降鈣素基因相關肽對血清饑餓作用下MC3T3-E1成骨細胞凋亡和自噬的影響
2017-05-04 00:35:44安洋張慧宇郭俊峰李鑫楊陽張綱譚穎徽
華西口腔醫學雜志 2017年2期
安洋 張慧宇 郭俊峰 李鑫 楊陽 張綱 譚穎徽
第三軍醫大學新橋醫院口腔頜面外科,重慶 400037
降鈣素基因相關肽對血清饑餓作用下MC3T3-E1成骨細胞凋亡和自噬的影響
安洋 張慧宇 郭俊峰 李鑫 楊陽 張綱 譚穎徽
第三軍醫大學新橋醫院口腔頜面外科,重慶 400037
目的 研究血清饑餓條件下降鈣素基因相關肽(CGRP)對小鼠MC3T3-E1成骨細胞凋亡、自噬的影響以及二者之間的關系,以進一步明確CGRP對成骨細胞的保護機制。方法體外培養小鼠MC3T3-E1成骨細胞。采用流式細胞術和蛋白質印跡檢測正常血清、無血清(血清饑餓)、3-MA預處理+血清饑餓培養的成骨細胞的凋亡和微管相關蛋白1輕鏈3(LC3)蛋白的表達。采用蛋白質印跡檢測正常血清、血清饑餓、血清饑餓+不同濃度(10-10、10-9、10-8、10-7mol·L-1)CGRP培養的成骨細胞的LC3和P62蛋白表達;采用蛋白質印跡檢測正常血清、血清饑餓、血清饑餓+10-8mol·L-1CGRP培養不同時間(2、6、12、24、48、72 h)的成骨細胞的LC3蛋白表達,流式細胞數檢測細胞凋亡,MDC染色檢測細胞自噬泡。采用流式細胞術檢測正常血清、血清饑餓、血清饑餓+10-8mol·L-1CGRP、3-MA預處理+血清饑餓、3-MA預處理+血清饑餓+10-8mol·L-1CGRP培養24 h的成骨細胞的凋亡。結果血清饑餓培養時成骨細胞的LC3Ⅱ蛋白表達及細胞凋亡較正常血清時增加,3-MA預處理+血清饑餓培養時成骨細胞的凋亡較血清饑餓時增加(P<0.01)。與正常血清相比,血清饑餓、血清饑餓+CGRP培養時LC3Ⅱ蛋白表達增加,P62蛋白表達降低,以血清饑餓+10-8mol·L-1CGRP培養24 h時LC3Ⅱ/Ⅰ比值最高;血清饑餓+10-8mol·L-1CGRP培養能抑制成骨細胞的凋亡,促進自噬泡的合成。3-MA預處理后MC3T3-E1成骨細胞凋亡增加,CGRP部分逆轉3-MA預處理所增加的成骨細胞凋亡。結論CGRP能夠增強血清饑餓下MC3T3-E1成骨細胞的自噬活性,并可能通過促進自噬抑制成骨細胞的凋亡。
降鈣素基因相關肽; 成骨細胞; 自噬; 凋亡
成骨細胞作為參與骨修復的重要細胞之一,主要依賴于微環境中源源不斷的促生存信號才能維持其正常的生理功能;當存在疾病或損傷導致營養因子供應受到限制時,細胞將會發生過度的凋亡,從而嚴重影響到骨的修復。目前已有大量的生長因子被證明在骨修復中具有重要的作用,但骨修復本身是一個高度精密復雜的調控過程,臨床上仍然不斷地研究和尋找更高效的骨活化劑[1]。
降鈣素基因相關肽(calcitonin gene-related peptide,CGRP)是一種由末梢神經分泌的神經遞質,在骨修復、骨形成和骨吸收中的積極作用表明其在骨折愈合中扮演重要的角色[2]。本課題組在前期研究[3]中已證實,CGRP在頜骨骨折的愈合過程中具有重要的促進作用。最近,CGRP還被證明對成骨細胞具有潛在的保護作用,能夠通過抑制成骨細胞的凋亡從而抑制骨丟失[4]。
自噬(autophagy)是一種廣泛存在于細胞內的膜轉運方式,主要用于遞送不必要的或者不正常的蛋白質和細胞器至溶酶體以降解和再利用[5]。自噬能防止細胞損傷的聚集,并確保細胞內的平衡。凋亡是依賴于半胱氨酸天冬氨酸特異性蛋白酶(cysteinecontaining aspartate-specific proteases,caspase)的自主有序的細胞死亡方式,形態學上主要表現為核固縮、染色質濃聚、核破裂及凋亡小體形成[6]。凋亡與自噬的平衡是維持細胞穩態必不可少的,在骨折愈合過程中常伴隨著凋亡與自噬的發生。在此過程中二者如何互相作用尚不清楚。本課題組在前期研究[3,7]中已證實,CGRP在頜骨骨折的愈合過程中具有重要的促進作用,在前期研究的基礎上,本課題組將研究在饑餓條件下CGRP對成骨細胞凋亡的影響,并首次從自噬入手,試圖進一步揭示CGRP對成骨細胞的保護作用所涉及的機制。
1 材料和方法
1.1 主要試劑和儀器
小鼠MC3T3-E1成骨細胞(購自于中國科學院上海細胞庫),α-MEM培養基、青鏈霉素混合液、胎牛血清(Gibco公司,美國),CGRP、單丹磺酰尸胺(monodansylcadaverin,MDC)、3-甲基腺嘌呤(3-methyladenine,3-MA)(Sigma公司,美國);微管相關蛋白1輕鏈3(microtubule associated protein 1 light chain 3,LC3)、P62蛋白(CST公司,美國),小鼠抗β-actin單抗(北京中杉金橋生物技術有限公司),辣根過氧化物酶標記羊抗兔IgG、辣根過氧化物酶標記羊抗鼠IgG(ABGENT公司,美國),Image Quant LAS 4000數字成像系統(GE公司,美國),MoFlo XDF流式細胞儀(Beckman公司,美國),TCS SP5激光共聚焦顯微鏡(Leica公司,德國)。
1.2 細胞培養
小鼠MC3T3-E1成骨細胞株在含10%胎牛血清和1%青鏈霉素混合液的α-MEM培養基中5%CO2、37 ℃條件下培養,每3 d換液一次,鏡下觀察細胞生長至密度約80%時0.25%胰蛋白酶消化傳代培養。本實驗使用第三代MC3T3-E1成骨細胞。
1.3.1 血清饑餓對MC3T3-E1成骨細胞凋亡和自噬的影響 1)將MC3T3-E1成骨細胞在含血清培養液(空白對照組)、無血清培養液(血清饑餓組)中分別培養12 h和24 h,采用蛋白質印跡(Western blot)檢測LC3蛋白的表達,流式細胞術檢測細胞凋亡。2)將MC3T3-E1成骨細胞分別在含血清培養液(空白對照組)、無血清培養液(血清饑餓組)、5 mmol·L-13-MA預處理1 h+無血清培養液(血清饑餓+3-MA組)中培養24 h,采用流式細胞術檢測細胞凋亡。
1.3.2 血清饑餓下CGRP對MC3T3-E1成骨細胞自噬和凋亡的影響 1)將MC3T3-E1成骨細胞分別在含血清培養液(空白對照組)、無血清培養液(血清饑餓組)、無血清+不同濃度(10-10、10-9、10-8、10-7mol·L-1)CGRP培養液(血清饑餓+CGRP組)中培養24 h,采用Western blot檢測LC3和P62蛋白的表達。2)將MC3T3-E1成骨細胞分別在含血清培養液(空白對照組)、無血清培養液(血清饑餓組)、無血清+10-8mol·L-1CGRP培養液(血清饑餓+CGRP組)培養2、6、12、24、48、72 h,采用Western blot檢測LC3蛋白的表達,流式細胞術檢測培養24 h時的細胞凋亡,MDC染色檢測培養24 h時細胞自噬泡的表達。3)將MC3T3-E1成骨細胞分別在含血清培養液(空白對照組)、無血清培養液(血清饑餓組)、無血清+10-8mol·L-1CGRP培養液(血清饑餓+ CGRP組)、5 mmol·L-13-MA預處理1 h+無血清培養液(血清饑餓+3-MA組)、3-MA預處理1 h+無血清+10-8mol·L-1CGRP培養液(血清饑餓+CGRP+3-MA組)培養24 h,采用流式細胞術檢測細胞凋亡。
1.4 檢測方法
1.4.1 Western blot檢測LC3和P62蛋白的表達 將培養的MC3T3-E1成骨細胞按常規方法裂解,提取細胞總蛋白。BCA法測定蛋白濃度后,十二烷基硫酸鈉聚丙烯酰胺凝膠電泳(sodium dodecyl sulfate polyacrylamide gel electrophoresis,SDS-PAGE)分離,將目的蛋白轉至0.20 μm的PVDG膜,LC3抗體(1︰1 000)、P62抗體(1︰1 000)和β-actin(1︰1 000)4 ℃孵育過夜;辣根過氧化物酶標記二抗(1︰5 000),用電化學發光法發光顯影,Image Quant LAS4000數字成像系統采集影像。
1.4.2 流式細胞術檢測細胞凋亡 將培養的MC3T3-E1成骨細胞以每孔1×106個接種于六孔板中,PBS液重懸細胞后加入1 μg·mL-1Annexin V-PI雙染,采用流式細胞術檢測細胞凋亡,計算凋亡細胞所占百分比。
1.4.3 MDC染色觀察自噬泡 將培養的MC3T3-E1成骨細胞進行爬片處理后,棄培養基,PBS沖洗3遍,4%多聚甲醛固定15 min,PBS洗2次,加入0.05 mmol·L-1MDC染料200 μL,37 ℃孵育10 min,PBS沖洗4遍,共聚焦激光顯微鏡下(激發波長380 nm,發射波長525 nm)觀察細胞自噬泡的變化。
1.5 統計學分析
卡錘頭的原因也比較多,常見的有以下幾種:一是沖孔操作中在經過兩種強度差別明顯的巖層時沒有對行程進行相應調整;二是地層中存在的硬度較大的探頭石將錘頭卡住;三是沖孔過程中由于操作不當使錘頭偏斜頂住孔壁導致錘頭無法順利移動。
采用SPSS 17.0軟件進行統計,數據以均數±標準差表示,Student t檢驗或單因素方差進行分析。
2 結果
2.1 血清饑餓對MC3T3-E1成骨細胞凋亡和自噬的影響
Western blot及流式細胞術檢測結果顯示,無論在12 h還是在24 h,與空白對照組相比,血清饑餓組LC3Ⅱ蛋白表達及細胞凋亡均增加(P<0.01),且隨著時間延長,LC3Ⅱ/Ⅰ比值差異加大(圖1~3)。
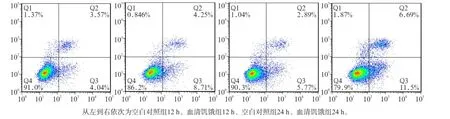
圖1 2組MC3T3-E1成骨細胞培養12、24 h的細胞凋亡水平Fig1 Apoptosis of MC3T3-E1 osteoblasts of two groups in the time point of 12 h and 24 h

圖2 2組MC3T3-E1成骨細胞培養12、24 h的細胞凋亡率Fig2 Apoptosis rate of MC3T3-E1 osteoblasts of two groups in the time point of 12 h and 24 h

圖3 2組MC3T3-E1成骨細胞培養12、24 h的LC3蛋白表達水平Fig3 Expression of LC3 of MC3T3-E1 osteoblasts of two groups in the time point of 12 h and 24 h
這提示,血清饑餓可以誘導MC3T3-E1成骨細胞的凋亡和自噬。
3-MA預處理后無血清培養液培養24 h,流式細胞術結果顯示MC3T3-E1成骨細胞的凋亡較血清饑餓組明顯增加(P<0.01,圖4、5)。這提示血清饑餓條件下,自噬的抑制會導致MC3T3-E1成骨細胞的凋亡增強。

圖4 3組MC3T3-E1成骨細胞培養24 h的細胞凋亡水平Fig4 Apoptosis of MC3T3-E1 osteoblasts of three groups in the time point of 24 h
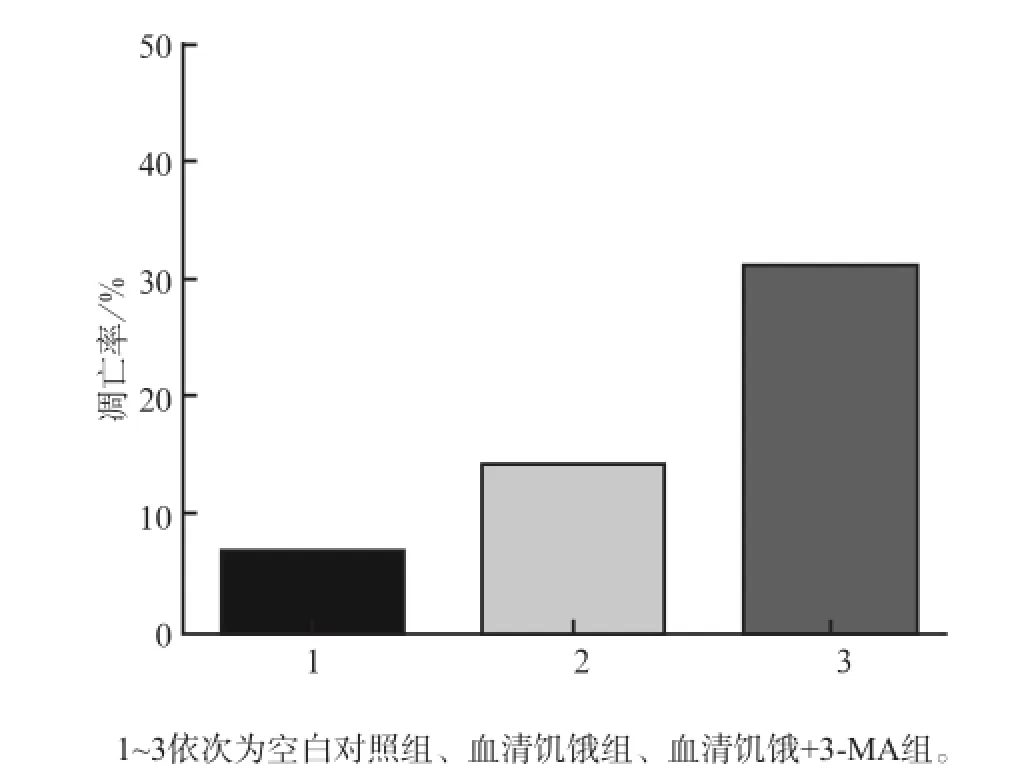
圖5 3組MC3T3-E1成骨細胞培養24 h的細胞凋亡率Fig5 Apoptosis rate of MC3T3-E1 osteoblasts of three groups in the time point of 24 h
2.2 血清饑餓下CGRP對MC3T3-E1成骨細胞自噬的影響
Western blot檢測結果表明,與空白對照組相比,血清饑餓組和不同濃度的血清饑餓+CGRP組MC3T3-E1成骨細胞培養24 h后LC3Ⅱ蛋白的表達均增加,在CGRP濃度為10-8mol·L-1時LC3Ⅱ/Ⅰ比值最高;P62蛋白的表達均降低,CGRP濃度為10-8mol·L-1為最大刺激濃度(圖6)。這表明,在血清饑餓的條件下,CGRP能促進MC3T3-E1成骨細胞的自噬,10-8mol·L-1為最適濃度。
相同濃度(10-8mol·L-1)CGRP作用MC3T3-E1成骨細胞不同時間(2,6,12,24,48,72 h)之間相比較,24 h時LC3Ⅱ/Ⅰ比值最高(圖7),提示在作用24 h時CGRP能顯著誘導MC3T3-E1成骨細胞的自噬。

圖6 各組MC3T3-E1成骨細胞培養24 h的LC3、P62蛋白表達水平Fig6 Expression of LC3 and P62 of MC3T3-E1 osteoblasts of each group in the time point of 24 h

圖7 各組MC3T3-E1成骨細胞培養不同時間的LC3蛋白表達水平Fig7 Expression of LC3 of MC3T3-E1 osteoblasts of each group at different time
2.3 血清饑餓下CGRP對MC3T3-E1成骨細胞凋亡的影響
流式細胞術檢測結果表明,血清饑餓+CGRP組能抑制MC3T3-E1成骨細胞的凋亡(P<0.05)(圖8);MDC染色檢測結果顯示,血清饑餓組的自噬囊泡熒光強度較空白對照組強,血清饑餓+CGRP組較血清饑餓組強(圖9)。

圖8 3組MC3T3-E1成骨細胞培養24 h的細胞凋亡率Fig8 Apoptosis rate of MC3T3-E1 osteoblasts of three groups in the time point of 24 h
這說明,在饑餓條件下,CGRP不僅能夠抑制MC3T3-E1成骨細胞的凋亡,同時也能夠促進MC3T3-E1成骨細胞自噬泡的合成。
2.4 血清饑餓下3-MA預處理后CGRP對MC3T3-E1成骨細胞自噬和凋亡的影響
流式細胞術檢測的結果表明,3-MA預處理后MC3T3-E1成骨細胞凋亡增加,CGRP部分逆轉3-MA預處理所增加的血清饑餓條件下MC3T3-E1成骨細胞凋亡的水平(圖10)。該結果提示CGRP可能通過促進MC3T3-E1成骨細胞的自噬從而抑制細胞的凋亡。

圖9 3組MC3T3-E1成骨細胞培養24 h的細胞自噬泡 MDC × 400Fig9 Autophagosome of MC3T3-E1 osteoblasts of three groups in the time point of 24 h MDC × 400

圖10 5組MC3T3-E1成骨細胞培養24 h的細胞凋亡率Fig10 Apoptosis rate of MC3T3-E1 osteoblasts of five groups in the time point of 24 h
3 討論
CGRP主要來源于感覺神經肽,是骨組織中分泌最廣泛的一種神經遞質。本課題組前期研究[8-9]發現,CGRP能夠促進骨折的愈合,也證實CGRP能促進成骨細胞的活性。作為骨修復與骨重建的活化劑,CGRP的功能引起越來越多的關注。新近的研究發現CGRP能夠抑制成骨細胞的凋亡,但這種抗凋亡的保護作用的具體機制少有報道。
自噬是一個循環的過程,通過降解受損的大分子物質及細胞器實現細胞本身的代謝需求和細胞器的更新,是細胞的一種生存機制[10]。同時,自噬也參與細胞的各種生理功能,Nollet等[11]研究發現,自噬參與成骨細胞的礦化過程,特異性成骨細胞自噬缺陷的小鼠的骨量會發生明顯的減少,提示成骨細胞的自噬同時涉及到骨礦化和骨平衡。在骨折愈合和自噬的研究[12-13]中發現,在手術致大鼠骨折的損傷初期階段LC3-Ⅱ與增殖細胞核抗體蛋白的表達都有明顯的升高,并且兩者之間存在相互關系;自噬增強劑雷帕霉素處理大鼠所構造的股骨骨折模型時能增加血管內皮生長因子和增殖細胞核抗體的表達水平,并且能顯著增加成骨細胞數量和礦化結痂,提示自噬對骨折愈合可能具有潛在的積極作用。
細胞凋亡是細胞程序性死亡的過程,過度的細胞凋亡會引起組織的壞死或修復的減少,在許多疾病和損傷的機制中具有重要的生物學作用。凋亡和自噬這二者之間在功能上既存在廣泛的聯系,又有著明顯的不同。自噬為凋亡所需,自噬通常發生在凋亡之前,進而啟動凋亡;自噬也可抑制凋亡,保護細胞免于發生凋亡和壞死;自噬可以向凋亡轉化,共同促進細胞死亡[6,14]。
本實驗首先研究了血清饑餓對MC3T3-E1成骨細胞凋亡和自噬的影響,并初步探索了二者之間的相互關系。實驗結果表明,血清饑餓會誘導MC3T3-E1成骨細胞的凋亡,同時也促進了MC3T3-E1成骨細胞的自噬。此外,通過自噬抑制劑3-MA的應用,本研究發現當抑制MC3T3-E1成骨細胞的自噬時,細胞的凋亡會明顯增加,這說明自噬的存在可能會抑制MC3T3-E1成骨細胞凋亡的發生。接下來,本實驗又研究了CGRP對MC3T3-E1成骨細胞自噬/凋亡的影響,結果表明,血清饑餓條件下,CGRP增強了細胞自噬并且抑制了MC3T3-E1成骨細胞的凋亡。進一步聯用自噬抑制劑研究發現,CGRP對血清饑餓條件下MC3T3-E1成骨細胞凋亡的抑制作用被明顯降低;這也反向說明了細胞自噬活性在CGRP對MC3T3-E1成骨細胞的保護中的重要性,這為CGRP在骨修復中的應用前景提供了新的證據,也為進一步揭示CGRP對成骨細胞的保護作用所涉及的機制提供了新的思路。
[1] Lamprou M, Kaspiris A, Panagiotopoulos E, et al. The role of pleiotrophin in bone repair[J]. Injury, 2014, 45(12):1816-1823.
[2] He H, Chai J, Zhang S, et al. CGRP May regulate bone metabolism through stimulating osteoblast differentiation and inhibiting osteoclast formation[J]. Mol Med Rep, 2016, 13 (5):3977-3984.
[3] Yan L, Yinghui T, Bo Y, et al. Effect of calcitonin generelated peptide on nitric oxide production in osteoblasts: an experimental study[J]. Cell Biol Int, 2011, 35(8):757-765.
[4] Mrak E, Guidobono F, Moro G, et al. Calcitonin gene-related peptide (CGRP) inhibits apoptosis in human osteoblasts by β-catenin stabilization[J]. J Cell Physiol, 2010, 225(3):701-708.
[5] Kang C, You YJ, Avery L. Dual roles of autophagy in the survival of Caenorhabditis elegans during starvation[J]. Genes Dev, 2007, 21(17):2161-2171.
[6] 朱文胤, 陳倩倩, 劉祁, 等. 饑餓誘導下小鼠成骨細胞自噬與凋亡相互作用研究[J]. 川北醫學院學報, 2015, 30(2): 206-211. Zhu WY, Chen QQ, Liu Q, et al. Study on the interaction between autophagy and apoptosis of MC3 T3-E1 at starvation state[J]. J North Sichuan Med Col, 2015, 30(2):206-211.
[7] Zhao Z, Fu X, Zhang G, et al. The influence of RAMP1 overexpression on CGRP-induced osteogenic differentiation in MG-63 cells in vitro: an experimental study[J]. J Cell Biochem, 2013, 114(2):314-322.
[8] 王飛, 張慧宇, 竇予昕, 等. 降鈣素基因相關肽通過Hippo通路調控小鼠骨髓間充質干細胞成骨分化的實驗研究[J]. 華西口腔醫學雜志, 2016, 34(3):286-290.
Wang F, Zhang HY, Dou YX, et al. Calcitonin gene-related peptide-induced osteogenic differentiation of mouse bone marrow stromal cells through Hippo pathway in vitro[J]. West Chin J Stomatol, 2016, 34(3):286-290.
[9] 蔡俊, 呂俊, 李適庭, 等. 降鈣素基因相關肽通過抑制Nod樣受體蛋白3表達促進小鼠成骨細胞分化的研究[J]. 華西口腔醫學雜志, 2016, 34(1):12-16.
Cai J, Lü J, Li ST, et al. Calcitonin gene-related peptide inhibits the expression of Nod-like receptor protein 3 to promote osteoblast differen-tiation in mouse osteoblasts in vitro [J]. West Chin J Stomatol, 2016, 34(1):12-16.
[10] Mizushima N, Komatsu M. Autophagy: renovation of cells and tissues[J]. Cell, 2011, 147(4):728-741.
[11] Nollet M, Santucci-Darmanin S, Breuil V, et al. Autophagy in osteoblasts is involved in mineralization and bone homeostasis[J]. Autophagy, 2014, 10(11):1965-1977.
[12] Zhou Q, Luo D, Li T, et al. Bone fracture in a rat femoral fracture model is associated with the activation of autophagy [J]. Exp Ther Med, 2015, 10(5):1675-1680.
[13] Yang GE, Duan X, Lin D, et al. Rapamycin-induced autophagy activity promotes bone fracture healing in rats[J]. Exp Ther Med, 2015, 10(4):1327-1333.
[14] Yang YH, Chen K, Li B, et al. Estradiol inhibits osteoblast apoptosis via promotion of autophagy through the ER-ERK-mTOR pathway[J]. Apoptosis, 2013, 18(11):1363-1375.
(本文采編 李彩)
Effect of calcitonin gene-related peptide on MC3T3-E1 osteoblast apoptosis and autophagy induced by serum starvation
An Yang, Zhang Huiyu, Guo Junfeng, Li Xin, Yang Yang, Zhang Gang,Tan Yinghui. (Dept. of Oral and Maxillofacial Surgery, Xinqiao Hospital, The Third Military Medical University, Chongqing 400037, China)
Objective To study the effect of calcitonin gene-related peptide (CGRP) on apoptosis and autophagy of mouse MC3T3-E1 osteoblast and their interaction and to further clarify protective mechanism of CGRP on osteoblasts. Methods MC3T3-E1 osteoblasts of mouse were cultured in vitro. Western blot and flow cytometry were used to detect expressions of microtubule-associated protein 1 light chain 3 (LC3) and P62 protein of MC3T3-E1 osteoblasts cultured with serum culture and serum-free (serum starvation) culture. Western blot was also used to detect expressions of LC3 and P62 protein of MC3T3-E1 osteoblast cultured at different concentrations (10-10, 10-9, 10-8, and 10-7mol·L-1) or without added CGRP. MC3T3-E1 osteoblasts were treated with 10-8mol·L-1CGRP at different times (2, 6, 12, 24, 48, and 72 h), protein expression levels of LC3 were assessed by Western blot and flow cytometry, and changes in autophagosome in cells were detected by monodansylcadaverin staining. Autophagy inhibitor 3-methyladenine (3-MA) was used to pretreat MC3T3-E1 osteoblasts. Cells were then treated with or without CGRP for 24 h. Flow cytometry was used to detect apoptosis level.ResultsUnder serum starvation conditions, LC3Ⅱ expression and apoptosis of osteoblasts increased compared with that of serum culture. Under 3-MApretreatment and serum starvation conditions, LC3Ⅱ expression of osteoblasts increased compared with that of serum culture (P<0.01). Compared with serum culture, serum starvation culture with or without CGRP significantly increased expression level of LC3 and reduced expression level of P62. LC3Ⅱ/Ⅰ of osteoblasts was the highest under serum starvation and 10-8mol·L-1CGRP conditions. Serum starvation and 10-8mol·L-1CGRP culture inhibited apoptosis of osteoblasts and promoted synthesis of autophagosome. Apoptosis of osteoblasts increased after 3-MA pretreatment, and CGRP reversed inhibitory effects of 3-MA CGRP on apoptosis.ConclusionCGRP can increase autophagy of MC3T3-E1 osteoblasts under serum starvation conditions. CGRP may also inhibit apoptosis of MC3T3-E1 osteoblasts by promoting autophagy.
calcitonin gene-related peptide; osteoblast; autophagy; apoptosis
Q 25
A
10.7518/hxkq.2017.02.005
Supported by: The National Natural Science Foundation of China (81371110). Correspondence: Tan Yinghui, E-mail: tanyhxqkq@163.com.
2016-08-16;
2016-10-10
國家自然科學基金(81371110)
安洋,碩士,E-mail:anyangqq@163.com
譚穎徽,教授,博士,E-mail:tanyhxqkq@163.com
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中老年保健(2021年3期)2021-08-22 06:50:04
天津醫科大學學報(2021年2期)2021-03-29 05:31:08
現代臨床醫學(2021年1期)2021-01-26 00:56:02
學苑創造·A版(2020年9期)2020-10-13 09:41:02
海峽科技與產業(2016年3期)2016-05-17 04:32:12
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
