澄江動物群研究與進展*
2017-05-12 06:22:40段艷紅盧龍斗
生物學通報 2017年3期
段艷紅 文 博 盧龍斗
(1新鄉學院生命科學技術學院 河南新鄉 453003 2華中科技大學同濟醫學院附屬同濟醫院 湖北武漢 430030 3新鄉醫學院三全學院生命科學技術學院 河南新鄉 453003)
1 云南澄江動物群
在寒武紀早期的古海洋中,存在著地球生命歷史中發展速度最快、規模最大、影響最為深遠的動物生命演化創新事件。中國云南省澄江動物群(圖1)和貴州省凱里動物群的發現,為中國古生物學家探討寒武紀生命大爆發提供了新證據和新材料[1-3]。云南澄江動物群在年代上較加拿大布爾吉斯頁巖軟軀體動物群還要早1 500萬年,而中國貴州凱里動物群和布爾吉斯頁巖軟軀體動物群在產生年代上相當,因此其發現意義十分重大。

圖1 云南澄江動物群化石發現點
云南澄江動物群的化石廣泛分布在云南省澄江馬鞍山剖面和澄江小濫田剖面等地,澄江生物化石產地約有 39 個以上[4](圖1,表1)。 其中安寧(已發現14生物門類的79個物種)、海口(已發現14生物門類的156個物種)和澄江(已發現24個生物門類的228個物種)是澄江動物群化石的3個主要產地,在細粒的薄層泥巖中完美地保存了軟軀體動物化石(圖2),因此具有極高的研究價值和觀賞價值。

表1 澄江動物群化石產地分布
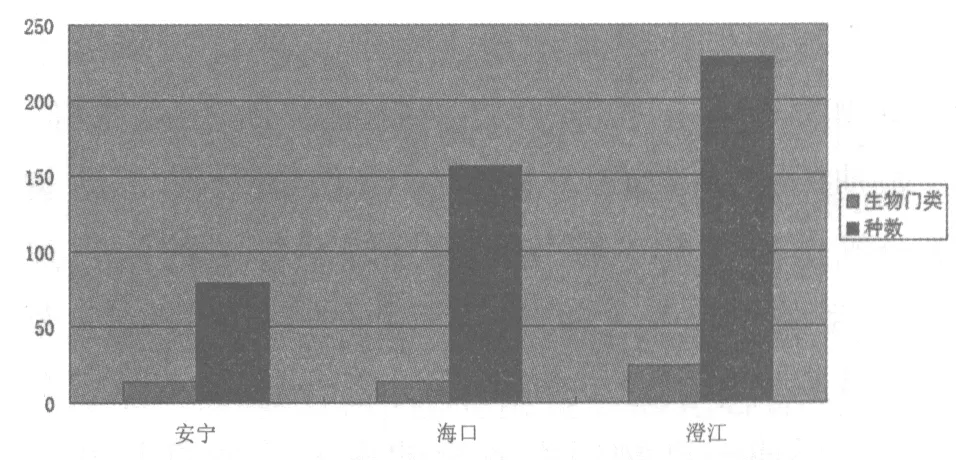
圖2 澄江動物群3個主要化石產地生物組成的比較
澄江動物化石群所處的沉積環境是淺海軟泥質基底的大陸架環境[5-6](圖3,圖4)。當時動物的生活類型有浮游動物、底表棲固著動物、內棲動物和游泳動物4種類型。取食方式有食泥、濾食、食腐或捕食、雜食和食草等類型;且具有多層次營養級的金字塔式食物網和交叉的食物鏈在寒武紀早期的澄江動物化石群已經形成[4]。澄江動物化石群的3個主要產地是安寧、海口、澄江,這3個地方生物物種的分異度較高[4],節肢動物在這3個產地均占據著絕對主導地位,屬于優勢動物類群,十分發達。

圖3 澄江動物群化石采集點之一

圖4 野外發現的澄江動物群部分化石
澄江動物群軟軀體保存化石的發現震撼了全世界,引發了轟動效應,它在時間上較布爾吉斯頁巖動物群還要早1 500萬年,無論在化石門類、化石面貌、化石數量、化石保存狀態、化石形態和結構及化石在生物演化地位上意義均要優于埃迪卡拉動物群和布爾吉斯頁巖動物群及凱里動物群。從埃迪卡拉動物群→梅樹村動物群→澄江動物群→凱里動物群→布爾吉斯頁巖動物群的演化過程中,動物的生命形式是從無數的實驗失敗中,獲得了最為簡單的形式和軀體構型→復雜的形式和結構的形成→動物門類齊全的爆發式的漫長和漸變過程如帚刺奇蝦、延長撫仙湖蟲、中華微網蟲、純真滇東貝、長形澄江蝦、寬跨馬蟲、朵氏小昆明蟲、始祖長江海鞘和鳳嬌昆明魚等。
2 澄江動物化石群的發現和生物組成
1984年,中國古生物學家侯先光在云南省澄江縣帽天山發現了保存軟軀體化石納羅蟲(Naraoia)的云南省澄江動物化石群(相當于國際標準界線層型的寒武紀第2統第3階)。1985年,張文堂和侯先光首次發表和命名了納羅蟲[7-8]。澄江動物化石群后來又在呈貢、馬龍、武定、曲靖等39個產地被發現[4]。接著又陸續發現大量其他軟軀體的無脊椎動物和脊椎動物化石[4,6]。截止至2016年4月,云南澄江動物化石群中至少發現了192屬228種生物化石(圖4,圖5),主要包括海綿動物、櫛水母動物、帚蟲動物、內肛動物、線形蟲動物、奇蝦類動物、毛顎動物、軟體動物、節肢動物、古蟲動物、頭索動物、開腔骨類動物等,屬于18個分類單元和24 個生物門類[4,8]。 澄江動物化石群無論是在動物化石組成和化石數量上,還是在保存質量上均可與加拿大布爾吉斯頁巖動物群中的動物組成和動物面貌相媲美,甚至略勝一籌。澄江動物化石群的動物組成反映了早寒武世赤道附近淺海軟泥質基底生活的古動物群落的生命大爆發[8-9]。
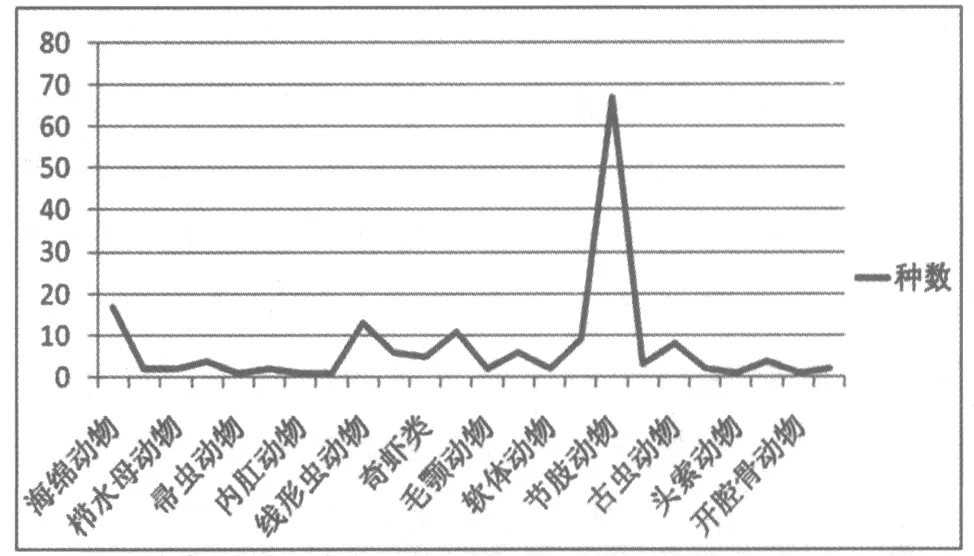
圖5 云南澄江動物群動物化石組成面貌
澄江動物化石群的生物多樣化和特殊的保存質量可以與布爾吉斯頁巖動物群相媲美。其中,與布爾吉斯頁巖化石動物群相同屬有18屬,分別是奇蝦(Anomalocaris)、鰓蝦蟲(Branchiocaris)、加拿大蟲(Canadaspis)、斗蓬海綿(Choia)、足杯蟲(Dinomischus)、伊爾東缽(Eldonia)、怪誕蟲(Hallusigenia)、海扎海綿(Hazelia)、等刺蟲(Isoxys)、林喬利蟲(Leanchoilia)、小細絲海綿(Leptomitella)、細絲海綿(Leptomitus)、納羅蟲(Naraoia)、奧代雷蟲(Ordaraia)、西德尼蟲(Sidneyia)、吐卓蟲(Tuzoia)、塞爾扣克蟲(Selkirkia)、塔卡瓦海綿(Takakkwia)。 澄江生物化石群與凱里生物化石群相同屬有7屬,分別是奇蝦(Anomalocaris)、等刺蟲(Isoxys)、微網蟲(Microdictyon)、納羅蟲(Naraoia)、古蠕蟲(Palaeoscolex)、尾頭蟲(Urocodia)、吐卓蟲(Tuzoia)等。許多動物在澄江動物群中首次出現,因此,在寒武紀大爆發過程中它起到了“源頭”和起源的作用,而凱里動物群和布爾吉斯頁巖動物群及京山石龍洞組動物群是澄江動物化石群的延伸和擴展,所以只是寒武紀大爆發的一段“流程”而已[6]。
3 澄江動物群化石的埋藏模式
澄江動物群的化石埋藏模式表現為:①寒武紀古海洋的大規模海侵事件(即海水侵入陸地),造成海水周期性貧氧或缺氧,導致大量動物死亡,并且迅速被泥沙埋藏而保存下來成為動物化石;②由風暴引起的懸浮泥質沉積物(即懸濁物)造成動物窒息死亡并且快速埋藏而被保存下來成為化石,例如節肢動物朵氏小昆明蟲[6,10]和翻吻動物[11];③由風從陸地帶來的泥質沉積物(即陸源碎屑)引起的快速沉積埋藏形成化石等[11-13]。
在寒武紀澄江動物群還有一些比較常見的動物化石埋藏的例子,例如異地生物經過泥質風暴流由搬運引起的快速埋藏的微網蟲和等刺蟲(可以保護動物不受捕食者所食,以至于后來形成化石,屬于異地埋藏);古生物學家韓健對澄江化石庫翻吻動物進行深入研究和探索時,發現似腕足動物附著在蠕蟲身體的表面和大量蠕蟲動物的遺跡化石,這是澄江化石庫原地埋藏的證據和佐證[12-15]。朱茂炎等(2001)認為澄江動物群的沉積環境為遠基濱外——深水濱前環境,保存了豐富的軟軀體動物化石。
澄江動物群的動物化石以原地埋藏和異地埋藏2種方式共存,使得古生物學家早就發現翻吻動物在早中寒武世獲得了空前的發展和演化,顯示了其形態和軀體構型方面的高分異度和趨異度,因此,在生活環境、運動模式和埋藏方式上也有了很大變化[12],由此推測在翻吻動物和翻吻動物之間可能也存在著捕食與被捕食的互動關系,并在寒武紀以后得到了進化和發展。
4 造成澄江動物群的化石動物多樣化的原因
古生物學界認為,澄江動物群保存如此豐富和多樣化的動物化石類型,主要原因在于:在寒武紀海洋災難到來時,動物被快速埋葬,使得各種軟軀體組織(例如表皮、肌肉、眼、眼瘤、附肢、卵和神經系統等)得以充分地保存下來而成為精美的動物化石[12-14];不同化石產地的動物組成面貌大體一致,優勢類群以節肢動物和海綿動物為主[5-6,15]。各動物化石產地保存的環境不同,會有不同層位的優勢種和特征種保存下來。例如在云南省昆明市晉寧縣玉案山段產生出大量的埃迪卡拉型動物[7],但是,在云南其他地區卻未見報道,可將之視作該地的特征種。原地埋藏和異地的快速埋藏可能是澄江生物群的化石得以保存的重要原因[5-6]。在布爾吉斯頁巖動物群與澄江動物群以及凱里動物群中,軟軀體動物化石的組成面貌是極其類似的,但是,布爾吉斯頁巖動物群和凱里動物群的動物軀體構型的分異度均要低于澄江動物群[1]。
寒武紀生命大爆發從云南省晉寧縣梅樹村小殼動物群開始,經過節肢動物的大發展到澄江動物群的門一級構型完成,是一個分階段和分層次的具有連續性的動物演化事件,澄江動物群處于寒武紀大爆發的“源頭”地位,而中寒武世貴州凱里動物群、湖北京山石龍洞組動物群和加拿大布爾吉斯頁巖動物群是寒武紀生命大爆發完成后的延續和演化及“流程”[1]。從整個動物的演化歷史來看,生命演化的過程中既有漸進又有爆發、既有漸變又有突變、既有漸滅又有絕滅、既有生存斗爭優勝劣汰又有協調生存和協同演化。寒武紀生命大爆發理論的誕生是對達爾文進化論的補充、修正和完善。在澄江動物化石群還出現了一些巨大的和高級的、處于頂層的食肉動物如奇蝦類,表明復雜的生態系統在早寒武世已經建立,以后有了更大的發展。
5 澄江動物群研究新進展
澄江動物群的研究新進展主要體現在:①揭示了寒武紀生命大爆發“主幕”的全貌,解釋和回答了寒武紀大爆發這個令達爾文當年也困惑不解的疑難問題;②探索了甲殼動物、螯肢動物、真節肢動物和脊椎動物的起源問題;③證實了現代動物復雜的生態系統、食物鏈和食物網起源于寒武紀;④補充了自下而上的倒錐形進化理論模型,被進化生物學家普遍認為是自上而下的爆炸式或蘑菇云式的進化模型,并且提供充分的化石證據,進行論證;⑤建立了無脊椎動物向脊椎動物進化的“五步走”假說,包括第1步是寒武紀云南澄江動物化石群中的后口動物“皇冠西大動物(1998年舒德干院士等人發現后以西北大學命名)”,第2步是半索動物“海口蟲”和“云南蟲”,它們是古蟲的近親,古蟲動物門是從原口動物向后口動物演化的關鍵環節,第3步是最古老的尾索動物“始祖長江海鞘”,它有與現生帚蟲動物的觸手冠類似的結構,第4步是似頭索動物“海口華夏鰻”和“中間型中新魚”,第5步是脊椎動物“海口魚”和“鳳姣昆明魚”,從而論證了從無脊椎后口動物向脊椎動物演化過渡的各種中間類型和過渡環節,勾勒出一幅較為完整的早期生命演化譜系,顯示了中國是脊椎動物起源的搖籃,這些研究進展是對達爾文進化論的補充、修正和發展,也對達爾文進化論及其發展做出了更加重大的貢獻。
[1]羅惠麟,胡世學,陳良忠.昆明地區早寒武世澄江動物群.昆明:云南科技出版社,1999:12.
[2]侯先光,馮向紅.澄江生物化石群.生物學通報,1999,34(12):6.
[3]朱立祥.“生物進化:II”的教學組織.生物學通報,2010,45(1):24.
[4]趙方臣,朱茂炎,胡世學.云南寒武紀早期澄江動物群古群落分析.中國科學:地球科學,2010,40(9):1135.
[5]段艷紅,韓健,張志飛,等.華南布爾吉斯頁巖型軟軀體化石生物群對比研究.西北大學學報(自然科學版),2012,42(2):288.
[6]段艷紅.寒武紀大爆發后生動物的一些探討:以澄江化石庫為例.西安:西北大學博士學位論文,2013:12.
[7]Shu Degan,Conway Morris,Han Jian,et al.Lower Cambrian Vendobionts from China and Early Diploblast Evolution.Science,2006,312(5774):731.
[8]張文堂,侯先光.Naraoia在亞洲大陸的發現.古生物學報,1985,24(6):591.
[9]Zhang X L,Liu W,Zhao Y L.Cambrian Burgess Shale-type Lagerstatten in South China:distribution and significance.Gondwana Research,2008(14):255.
[10]段艷紅.朵氏小昆明蟲的攜卵行為.生物學通報,2015,50(7):10.
[11]韓健,張志飛,劉建妮.澄江動物群中翻吻動物的埋藏學與生態學研究.西北大學學報(自然科學版),2004,34(2):208.
[12]Zhang X L,Shu D G,LI Y,et al.New sites of Chengjiang fossils:crucial windows on the Cambrian explosion.Journal of the Geological Society,2001(158):211.
[13]Duan Y H,Han J,Fu D J,et al.Reproductive strategy of the bradoriids arthropod Kunmingella douvillei from the Lower Cambrian Chengjiang Lagerst?tte,South Chian.Gondwana Research,2014(25):983.
[14]劉年方.化石保存方式.生物學通報,1997,32(5):26.
[15]Han J,Zhang Z F,Liu J N.A preliminary note on the dispersal of the Cambrian Burgess Shale-type faunas.Gondwana Research,2008(14):269.
(E-mail:dyh6065@153.com)
