米曲霉生產(chǎn)曲酸的發(fā)酵條件研究
2017-05-17 16:51:23劉容孫衛(wèi)東李軍委
廣西農(nóng)學報 2016年6期
劉容+孫衛(wèi)東+李軍委

摘要:【目的】優(yōu)化米曲霉生產(chǎn)曲酸的發(fā)酵條件,提高生產(chǎn)效率,對米曲霉(Aspergillus oryzae)GIM3.423生產(chǎn)曲酸的發(fā)酵條件進行研究。【方法】采用單因素試驗對搖瓶發(fā)酵條件進行優(yōu)化。【結(jié)果】發(fā)酵液pH、發(fā)酵溫度、裝液量、搖床轉(zhuǎn)速、接種量等因素影響和制約整個發(fā)酵過程,發(fā)酵溫度30℃,裝液量60ml/250ml,搖床轉(zhuǎn)速200rpm,種齡6h,接種量10%,發(fā)酵周期控制在13d條件下,曲酸生產(chǎn)的轉(zhuǎn)化率可達23.6%,。【結(jié)論】搖瓶發(fā)酵最佳條件為發(fā)酵溫度30℃,裝液量60ml/250ml,搖床轉(zhuǎn)速200rpm,種齡6h,接種量10%,發(fā)酵周期控制在13d。
關(guān)鍵詞:曲酸;米曲霉;發(fā)酵條件
中圖分類號:TQ925 文獻標識碼:A 文章編號:1003-4374(2016)06-0025-05
Abstract:【Objective】 In order to optimize the fermentation conditions for producing kojic acid by aspergillus oryzae, and improve production efficiency, in this paper, the fermentation conditions for production of kojic acid by Aspergillus oryzae GIM3.423 have been studied. 【Method】 Shaking flask fermentation was optimized by using single factors test.【Conclusion】The optimum conditions were identified as follow: temperature 30℃,liquid-loading capacity 60mL/250mL,revolution fermentation yield was improved.
Key words:kojic acid;aspergillus oryzae;fermentation condition
曲酸(Kojic acid),化學名為5-羥基-2-羥甲基-1,4-吡喃酮,是一種重要的微生物合成有機酸,存在于醬油、豆瓣醬、酒類的釀造中,在許多以曲霉發(fā)酵的發(fā)酵產(chǎn)品中都可以檢測到曲酸的存在。曲酸具有抗菌、抑菌,抑制酪氨酸酶活性,螯合金屬離子,抗細胞增殖,增強免疫細胞活力,清除自由基等生物活性[1-3],被廣泛地應用在食品、化妝品、農(nóng)業(yè)等領(lǐng)域極具開發(fā)潛力和應用前景[4-5]。
隨著曲酸及其衍生物的應用范圍不斷擴大,其市場需求也不斷增加,提高曲酸的生產(chǎn)效率,降低生產(chǎn)成本成為當前的主要的研究課題。本文以曲酸產(chǎn)生菌米曲霉為菌種,以來源廣的蔗糖為原料進行游離態(tài)發(fā)酵試驗,研究并優(yōu)化曲酸發(fā)酵工藝條件,以期提高曲酸轉(zhuǎn)化率和生產(chǎn)效率,降低生產(chǎn)成本。
1 材料與方法
1.1 材料
1.1.1 菌種 米曲霉(Aspergillus oryzae)GIM3.423,購于廣州微生物研究所微生物菌種保藏中心。
1.1.2 培養(yǎng)基 種子培養(yǎng)基(g/L):蔗糖100,酵母膏6,KH2PO4 5,MgSO4·7 H2O 0.5,吐溫(-80)0.01,pH6.0,121℃濕熱滅菌15min。
發(fā)酵培養(yǎng)基(g/L):按照前期研究優(yōu)化的培養(yǎng)基配比,蔗糖8%,酵母膏0.3%,乙醇2%,KH2PO4 0.1%,MgSO4·7 H2O 0.05%,KCl 0.05%,121℃濕熱滅菌15min。
1.1.3 主要試劑 曲酸,AR,上海源葉生物科技有限公司;3,5-二硝基水楊酸,CP,中國湖州菱湖精細化廠;蔗糖,AR,國藥化學試劑有限公司;乙醇,AR,天津市大茂化學試劑廠。
DNS試劑:分別稱取DNS(3,5-二硝基水楊酸)10.00g,氫氧化鈉16.00g,苯酚5.00g,酒石酸鉀鈉300.00g,依次加入到蒸餾水中溶解,定容到1000mL,避光保存一周,繪制葡萄糖標準曲線。
FeCl3顯色劑:以濃度為0.1% FeCl3-HCl作為曲酸顯色劑:準確稱取10.000gFeCl3用蒸餾水溶解添加到1000mL容量瓶中,量取22.5mL濃鹽酸緩慢加入到容量瓶中,以蒸餾水定容,搖勻,保存?zhèn)溆谩?/p>
1.2 主要儀器
紫外可見分光光度(UV-2802H),尤尼柯(上海)儀器有限公司;全溫度恒溫培養(yǎng)振蕩器ZHWY-Z11C,上海智誠分析儀器制造有限公司;低速大容量多管離心機LXJ-Ⅱ,成都欣宇實驗設(shè)備有限公司;DELTA 320 pH計,梅特勒-托利多儀器(上海)有限公司。
1.3 方法
1.3.1 發(fā)酵實驗。
(1)孢子懸液的制備。將平展米曲霉GIM3.446接到PDF培養(yǎng)基上活化數(shù)次,使菌種活力完全恢復,無菌生理鹽水(含有0.01%土溫-80)將培養(yǎng)4天菌種的孢子沖洗下來,制成濃度為1.0×107個/mL的孢子懸液,4℃保存?zhèn)溆谩?/p>
(2)種子培養(yǎng)。按4%的接種量接種將孢子懸液接種到50mL/250mL種子培養(yǎng)基中,30℃,200rap搖瓶培養(yǎng)24h。
(3)發(fā)酵培養(yǎng)。種子液按照10%的接種量接種到50mL/250mL發(fā)酵培養(yǎng)基中,30℃,200rap搖瓶培養(yǎng)12d檢測殘?zhí)橇俊⑶岷亢蜕锪俊?/p>
1.3.2 分析方法。
(1)孢子濃度的測定。血球計數(shù)板直接計數(shù)。
(2)曲酸含量的測定。采用FeCl3比色法測定發(fā)酵液中曲酸含量[2]。曲酸標準曲線的繪制:準確稱取曲酸1g溶解于蒸餾水中,定容至1000mL,搖勻,制得濃度為1mg/mL的曲酸溶液。分別準確移取0.5mL~4.5mL溶液分別添加得到100mL的容量瓶中,分別加入2mL顯色劑,蒸餾水定容至100mL,搖勻。以蒸餾水為空白對照,測定溶液在500nm波長下的吸光值(OD500),以O(shè)D500為縱坐標,以曲酸含量(mg)為橫坐標,根據(jù)測定結(jié)果繪制出標準曲線。如圖1標準曲線方程為Y=0.0839x+0.0003(R2=0.9999)。
發(fā)酵液中曲酸含量的測定:發(fā)酵液經(jīng)抽濾除掉菌體后,準確移取0.2~1mL濾液到100mL容量瓶中,加入FeCl3顯色劑2mL,蒸餾水定容,搖勻,測定溶液在500nm波長下的吸光度(OD500),根據(jù)標準曲線計算曲酸含量。
(3)殘?zhí)堑臏y定。發(fā)酵液殘?zhí)遣捎肈NS方法測定[3]。
葡萄糖標準曲線的繪制:采用DNS顯色法繪制葡萄糖的標準曲線,取6支25mL具塞刻度試管,分別加入0mL~1mL濃度為2mg/mL葡萄糖標準溶液、1mL~0mL蒸餾水和1.5m3,5-二硝基水楊酸溶液(DNS),搖勻,配成葡萄糖含量不同的反應液。將反應液在沸水浴中加熱5min后取出,在自來水中迅速冷卻至室溫,用蒸餾水定容至25mL,搖勻后測定在540nm波長下的吸光值(OD540),以O(shè)D540為縱坐標,葡萄糖含量(mg)為橫坐標,根據(jù)測定結(jié)果繪制標準曲線。如圖2所示,Y=0.5466x+0.0269(R2=0.9947)。
發(fā)酵液殘?zhí)堑臏y定:準確移取1mL發(fā)酵慮液到100mL容量瓶中,蒸餾水定容。取稀釋液1mL到25mL具塞刻度試管中,后續(xù)操作同上。最后根據(jù)標準曲線計算出發(fā)酵液中殘?zhí)呛俊?/p>
(4)生物量的測定。將分析濾紙放置于105℃下干燥至恒重,用于過濾發(fā)酵液。過濾結(jié)束后,用蒸餾水沖洗菌體3~4次,再將粘有菌體的濾紙放置于105℃下干燥至恒重,稱量。根據(jù)前后質(zhì)量差求得發(fā)酵液的生物量。
2 發(fā)酵條件優(yōu)化
2.1 初始pH對曲酸產(chǎn)量和轉(zhuǎn)化率的影響
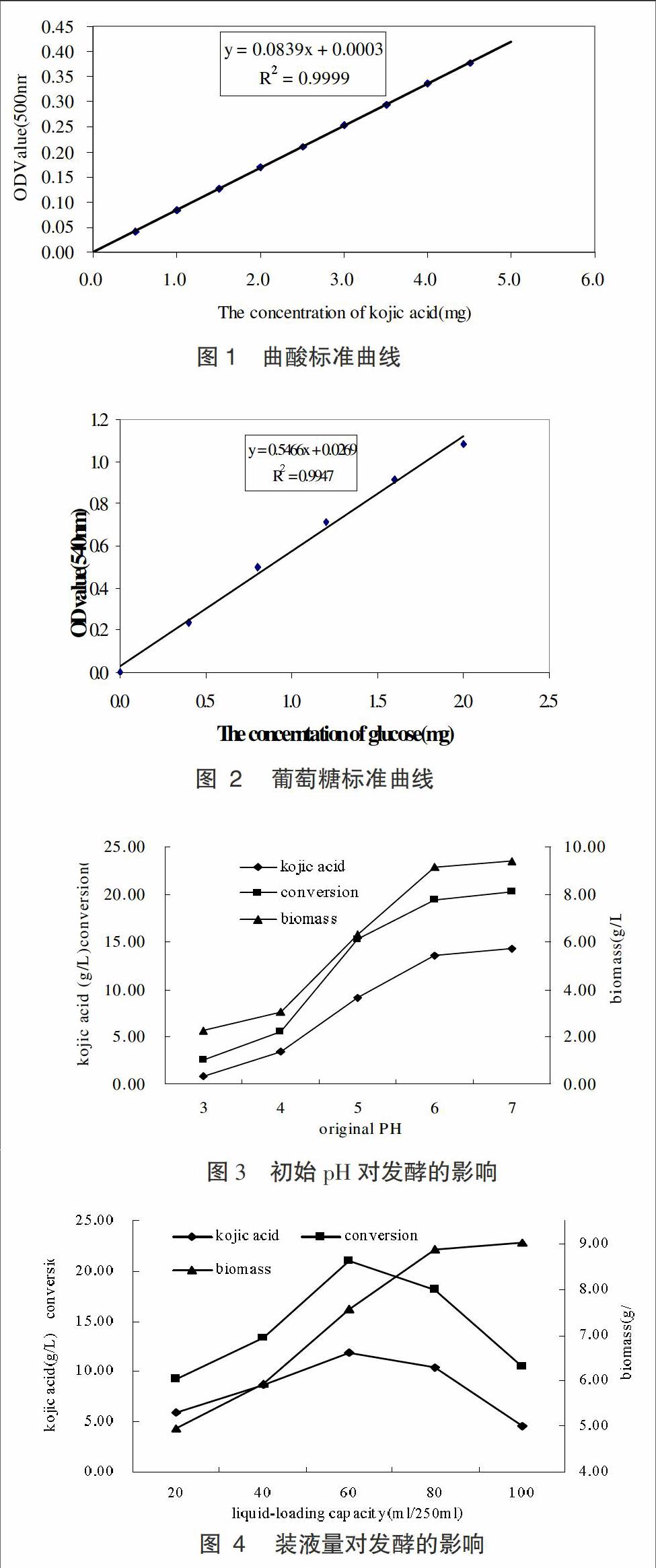
按照優(yōu)化后的培養(yǎng)基配比(見1.1.2)進行發(fā)酵試驗,調(diào)節(jié)發(fā)酵初始pH為3、4、5、6、7進行發(fā)酵試驗,研究不同初始pH對曲酸發(fā)酵的影響。
2.2 裝液量對曲酸產(chǎn)量和轉(zhuǎn)化率的影響
保持其他條件發(fā)酵條件不變,調(diào)節(jié)搖瓶裝液量為20mL/250mL、40mL/250mL、60mL/250mL、80mL/250mL、100mL/250mL,進行發(fā)酵試驗,研究不同裝液量對曲酸發(fā)酵的影響。
2.3 種齡對曲酸產(chǎn)量和轉(zhuǎn)化率的影響
保持其他條件發(fā)酵條件不變,調(diào)節(jié)用于接種的種子種齡分為6h、12h、18h、24h,接入到發(fā)酵培養(yǎng)基中進行發(fā)酵試驗,研究不同種子的種齡對發(fā)酵的影響。
2.4 接種量對曲酸產(chǎn)量和轉(zhuǎn)化率的影響
保持其他條件發(fā)酵條件不變,調(diào)節(jié)接種量為、6%、8%、10%、12%、14%,進行發(fā)酵試驗,研究不同接種量對曲酸發(fā)酵的影響。
2.5 溫度對曲酸產(chǎn)量和轉(zhuǎn)化率的影響
保持其他條件發(fā)酵條件不變,調(diào)節(jié)發(fā)酵溫度為28℃、30℃、32℃,進行發(fā)酵試驗,研究不同接種量對曲酸發(fā)酵的影響。
2.6 搖床轉(zhuǎn)速對曲酸產(chǎn)量和轉(zhuǎn)化率的影響
保持其他條件發(fā)酵條件不變,調(diào)節(jié)搖床轉(zhuǎn)速120rpm、160rpm、200rpm、240rpm,進行發(fā)酵試驗,研究不同搖床轉(zhuǎn)速對曲酸發(fā)酵的影響。
3 結(jié)果與分析
3.1 初始pH對曲酸產(chǎn)量和轉(zhuǎn)化率的影響
發(fā)酵培養(yǎng)液的pH值是微生物發(fā)酵生產(chǎn)的一個重要參數(shù),它對菌體的生長和產(chǎn)物的合成有很大的影響,微生物生長和生物合成都有其最適和能夠耐受的pH范圍,而選擇最適pH的原則是既有利于菌體的生長繁殖,又可以最大限度地獲得高的產(chǎn)量。本實驗考察了不同初始pH對曲酸合成的影響如圖3。
圖3實驗結(jié)果顯示,發(fā)酵培養(yǎng)基初始pH對整個發(fā)酵過程表現(xiàn)出比較顯著的影響。首先,初始pH在較低的情況下對菌體的生長具有顯著的抑制性,結(jié)果顯示初始pH在3~4時,生物量維持在較低水平;當初始pH值4~6時,生物量隨之急劇增加;當初始pH值6~7時,由于受到生長空間的限制,生物量維持在相對穩(wěn)定的程度。其次,菌體的產(chǎn)酸能力和曲酸轉(zhuǎn)化率也受到了初始pH的影響,較低的pH抑制了菌體生長的同時也抑制了曲酸的合成,當初始pH>4時,曲酸濃度和曲酸轉(zhuǎn)化率隨著pH值的增加同時增加,曲酸濃度最高可達14.31g/L,轉(zhuǎn)化率提高至20.31%。故將初始pH值定為7較為適宜。
3.2 裝液量對曲酸產(chǎn)量和轉(zhuǎn)化率的影響
米曲霉發(fā)酵生產(chǎn)曲酸是一種液態(tài)深層好氧發(fā)酵,底物經(jīng)微生物內(nèi)部的氧化酶經(jīng)過一系列的復雜轉(zhuǎn)化生成產(chǎn)物,因此,氧氣的供應在發(fā)酵過程中的作用就顯得尤為重要,裝液量對于菌體發(fā)酵的供氧具有重要的意義。
圖4實驗結(jié)果顯示,裝液量在20~40mL/250mL,較低的裝液量有利于提高發(fā)酵液含氧量,同時發(fā)酵液的具有較強的流動剪切力,于是形成的菌絲體結(jié)構(gòu)緊密,菌絲體的生長受到限制,具有較低的生物量,菌體的發(fā)酵能力相對較低,但是隨著裝液量的增加,生物量不斷增加,發(fā)酵液中曲酸濃度也逐漸增加,裝液量為60mL/250mL曲酸濃度和曲酸轉(zhuǎn)化率達到最高,曲酸濃度為11.86g/L,轉(zhuǎn)化率21.01%,之后隨著裝液量的增加,生物量繼續(xù)增加,超過80mL/250mL以后由于受到缺氧的限制生物量也趨于平穩(wěn)。結(jié)果表明,半缺氧的狀態(tài)下有益于曲酸發(fā)酵。裝液量為60mL/250mL較為適宜。
3.3 搖床轉(zhuǎn)速對曲酸產(chǎn)量和轉(zhuǎn)化率的影響
搖床震蕩對微生物發(fā)酵具有多方面的影響,研究搖床轉(zhuǎn)速對發(fā)酵生產(chǎn)具有十分重要的意義。從圖5中可以看出,隨著搖床轉(zhuǎn)速的不斷提高,發(fā)酵液的流動剪切力也不斷增強導致菌絲體結(jié)構(gòu)結(jié)實緊密,阻礙了。
菌絲體的生長,發(fā)酵液中的生物量隨之持續(xù)下降。搖床轉(zhuǎn)速在150~200轉(zhuǎn)/min時,發(fā)酵液中的曲酸濃度和曲酸轉(zhuǎn)化率與轉(zhuǎn)速呈正相關(guān),分別達到了14.91g/L和18.75%,當搖床轉(zhuǎn)速超過200rap以后呈穩(wěn)中有降的趨勢,可能是由于生物量持續(xù)降低的緣故所致。綜合分析,搖床轉(zhuǎn)速控制在200rap較為適宜。
3.4 種齡對曲酸產(chǎn)量和轉(zhuǎn)化率的影響
微生物生長隨著生存環(huán)境的變化會出現(xiàn)周期性的變化,用于合成次級代謝產(chǎn)的微生物多在穩(wěn)定期才發(fā)揮合成轉(zhuǎn)化功能,而在進入穩(wěn)定期之前微生物均要經(jīng)歷延滯期,在發(fā)酵工業(yè)中為了縮短延滯期,在進行微生物發(fā)酵之前首先對微生物進行種子擴大培養(yǎng),使生物量獲得迅速增殖。種子培養(yǎng)時間(種齡)過短,菌體濃度低,發(fā)酵前的延滯期長;種齡過長,菌體老化,酶活降低,生產(chǎn)效率低,因此選擇合適的種齡對后期發(fā)酵生產(chǎn)具有很大的影響,本實驗考察了不同種齡對發(fā)酵產(chǎn)酸的影響如圖6。
微生物接入到種子液中進行培養(yǎng)擴增,在適應液體深層培養(yǎng)的環(huán)境的同時,生長繁殖到一定數(shù)量和并初步具備發(fā)酵能力之后接入到發(fā)酵培養(yǎng)基中進行發(fā)酵轉(zhuǎn)化合成目的產(chǎn)物。用于發(fā)酵試驗的種子液中微生物的數(shù)量和發(fā)酵性能與種齡具有直接的關(guān)系,同時對后續(xù)的發(fā)酵試驗也具有一定的影響。
圖6實驗結(jié)果顯示,隨著種齡的增加發(fā)酵液生物量隨之增加,轉(zhuǎn)化率卻逐漸降低,當種齡超過6h后生物量增加放緩。種齡為6h是曲酸濃度達到最高,最高可達14.21g/L,轉(zhuǎn)化率為22.35%。綜合分析,用于發(fā)酵菌種的種齡應控制在6h。
3.5 接種量對曲酸產(chǎn)量和轉(zhuǎn)化率的影響
微生物在種子培養(yǎng)階段獲得充分的增殖后接入到發(fā)酵培養(yǎng)基中進行產(chǎn)物合成,接種量的多少直接影響到發(fā)酵液的生物量,因此采用較大的接種量能夠有效縮短發(fā)酵周期,但是過高的接種量往往導致發(fā)酵液中生物量迅速增加,底物消耗過快,在營養(yǎng)物質(zhì)急劇降低的同時菌體出現(xiàn)代謝放緩。
圖7試驗結(jié)果顯示,隨著接入到發(fā)酵培養(yǎng)基中種子液的增加,發(fā)酵液的生物量也隨之增加,但是當接種量超過12%以后,由于受到空間的限制,最終生物量不在增加趨于恒定;但是發(fā)酵液中曲酸濃度和曲酸轉(zhuǎn)化率隨著接種量的增加卻經(jīng)歷了先升后降的過程,接種量在6~10%曲酸濃度和曲酸轉(zhuǎn)化率隨之增加,最高分別達到了14.38g/L和21.51%。但是當接種量超過10%以后,由于微生物在消耗過多蔗糖用于合成自身結(jié)構(gòu)物質(zhì)的同時,也導致溶氧的降低,導致曲酸濃度和曲酸轉(zhuǎn)化率的降低,故以10%接種量較為適宜。
3.6 溫度對曲酸產(chǎn)量和轉(zhuǎn)化率的影響
微生物發(fā)酵主要依靠生物酶的催化作用實現(xiàn)產(chǎn)物的合成,而溫度對酶的活性具有顯著的影響,穩(wěn)定的發(fā)酵溫度是保證發(fā)酵系統(tǒng)高效運作的重要保證。不同微生物具有不同的最適生長和代謝溫度,只有在最適溫度下,才能獲得最大的發(fā)酵能力。
絲狀真菌最適生長溫度和產(chǎn)物合成溫度一般是在28℃~34℃,種屬之間存在較大的差異,即使是同一株菌的最適生長溫度和目的產(chǎn)物合成溫度也存在很大的差異。本實驗考察了不同溫度下米曲霉發(fā)酵產(chǎn)酸能力如圖8。
根據(jù)圖8實驗結(jié)果顯示,發(fā)酵溫度為30℃時獲得了較高的曲酸產(chǎn)量和較高的曲酸轉(zhuǎn)化率,故本實驗菌種最適的產(chǎn)酸溫度為30℃,但是溫度在32℃時,產(chǎn)生了更高的生物量,說明該菌株的最適生長溫度高于30℃。為了在控制生物量的同時獲得較高的曲酸產(chǎn)量,故將發(fā)酵溫度控制在30℃較為適宜。
4 結(jié)論
微生物發(fā)酵需要合適的發(fā)酵條件,發(fā)酵液pH、發(fā)酵溫度、裝液量、搖床轉(zhuǎn)速、接種量等,這些因素都將影響和制約整個發(fā)酵過程,本研究對米曲霉(Aspergillus oryzae)GIM3.423發(fā)酵蔗糖生產(chǎn)曲酸的工藝條件進行優(yōu)化,通過試驗確定了最佳的搖瓶發(fā)酵條件:發(fā)酵溫度30℃,裝液量60mL/250mL,搖床轉(zhuǎn)速200r/min,種齡6h,接種量10%,發(fā)酵周期控制在13d,在此條件下,曲酸的轉(zhuǎn)化率可達23.6%,大大提高了曲酸的發(fā)酵產(chǎn)率。
參考文獻:
[1]Jian-long Wang,Can Chen. Biosorbents for heavy metals removal and their future[J].Biotechnology Advances,2009,27(2):195 -226.
[2]H.M.Wan,C.Chen,R.Giridhar.T.S.Chang,et al.Repeated-batch production of kojic acid in a cell-retention fermenter using Aspergillus oryzae M3B9[J].Journal of Industrial Microbiology & Biotechnology,2005,32:227-233.
[3]Ana Paula D.Rodrigues,Antonio Sergio C. Carvalho.Alberdan S. Santos,et al.Kojic acid,a secondary metabolite from Aspergillus sp.,acts as an inducer of macroPHage activation[J].Cell Biology International,2011,35:335-343.
[4]Kazuhiro Takamizawa,Shinobu Nakashima,Yuuichi Yahashi,et al.Optimization of kojic acid production rate using the Box-Wilson method[J].Journal of fermentation and Bioengineering,1996,82 (4):414-416.
[5]董靜,孟憲軍.米曲霉UC6-90產(chǎn)曲酸的發(fā)酵培養(yǎng)基優(yōu)化[J].食品研究與開發(fā),2007,28(6):52-56.
[6]張理珉,程立忠,陸和生.發(fā)酵液中曲酸的提取方法比較[J].生物技術(shù),2000,10(5):44-46.
[7]王俊麗,聶國興,曹香林等.不同DNS試劑測定木糖含量的研究[J].食品研究與開發(fā),2010,31(7):1-4.
