措施和混播水平對高寒地區燕麥人工草地各器官生物量分配的影響,受試驗設計的限制,未考慮 3 個因素間的交互效應。
4 結論
品種、施肥措施和箭筈豌豆混播水平均能顯著改變燕麥人工草地生物量分配格局。莖生物量分配在生長前期主要受施肥措施的影響,后期受品種的影響;葉生物量分配在整個生育期受混播水平的影響;穗生物量分配主要受品種和混播措施的影響;地下生物量分配在前期和后期分別受施肥措施和品種影響最大。
燕麥人工草地生長前期以葉為主,中后期以莖為主。在品種 (青燕 1 號和青海 444)、施肥措施 (尿素 + 二銨 + 有機肥處理和尿素 + 二銨)和不混播條件下燕麥人工草地地上生物量分配最高。
燕麥、箭筈豌豆和總生物量地上部分生長高于地下部分的生長,地上部分對資源的競爭始終處于主導地位。整個生育期多表現為異速生長關系。
[1]Poorter H, Remkes C, Lambers H. Carbon and nitrogen economy of 24 wild species differing in relative growth rate[J]. Plant Physiology, 1990, 94(2): 621-627.
[2]田福平, 時永杰, 周玉雷, 等. 燕麥與箭筈豌豆不同混 播比例對生物量的影響研究[J]. 中國農學通報, 2012, 28(20): 29-32. Tian F P, Shi Y J, Zhou Y L, et al. Effect of different mixture proportion of oat and common vetch on biomass[J]. Chinese Agricultural Science Bulletin, 2012, 28(20): 29-32.
[3]王桃. 高寒 牧 區36種 燕 麥營養生態特性及其 生 產 效能評價[D]. 蘭州: 蘭州大學碩士學位論文, 2010. Wang T. Evaluation on nutritional and ecological characteristics and productive efficiency[D]. Lanzhou: MS Thesis of Lanzhou University, 2010.
[4]馬春暉 , 韓建國. 高寒地區燕麥及其混播草地最佳刈割期 的 研 究[J]. 塔里木農墾大學學報, 2000, 12(3): 15-19. Ma C H, Han J G. The studies on the optimal harvest time of oat and vetch in both monoculture and mixture in high-cold area[J]. Journal of Tarim University of Agricultural Reclamation, 2000, 12(3): 15-19.
[5]楊麗娜, 趙桂琴, 侯 建杰. 播期、 肥 料種類及其配 比對燕麥生長及產量的影響[J]. 中國草地學報, 2013, 35(4): 47-51, 60. Yang L N, Zhao G Q, Hou J J. Effects of seeding dates, fertilizers and their ratio on oat growth and yield[J]. Chinese Journal of Grassland, 2013, 35(4): 47-51, 60.
[6]陳功, 張自和, 胡自 治. 高寒地區 一 年生人工草地 地上生物量動態及光能轉化效率[J]. 草業學報, 2003, 12(1): 69-73. Chen G, Zhang Z H, Hu Z Z. Herbage yield dynamics and solar energy conversion of annual cultivated grassland in Xining City[J]. Acta Prataculturae Sinica, 2003, 12(1): 69-73.
[7]崔瑩. 燕麥和 箭 筈 豌豆混播對人工草 地 生 產性能及土壤性質的 影響[D]. 蘭州: 甘肅農業大學碩士學位論文, 2014. Cui Y. Effect on production performance and soil properties of artificial grassland with mixed sowing oats and common vetch [D]. Lanzhou: MS Thesis of Gansu Agricultural University, 2014.
[8]韓文 軒, 方精云. 冪指數異速生長機制模型綜述[J]. 植物生態學報, 2008, 32(4): 951-960. Han W X, Fang J Y. Review on the mechanism models of allometric scaling laws: 3/4 vs. 2/3 power[J]. Journal of Plant Ecology, 2008, 32(4): 951-960.
[9]楊昊 天, 李新榮, 劉立超, 等. 荒 漠草地4種灌木生物量分配特征[J].中國沙漠, 2013, 33(5): 1340-1348. Yang H T, Li X R, Liu L C, et al. Biomass allocation patterns of four shrubs in desert grassland[J]. Journal of Desert Research, 2013, 33(5): 1340-1348.
[10]宋創業, 郭柯, 劉高煥. 播期對渾善達克沙地青貯玉米產量及生物量分配的影響[J]. 中國生態農業學報, 2008, 16(4): 865-868. Song C Y, Guo K, Liu G H. Effect of sowing date on yield and phytomass allocation of silage maize in Hunshandak sandy land[J]. Chinese Journal of Eco-Agriculture, 2008, 16(4): 865-868.
[11]劉佳. 水分狀況對裸燕麥生物量分配和產量形成的影響[D]. 蘭州:蘭州大學碩士學位論文, 2011. Liu J. Effects of water condition on the biomass allocation and yield formation of naked oats[D]. Lanzhou: MS Thesis of LanzhouUniversity, 2011.
[12]趙宏魁, 馬真, 張春輝, 等. 種植密度和施氮水平對燕麥生物量分配的影響[J]. 草業科學, 2016, 33(2): 249-258. Zhao H K, Ma Z, Zhang C H, et al. The reproductive allocation of Avena sativa under different planting densities and nitrogen addition treatments[J]. Pratacultural Science, 2016, 33(2): 249-258.
[13]寇明科, 王安碌, 張生璨, 等. 不同施肥處理對提高高寒人工混播草地產草量的試驗研究[J]. 草業科學, 2003, 20(4): 14-15. Kou M K, Wang A L, Zhang S C, et al. Study on the effects of different fertilizer treatments on mixed grassland yield in Luqu, Gannan Autonomous Region[J]. Pratacultural Science, 2003, 20(4):14-15.
[14]Coyle D, Coleman M D, Aubrey D P. Above and below-ground biomass accumulation, production, and distribution of sweetgum and loblolly pine grown with irrigation and fertilization [J]. Canadian Journal of Forest Research, 2008, 38(6): 1335-1348.
[15]Haynes B E, Gower S. Belowground carbon allocation in unfertilized and fertilized red pine plantations in northern Wisconsin[J]. Tree Physiology, 1995, 15(5): 1134-1140.
[16]張靜, 趙成章. 高寒山區混播草地燕麥和毛苕子生物量分配格局對組分密度比的響應[J]. 生態學雜志, 2013, 32(2): 266-270. Zhang J, Zhao C Z. Distribution patterns of the biomass of Avena sativa and Vicia villosa in response to the component density ratio of the grass species in a mixed-seeding grassland in alpine region of Qilian Mountains, Northwest China[J]. Chinese Journal of Ecology, 2013, 23(2): 266-270.
[17]張大勇. 植物生活史進化與繁殖生態學[M]. 北京: 科學出版社, 2004. 430. Zhang D Y. Plant life history evolution and reproductive ecology [M]. Beijing: Science Press, 2004. 430.
[18]劉琳, 徐炳成, 李鳳民, 等. 土壤水分對兩個冬小麥品種產量和競爭能力的影響[J]. 生態學報, 2007, 27(8): 3442-3449. Liu L, Xu B C, Li F M, et al. Effects of soil water contents on competitive abilities and yield of two winter wheat cultivars[J]. Acta Ecologica Sinica, 2007, 27(8): 3442-3449.
[19]張學洲, 李學森, 張麗萍, 等. 復種初島燕麥+中豌六號混播效應的動態研究[J]. 草業科學, 2010, 27(11): 101-108. Zhang X Z, Li X S, Zhang L P, et al. Yield and quality of mixedsowing pasture of Avena sativa and Pisum sativum[J]. Pratacultural Science, 2010, 27(11): 101-108.
[20]Weiner J, Thomas S C. Size variability and competition in plant monocultures[J]. Oikos, 1986, 47(2): 211-222.
[21]黎磊, 周道瑋, 盛連喜. 密度制約決定的植物生物量分配格局[J].生態學雜志, 2011, (8): 1579-1589. Li L, Zhou D W, Sheng L X. Density dependence-determined plant biomass allocation pattern[J]. Chinese Journal of Ecology, 2011, (8):1579-1589.
[22]宋清華, 趙成章, 史元春, 等. 祁連山北坡混播草地密度制約下燕麥和毛苕子比根長分布格局[J]. 生態學雜志, 2015, 34(2): 497-503. Song Q H, Zhao C Z, Shi Y C, et al. Spatial distribution patterns of specific root lengths of Avena sativa L. and Vicia villosa Roth in mixed-sowing grassland under density dependence in northern slope of Qilian Mountains[J]. Chinese Journal of Ecology, 2015, 34(2):497-503.
[23]雷占蘭, 周華坤, 劉澤華, 等. 密度氮肥交互處理下高寒地區燕麥的生長特性與生殖分配[J]. 草業科學, 2014, 31(6): 1110-1119. Lei Z L, Zhou H K, Liu Z H, et al. The growth performance and reproductive allocation of Avena sativa under density and nitrogen interaction[J]. Pratacultural Science, 2014, 31(6): 1110-1119.
[24]張莉, 王長庭, 劉偉, 等. 不同建植期人工草地優勢種植物根系活力、群落特征及其土壤環境的關系[J]. 草業學報, 2012, 21(5):185-194. Zhang L, Wang C T, Liu W, et al. Relationships of dominant species root activity, plant community characteristics and soil microenvironment in artificial grassland over different cultivation periods[J]. Acta Prataculturae Sinica, 2012, 21(5): 185-194.
[25]毛偉, 李玉霖, 崔奪, 等. 沙質草地不同生活史植物的生物量分配對氮素和水分添加的響應[J]. 植物生態學報, 2014, 38(2): 125-133. Mao W, Li Y L, Cui D, et al. Biomass allocation response of species with different life history strategies to nitrogen and water addition in sandy grassland in Inner Mongolia[J]. Chinese Journal of Plant Ecology, 2014, 38(2): 125-133.
[26]雷占蘭, 周華坤, 劉澤華, 等. 密度氮肥交互處理下垂穗披堿草生長與生殖特性[J]. 中國草地學報, 2014, 36(2): 12-19. Lei Z L, Zhou H K, Liu Z H, et al. The growth and reproductive characteristics of Elymus nutans under the interaction between different planting density and nitrogen application[J]. Chinese Journal of Grassland, 2014, 36(2): 12-19.
Effect of variety, fertilization and mixture sowing on oat biomass allocation in the alpine cultivated pasture
【Objectives】This study focused on the response of biomass allocation of oat artificial grassland in alpine region to oat variety, fertilization and common vetch mixture level for providing a theoretical basis to establish the artificial grassland. 【Methods】According to the orthogonal design of three factors (variety, fertilization and legume mixture) and four levels, there were four oat varieties, namely Avena sativa L. cv. Qingyan No.1, A. sativa L. cv. Lena, A. sativa L. cv. Qinghai 444 and A. sativa L. cv. Qinghai, four fertilizationmeasures, including without applying any fertilizers (CK0), 75 kg/hm2urea + 150 kg/hm2NH4H2PO4(NP), 1500 kg/hm2organic manure (OM) and 37.5 kg/hm2urea + 75 kg/hm2NH4H2PO4+ 750 kg/hm2organic manure (NP + OM), and four common vetch mixture levels with 0 kg/hm2, 45 kg/hm2, 60 kg/hm2and 75 kg/hm2. Different organ biomass of oats and common vetch were measured at the jointing, heading, flowering and milk stages of oats respectively. 【Results】The aboveground biomass allocation of the oat artificial grassland was the highest under the fertilization of 37.5 kg/hm2urea + 75 kg/hm2NH4H2PO3+ 750 kg/hm2organic manure (NP + OM) (84.91%) with the A. sativa cv. Qingyan No.1 (85.57%) without common vetch mixing (85.81%), while the underground biomass allocation was the highest under the common vetch mixture level 30 kg/hm2(16.29%) with the A. sativa cv. Lena (18.04%) without fertilization (16.02%). The ranges of the biomass allocation rates of stems, leaves, ears and roots were 44%-48%, 19%-24%, 17%-22% and 14%-18% respectively. The biomass allocation rates were changed with the growth stages: increasing, decreasing at the beginning and increasing later, increasing, and decreasing for stem, leaf, ear and root respectively. Under the influence of various factors, it was the oat’s leaves that played a dominant role in the early growing period, and the stems took place in the mid-late period. The common vetch’s root biomass, on the contrary, was leading at the beginning and its leaves took place in the mid-late period. Obviously, the biomass allocation of the oats played a significant role in the whole growing process. Meanwhile, there was a relationship defined as the allometric growth relationship, in other words, the aboveground biomass was always higher than the underground biomass in the whole growth period. The stem and root biomass allocation at the early and late growth stages were affected by the fertilization and variety separately, while the level of mixed sowing in the whole growth period affected the leaf growth, and the fertilization and the mixture level mainly influenced the ear biomass allocation. 【Conclusions】 It was variety, fertilization and the common vetch mixture level that significantly affected the biomass allocation in the alpine region. The highest aboveground biomass allocation was under the fertilization of 37.5 kg/hm2urea + 75 kg/hm2NH4H2PO3+ 750 kg/hm2organic manure with the A. sativa cv. Qingyan No.1 without common vetch, and the highest belowground biomass allocation was the treatment without fertilization with the A. sativa L. cv. Lena mixed with common vetch of 30 kg/hm2.
alpine area; oat variety; fertilization; mixture sowing; biomass allocation
LIU Wen-hui1,2, ZHANG Ying-jun1,3*, SHI Shang-li1, SUN Jian4, WEI Xiao-xing2
( 1 Pratacultural College of Gansu Agricultural University, Lanzhou 730070, China; 2 Key Laboratory of Superior Forage Germplasm in the Qinghai-Tibetan Plateau/Qinghai Academy of Animal Science and Veterinary Medicine, Xining 810016, China; 3 Institute of Grassland Sciences, China Agricultural University, Beijing 100094, China; 4 Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101, China )
2016-05-11 接受日期:2016-08-16
“現代農業產業技術體系建設專項資金”(CARS-35-41);農業部“青藏高原牧草種質資源保護利用”項目(070401);青海省飼草產業科技創新平臺建設資助。
劉文輝(1979—),男,青海貴德人,博士研究生,副研究員,主要從事牧草栽培和育種方面的研究。
Tel:0971-8210200;E-mail:qhliuwenhui@163.com。* 通信作者 Tel:010-62734593;E-mail:zhangyj@cau.edu.cn
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
小讀者(2021年2期)2021-03-29 05:03:48
鐵道通信信號(2020年9期)2020-02-06 09:15:22
華人時刊(2019年13期)2019-11-17 14:59:54
數學大王·趣味邏輯(2019年5期)2019-06-13 20:27:43
小學科學(學生版)(2019年5期)2019-05-21 01:00:18
經濟技術協作信息(2018年30期)2018-11-22 06:20:24
文苑(2018年22期)2018-11-19 02:54:14
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
