民勤綠洲邊緣地下水埋深對檉柳灌叢生長及物種多樣性的影響
2017-05-17 13:01:56徐高興徐先英金紅喜柴成武
草原與草坪 2017年2期
徐高興,王 立,徐先英,金紅喜,柴成武
(1.甘肅農業大學 林學院,甘肅 蘭州 730070; 2.甘肅省治沙研究所/荒漠化與風沙災害防治國家重點實驗室培育基地,甘肅 蘭州 730070)
民勤綠洲邊緣地下水埋深對檉柳灌叢生長及物種多樣性的影響
徐高興1,2,王 立1,徐先英1,2,金紅喜2,柴成武2
(1.甘肅農業大學 林學院,甘肅 蘭州 730070; 2.甘肅省治沙研究所/荒漠化與風沙災害防治國家重點實驗室培育基地,甘肅 蘭州 730070)
采用空間替代時間方法,以檉柳灌叢為研究對象,研究了民勤綠洲邊緣地下水深對檉柳灌叢的生長、演變及物種多樣性的影響。結果表明:隨著地下水位的不斷下降,檉柳灌叢逐漸衰退,以白刺、紅沙為主的荒漠植物逐步發展為優勢種群。檉柳生長的最適生態水位2~3 m,生態警戒水位4 m,死亡臨界地下水位10 m。當地下水位10 m,降水和地下水共同作用導致組成檉柳灌叢的物種數最多。當地下水位下降到10 m以下,生長在固定沙丘上的檉柳衰退速度快于半固定沙丘。
地下水埋深;檉柳灌叢;生長與物種多樣性;民勤綠洲邊緣
水作為干旱、半干旱地區生態環境最活躍的因素[1],不僅是干旱區綠洲生態系統構成、發展和穩定的基礎與依據,而且決定著干旱區綠洲化與荒漠化的生態環境演化過程[2]。在地表水缺乏的干旱地區,地下水作為制約干旱荒漠綠洲區植被建設的關鍵因素,主要影響植被分布、生長、種群演替以及物種多樣性[3]。塔里木河、黑河以及石羊河向下游進行生態輸水后[4-6],河道附近及輸水區地下水位大幅度上升,植被種類和蓋度明顯增加,部分地區的沙漠化得到逆轉。荒漠植被之間的平衡關系實質是地下水與植被的平衡[7]。地下水與檉柳的關系,國內外已開展許多研究,Horton等[8]研究了地下水位下降速度對檉柳幼苗生長率和存活率的影響;王芳等[9]用高斯模型模擬檉柳出現頻率峰值的地下水位為3.7 m;Gries等[10]和Xu等[11]研究了塔克拉瑪干沙漠多枝檉柳對雨水和地下水的利用特征及響應;張麗等[12]和陳亞寧等[13]提出了塔里木河流域檉柳適宜的地下水位。但民勤綠洲邊緣地下水位變化對檉柳灌叢物種多樣性的影響,仍較少報道。
民勤綠洲位于巴丹吉林和騰格里沙漠之間,是一個“十地九沙,非灌不殖”和“地盡水耕”的灌溉綠洲[14]。檉柳(Tamarixramosissima)作為民勤綠洲邊緣主要的防風固沙樹種,生長在民勤綠洲西緣和西北緣沿大西河、小西河的綠洲-荒漠過渡帶和湖區鹽堿灘地以及“柴灣”,常形成檉柳沙包。20世紀60年代在民勤西沙窩東緣[15]、泉山區西部以及湖區北部綠洲邊緣,仍發育著一條寬一公里到數公里的檉柳沙包和白刺沙包帶,植被蓋度30%~50%。但隨著地下水位的下降和綠洲擴墾,民勤綠洲邊緣檉柳灌叢出現大范圍退化,湖區的檉柳寬度不足1 000 m,泉山區100~800 m,壩區僅有30~80 m,且呈斷續分布或“島狀”分布。民勤綠洲邊緣壩區、泉山區和湖區檉柳林均出現退化,引起退化的主要因素是地下水位下降和林地土壤水分減少[16]。因此,利用定位觀測數據并結合樣地調查,研究民勤綠洲邊緣地下水位變化對檉柳灌叢生長、演替及物種多樣性的影響,為退化檉柳灌叢恢復提供科學依據。
1 材料和方法
1.1 研究區自然概況
研究區位于石羊河下游蔡旗斷面以下的民勤綠洲邊緣,地理位置N 38°05′~39°06′,E 103°02′~104°02′,海拔1 367~1 311 m。屬典型大陸性氣候,年均氣溫7.4℃;年均降水量115.2 mm,多集中在7~9月,占全年降水量73%。年均蒸發量2 419.6 mm,為降水量的21倍;年平均風速2.35 m/s,年均沙塵暴日數25.8 d;研究區土壤分為灰棕漠土、風沙土、鹽土、草甸土和綠洲灌耕土;植被類型由人工和天然植被組成。人工植被梭梭(Haloxylonammodendron)為主的防風固沙林,面積約4.5×104hm2。綠洲邊緣的天然植被由白刺(Nitvariatangutorum)、檉柳(Tamarixramosissima)、紅沙(Reaumuriasoongorica)沙蒿(Artemisiaareanaria)等組成,其中,檉柳和白刺是主要的天然固沙植被,在綠洲壩區、泉山區和湖區均有分布。
1.2 研究方法
1.2.1 植被與地下水調查 2015年8~9月在民勤綠洲邊緣沿壩區向湖區方向選擇檉柳灌叢群落樣地10個(表1),每個樣地100 m×100 m,每個樣地采用4點法設10 m×10 m樣方4個,共40個樣方,調查植物種類、植被蓋度、個體數、高度、冠幅、新生枝長度、結實情況等指標。同時,在樣地附近選擇不同地下水位的水井,用重物下繩法測定地下水位,并取水樣測定礦化度。
數據分析采用SPSS 17.0和Excel軟件進行分析。
1.2.2β多樣性指數 用Sorensen指數和Cody指數作為β多樣性的測度方法,Sorensen指數反映群落和樣地間物種的相似性,表明群落物種構成方面的差異性;而Cody指數則反映樣方物種組成沿環境梯度的替代速率,可揭示不同環境梯度下生境被物種分割的程度或不同地段的生境多樣性,也反映生物變化速率,Cody指數越小,表明群落間越相似[17]。
Si=2c/(a+b)
Ci=(a+b-2c)/2
式中:Si和Ci分別為Sorensen指數和Cody指數;a和b為2個群落的物種數;c為2個群落的共有的物種數。
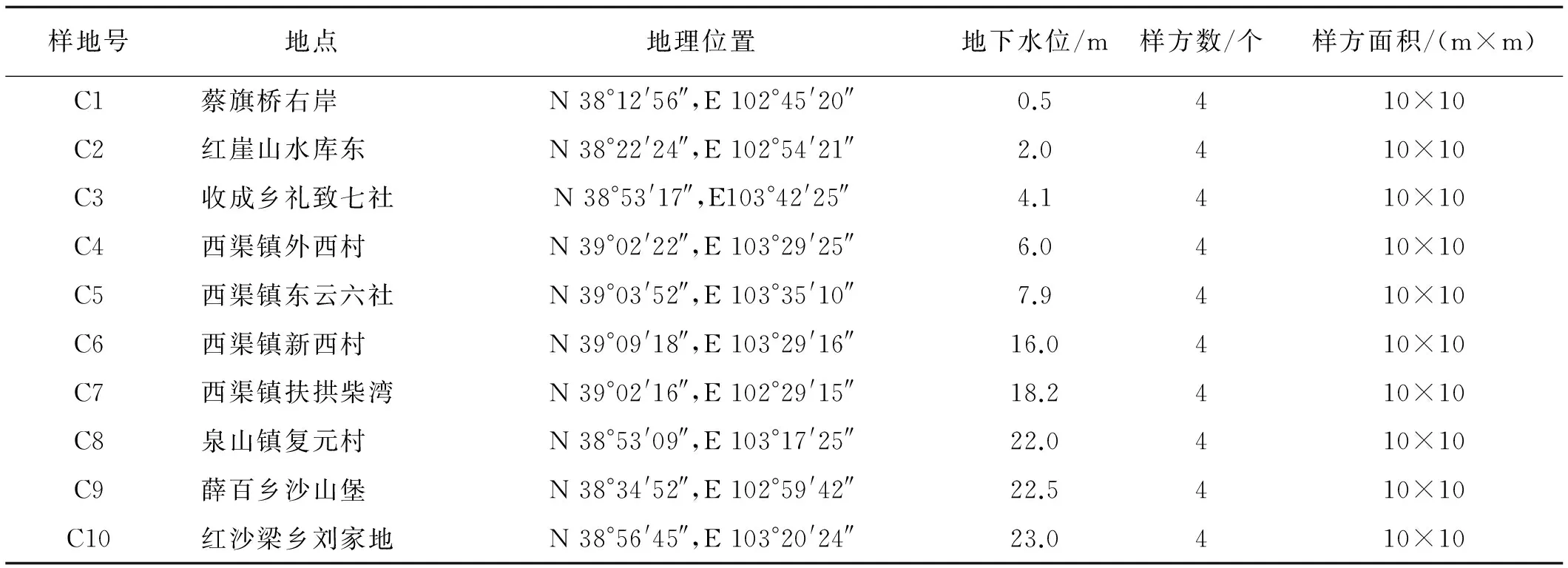
表1 民勤綠洲邊緣檉柳群落樣地狀況Table 1 Sample sties of T.ramosissima community
2 結果與分析
2.1 地下水位變化與檉柳灌叢演變
民勤綠洲邊緣檉柳種群演變取決于地表水和地下水的變化。20世紀50年代末,綠洲邊緣的檉柳固沙林依靠河流側滲和地下水生長;隨著20世紀50年代末紅崖山水庫的修建,進入大西河、小西河的地表水消失,檉柳固沙林主要依靠地下水繁衍和生長。20世紀60年代初,地下水位1~2 m,河流沿岸和鹽堿灘地檉柳、白刺和鹽爪爪開始生長;當地下水位下降到2~4 m,土壤趨于干燥和鹽漬化,檉柳、白刺在群落中成分增大,在地勢低洼地和鹽漬化較重地方,以檉柳為建群種的群落逐漸形成了檉柳+芨芨草(Achnatherumsplendens)+鹽爪爪群叢、檉柳+花花柴(Kareliniacaspia)群叢、檉柳+鹽爪爪群叢、檉柳+沙蒿(Artemisiasphaerocephala)群叢、檉柳群叢和檉柳+白刺群叢等6個群叢。到90年代中期,地下水位下降到13.9 m,以檉柳為建群種的6群叢變為檉柳和檉柳+白刺2個群叢[18];21世紀10年代中期,地下水位下降到19.5 m,檉柳群叢衰退嚴重,只有在農田邊緣生長較好的檉柳群叢。檉柳+白刺群叢和檉柳+紅紗群叢,已演變為白刺+檉柳和紅沙+檉柳群叢,檉柳逐漸被白刺和紅沙替代。
2.2 地下水位變化對檉柳灌叢物種組成的影響
當C1樣地的地下水位為0.5 m,檉柳群落組成僅有3個種,種數較少。當C8,C9和C10樣地地下水位下降到20 m以下時,群落組成植物種數也較少,只有6種;當C5和C6樣地平均地下水位在10 m,檉柳群落的物種數可達8~9種。檉柳灌叢群落植物種數隨地下水位的變化符合二次多項式分布,R2=0.8437(圖1)。在植物群落中,優勢種群對群落結構和環境的形成有明顯的控制作用[19]。高地下水位,檉柳自身密度大、蓋度高,同時土壤鹽堿重,檉柳的競爭排斥作用,導致檉柳群落中只有一些耐鹽的鹽生植物和靠降水生長的草本植物,如蘆葦、鹽生草(Halogetonglomeratus)、白刺、五星蒿(Bassiadasyphylla)等;當地下水位下降到4 m以下,檉柳開始逐漸退化,種群優勢逐漸下降。在水分脅迫下,根系通過吸收利用土壤水維持生長,檉柳灌叢沙堆仍處于固定、半固定狀態,一些鹽生植被逐漸退出,其他物種的種子在沙面居留,在適宜的降水條件下生長繁衍,種類數量增加;當地下水位下降到12m,直至23m,深層的土壤水分被進一步消耗,出現極度干旱脅迫,群落中的檉柳種群衰退嚴重,固定、半固定檉柳沙堆呈半流動狀態,甚至演變為流動沙丘,鹽生物種消失,其他物種一方面由于檉柳對降水的競爭限制,一些物種也退出檉柳群落,另一方面可能流動的沙面,不利于種子定居而無法生存,導致物種數降低。
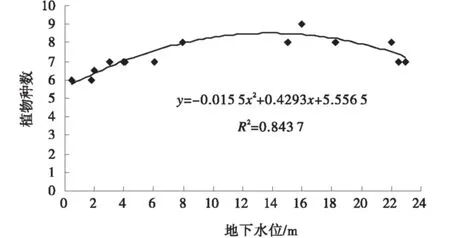
圖1 地下水位與檉柳群落植物種數關系Fig.1 Relationship between groundwater table and vegetation species of T.ramosissima community
不同地下水位檉柳群落物種組成表明(表2),檉柳群落植物組成結構簡單,物種數較少,共有10科、20屬,21種,主要以草本植物為主,共13種,占群落植物種的61.9%,其中,一年生草本9種,占草本種的69.2%。植物種主要以藜科植物為主,占33.3%,其次蒺藜科,占14.3%。當地下水位較高時,檉柳群落植物種組成以濕生和鹽生植物為主,隨著地下水位不斷下降,鹽生和濕生植物逐漸退出,沙生和旱生植物逐漸成為檉柳群落的組成種類。
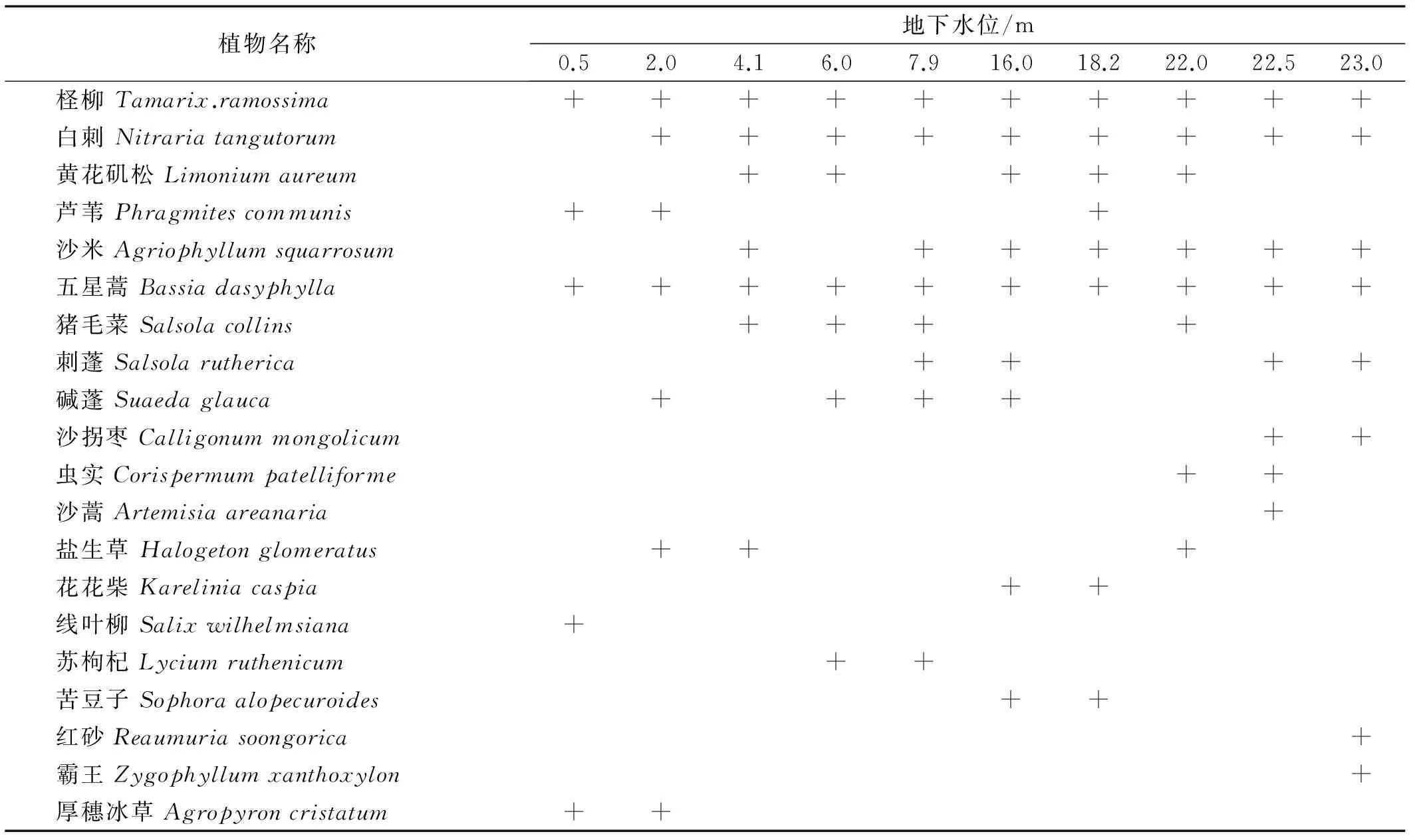
表2 不同地下水位檉柳群落植物種類組成Table 2 The species composition of T.ramosissima community at different groundwater levels
注:+++代表該物種存在
2.3 地下水位對檉柳灌叢β多樣性指數影響
β多樣性可直觀反映生境之間物種組成的差異性。控制β多樣性的生態因子有水分、土壤、地貌及干擾等[20]。在土壤、地貌以及干擾相似的民勤綠洲邊緣,不同地下水位檉柳群落的β多樣性表明(表3),地下水位相近樣方間的Sorensen多樣性指數均大于地下水位差距大的值,說明地下水環境梯度的變化,引起各樣地植物種類間的變化。Sorensen值越小,表明各樣地間共有種類較少,相似程度也越低,植物生境之間的異質性差異大。樣方C3對樣方C4、C5、C6之間變化小,說明地下水位差異不大,群落結構相似性較好。樣方C6和C7之間Sorensen指數最大,反映其相似程度最高,這2個樣地地下水位分別為16 m和18.2 m。樣方C3~C6、C8~C10變化不大,其之間群落結構相似性較好。
樣方C6和C7的Cody最小,表明這2個樣方間相似程度最高。樣方C4和樣方C9、C10之間的Cody最大,表明樣方間的相似程度最低(表4);相鄰樣方在地下水位2~4 m,6~8 m和18~22 m之間出現波動,說明檉柳群落植被組成從一種類型向另一種類型轉變,由于地下水位、土壤鹽分、降水以及沙面的變化所帶來的干擾形成,表明影響群落物種多樣性的因素是多方面的。
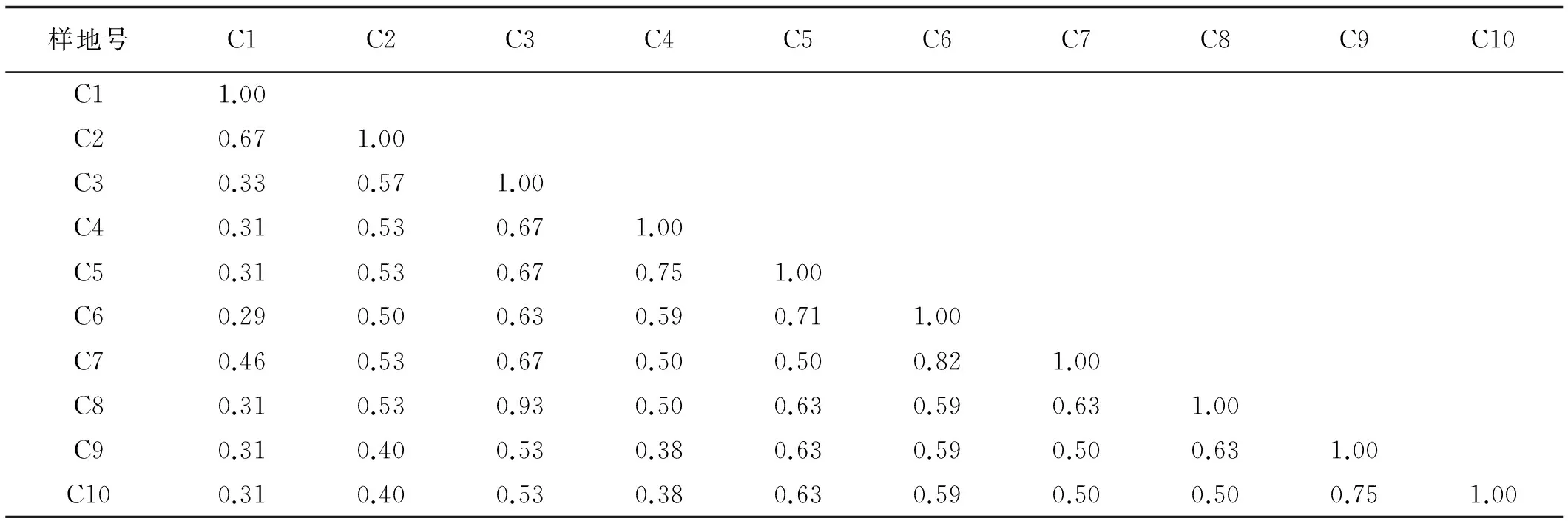
表3 不同地下水位檉柳群落生境間物種Sorensen指數Table 3 The Sorensen indexes of T.ramosissima community at the gradients of groundwater level
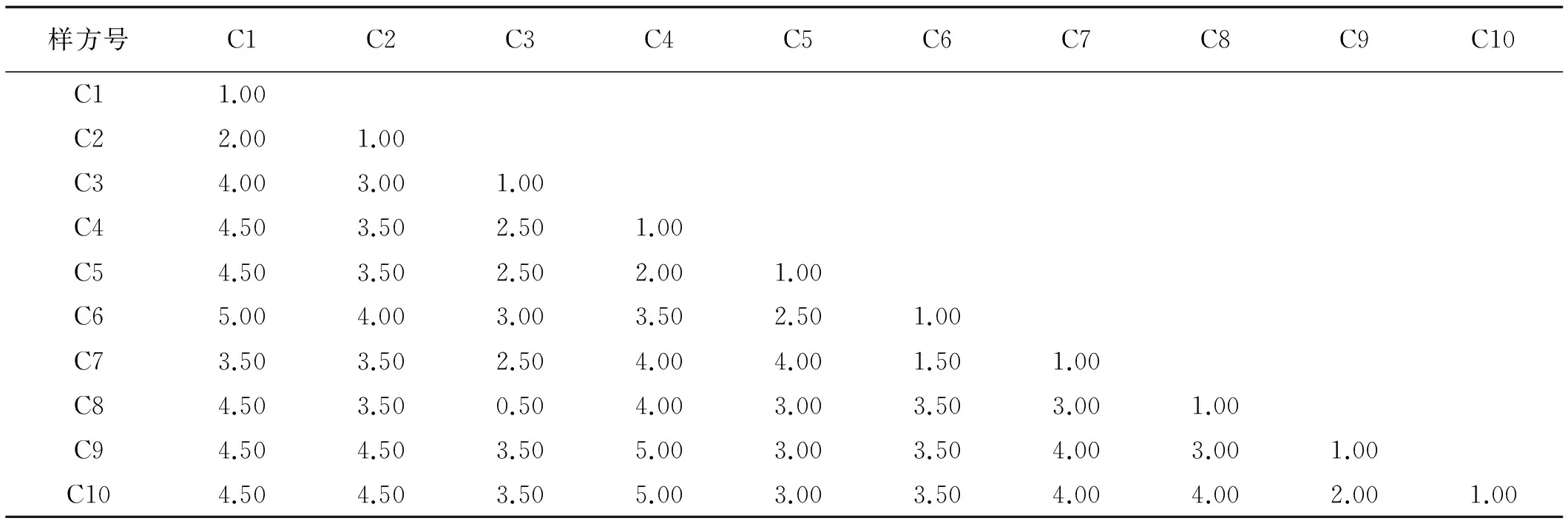
表4 不同地下水位檉柳群落生境間物種Cody指數Table 4 The Cody indexes of T.ramosissima community at the gradients of groundwater level
2.4 檉柳灌叢生長與地下水位關系
檉柳灌叢生長狀況與地下水位的關系表明(圖2),隨著地下水位下降,檉柳灌叢的高度生長和群落蓋度呈現降低,當地下水位下降到4 m以下,檉柳枝條的枯死率隨地下水位下降逐漸增大。當C1樣地地下水位在0.5 m時,種群密度高,雖然植被蓋度80%,但由于植物根系長期浸泡在水中,影響根系呼吸,從而抑制其高度;當紅崖山水庫東側地下水位在2 m時,檉柳新稍平均生長量達77.0 cm,形成的灌叢沙堆體積大,種群生長良好。當樣地C3地下水位達到4.1 m,盡管植被蓋度接近30%,但檉柳枝條枯死率為5.4%。當C3~C5地下水在4~8 m,固定沙丘上生長的檉柳逐漸衰退,生長高度降低,平均枝條枯死率13.1%;當C6和C7樣地地下水位在16~18 m時,檉柳枝條枯死率達49.4%;C8,C9和C10樣地地下水位超過20 m,固定沙丘上生長的檉柳枯死率在77.2%~85.0%,檉柳灌叢沙堆出現極度衰退狀態。
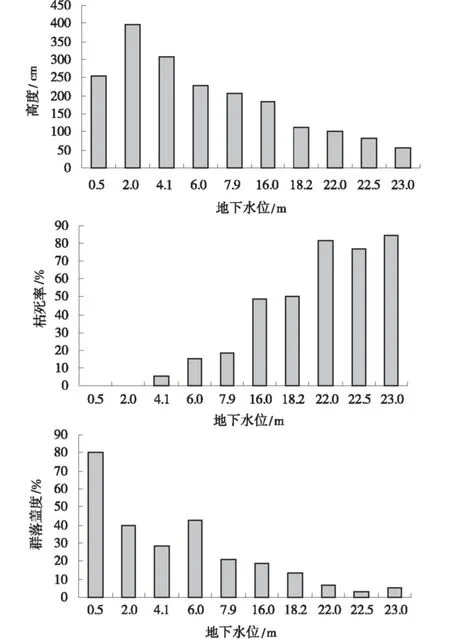
圖2 綠洲邊緣檉柳生長狀況與地下水Fig.2 Effect of groundwater level on growth of T.ramosissima
1984~1992年民勤綠洲邊緣沙井子連續固定樣地觀測表明(圖3),檉柳在植物群落中蓋度占總蓋度比率隨著地下水位的下降總趨勢在降低[21]。但觀測初期,地下水位7.5 m,已超出檉柳4 m的生態警戒水位,檉柳開始利用土壤水分和降水維持生長。當地下水位在7.5~10.9 m,檉柳蓋度占群落總蓋度比率下降不顯著,表明土壤水分和降水共同維持檉柳生長。8.0 m和8.2 m地下水位年份,降水量分別比平均年份增加25.7%和29.8%,檉柳蓋度占群落總蓋度的比率有所增加,說明檉柳的生長和衰退是地下水與降水共同作用的結果,主要受地下水的影響。當地下水位下降到10 m以下,檉柳蓋度在群落中的占比下降很快,表明檉柳達到最大生態地下水位。地下水位的下降對白刺群落中的檉柳生長發育有明顯的抑制作用,隨著地下水位的下降,退化明顯,但個別年份較多的降水也能使退化過程有所改善[21]。
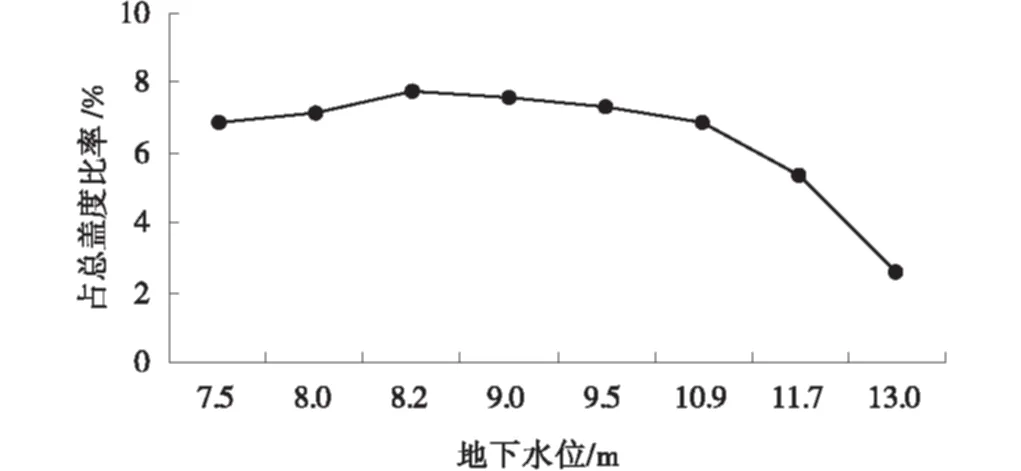
圖3 檉柳在植物群落中蓋度占比對地下水位的響應Fig.3 Coverage ratio of T.ramosissima response to groundwater table changes in vegetation community
3 討論
民勤綠洲邊緣檉柳灌叢的發育與石羊河下游的大西河、小西河以及青土湖地表水與地下水密切相關。當人類活動造成石羊河下游流入民勤綠洲的河流變成季節性河流或完全斷流干涸后,荒漠區植被的演替主要取決于地下水的變化。隨著地下水位的下降,荒漠植物群落從濕生系列過度到旱生系列,植被的旱生化程度逐漸加強[22],群落結構趨向于簡單[23]。民勤綠洲邊緣生態系統演替序列中,檉柳群落由多途徑發育而成,而且其最終演替方向應當是多元的。民勤綠洲邊緣檉柳群落演替表現出的相似性也是地下水位不斷下降的結果,當地下水位1~2 m,河流沿岸和鹽堿灘地檉柳開始發育;當地下水位下降到2~4 m時,檉柳有6個群叢;地下水位下降到10 m以下,民勤綠洲邊緣以檉柳為建群種群叢的只有2個,而且以檉柳為建群種逐漸被白刺和紅沙替代。張武文等[24]、彭鴻嘉等[25]、劉加珍等[26]的研究也得出類似的結論。
研究認為,當地下水埋深較小時,組成檉柳群落植物種數量少,主要由于大量潛水蒸發使地表鹽分聚集或根系呼吸不暢,使非耐鹽植物難以生長;隨著地下水埋深的增加,植物個體數和蓋度增加,主要是潛水蒸發減弱,植物生境的改善逐漸有利于植物生存。這與宋冬梅等[27]研究報道一致。當地下水位下降到一定深度,如民勤綠洲邊緣地下水位在10 m,降水和地下水共同作用導致檉柳群落物種數增加,組成檉柳群落的物種數最多;由于檉柳根系存在水力提升現象[28-29],植物根系通過水力提升來保持淺層土壤相對濕潤,維持淺根系植物生長的水分條件,從而維持生物多樣性的群落結構[30-31]。隨著地下水位進一步下降,地下水埋深超出檉柳生長的最大生態水位,檉柳群落生長僅依靠降水維持生長,導致組成檉柳群落的物種多樣性降低,檉柳衰退加速。
據徐海量等[32]研究,塔里木河流域地下水位在3.5 m 以上時,土壤含水率隨地下水位的下降而明顯下降;地下水位在4.0 m以下時,這種變化趨勢減弱,地下水位對表層土壤水影響區間在3.5~4.0 m。當地下水位超過3.5 m,地下水可以通過大氣蒸發和毛管作用影響表層土壤濕度;當地下水超過4 m時,水位的變化對土壤含水率的影響基本消失。檉柳出現頻率峰值所對應的地下水位在2.29 m[12]。新疆塔里木河流域檉柳生長的適宜地下水位為2~4.5 m[33]。考慮石羊河下游民勤綠洲與塔里木河流域綠洲的相似性,確定民勤綠洲邊緣檉柳的最適地下水位2~3 m,警戒水位4 m。陳亞寧等[13]報道,塔里木河檉柳死亡的臨界地下水位為9 m;馮起等[34]認為,黑河下游的額濟納地區,地下水埋深小于5 m,檉柳生長旺盛,5~7 m可維持生長,7~8 m以上生長嚴重衰退、大部分植株死亡。但據楊維康等研究[35],在塔里木河古老沖積平原以及沙漠中常與風沙土呈復區分布的丘間低地,龜裂土生境下地下水在8~10 m或更低時,常有稀疏的檉柳沙包分布,而且檉柳灌木層高1~2 m,群落蓋度在10%,而民勤綠洲邊緣多龜裂地分布,考慮到塔里木河下游多年平均降水量在42 mm以下[36]、黑河下游38.2 mm[37],而民勤綠洲邊緣降水113.5 mm,確定民勤綠洲邊緣檉柳死亡的臨界地下水位為10 m。
周洪華等[38]報道了塔里木河下游綠洲-荒漠過渡帶植物群落的β多樣性,表明從綠洲經過過渡帶到荒漠,地下水埋深對生境變化起著重要作用,與此次研究結論相似,從而進一步證明了群落多樣性隨地下水位的變化發生相應變化。
檉柳衰退的指標應以枝條枯死率為主,在沙丘上,當檉柳枝條枯死率達20%檉柳灌叢處于輕度退化,枯死率50%,中度退化;枯枝率達到80%時,灌叢處于嚴重退化[39]。研究中,當地下水位在4~8 m,固定沙包檉柳生長處于輕度退化狀態,與趙雪等[39]、張元禧[40]研究結論基本一致;當地下水位在16~18 m時,檉柳枯死率49.4%,處于中度退化狀態,與張元禧[40]研究當地下水位>10 m時,檉柳全部植株死亡不一致,可能與檉柳生長立地類型有關。根據調查發現,半固定沙丘甚至半流動沙丘的檉柳灌叢隨地下水位的下降,退化程度和速度慢于固定沙丘。由于檉柳為耐沙埋植物,沙埋后易產生大量不定根。當地下水位下降超過檉柳根系時,固定沙丘沒有適度的流沙埋壓枝條,不定根無法形成,其衰退速度加快。而半固定沙丘由于存在適度沙埋產生不定根,對降水的利用率高,其衰退速度比固定檉柳沙包慢。
4 結論
(1)民勤綠洲邊緣檉柳群落的發育取決于石羊河下游的地下水位變化。當地下水位下降到1~2 m時,檉柳群落開始發育。隨著地下水位的不斷下降,檉柳群落逐漸將向以白刺、紅沙為主的荒漠群落發展。
(2)民勤綠洲邊緣檉柳的最適地下水位2~3 m,警戒水位4 m,死亡臨界地下水位10 m。
(3)當地下水位在10 m,降水和地下水共同作用導致民勤綠洲邊緣檉柳群落物種多樣性最大。
(4)在民勤綠洲邊緣,當地下水位超過生態警戒水位后,隨著地下水位的進一步下降,檉柳開始衰退,但不同立地條件,其衰退程度存在差異。在相同地下水位條件下,半固定沙丘生長的檉柳衰退速度慢于固定沙丘生長的。
[1] 蘇德榮.干旱地區草坪的水分管理[J].草原與草坪,2000(4):26-29.
[2] 陳亞寧,李衛紅,徐海量,等.塔里木河下游地下水位對植被的影響[J].地理學報,2003,58(4):542-549.
[3] 馬玉蕾,王德,劉俊民,等.地下水與植被關系的研究進展[J].水資源與水工程學報,2013,24(5):36-44.
[4] 徐海量,陳亞寧,雷加強.塔里木河下游生態輸水對沙漠化逆轉的影響[J].中國沙漠,2004,24(2):173-176.
[5] 郭巧玲,馮起,任韶雯,等.黑河生態輸水對流域植被的影響研究[J].中國農村水利水電,2009(3):36-43.
[6] 董志玲,徐先英,金紅喜,等.生態輸水對石羊河尾閭湖區植被的影響[J].干旱區資源與環境,2015,29(7):101-106.
[7] 馮起,程國棟,譚志剛.荒漠綠洲植被生長與生態地下水位的研究[J].中國沙漠,1998,18(增刊):106-109.
[8] Horton J L,Kolb T E,Hart S C.Responses of riparian trees to inter-annual variation in ground water depth in a semi-arid river basin[J].Plant,Cell and Environment,2001,24:293-304.
[9] 王芳,梁瑞駒,楊小柳,等.中國西北地區生態需水研究Ⅰ.干旱半干旱地區生態需水理論分析[J].自然資源學報,2002,17(1):1-8.
[10] Gries D,Zeng F,Foetzki A.etal.Growth and water relations of Tamarix ramosissima and Populus euphratica on Taklamakan desert dunes in relation to depth to a permanent water table[J].Plant,Cell and Environment,2003,26:725-736.
[11] Xu H,Li Y.Water-use strategy of three central Asian desert shrubs and their responses to rain pulse events[J].Plant and Soil,2006,285:5-17.
[12] 張麗,董增川,黃曉玲.干旱區典型植物生長與地下水位關系的模型研究[J].中國沙漠,2004,24(1):110-113.
[13] 陳亞寧,王強,李衛紅,等.植物生理生態學數據表征的合理地下水位研究-以塔里木河下游生態恢復過程為例[J].科學通報,2006,51(增1):7-13.
[14] 李宗禮.民勤綠洲水資源承載能力初步研究[C]∥王繼和.中國西北荒漠區持續農業與沙漠綜合治理國際學術交流會論文集.蘭州:蘭州大學出版社,1998:188-196.
[15] 李并成.殘存在民勤西沙窩中的古代遺址[J].中國沙漠,1990,10(2):35-42.
[16] 馬全林,王繼和,劉虎俊,等.民勤綠洲邊緣檉柳荒漠林的時空變化及其驅動因素[J].中國沙漠,2006,26(9):802-808.
[17] Patricia K,Kevin J G,Jack J L.Measuring beta diversity for presence-absence data[J].Journal of Animal Ecology,2003,72:367-382.
[18] 楊自輝.民勤荒漠植被的初步研究[J].甘肅林業科技,1995,3:26-28.
[19] 董鳴,王義風,孔繁志,等.陸地生物群落調查觀測與分析[M].北京:中國標準出版社,1996:21-22.
[20] Loreau M.Are communities saturated on the relationship between a,b and g diversity[J].Ecology Letter,2000(3):73-76.
[21] 楊自輝,高志海.荒漠綠洲邊緣降水和地下水對白刺群落消長的影響[J].應用生態學報,2000,11(6):923-926.
[22] 趙文智,劉鵠.荒漠區植被對地下水埋深響應研究進展[J].生態學報,2006,26(8):2702-2708.
[23] 張宏峰,陳亞寧,陳亞鵬,等.塔里木河下游植物群落的物種數量變化與生態系統動態研究[J].生態學雜志,2004,23(4):21-24.
[24] 張武文,馬秀珍,譚志剛.額濟納平原植被分布與地下水關系的研究[J].干旱區資源與環境,2000,14(5):31-35.
[25] 彭鴻嘉,傅伯杰,陳利頂,等.甘肅民勤荒漠區植被演替特征及驅動力研究[J].中國沙漠,2004,24(5);628-633.
[26] 劉加珍,陳亞寧,李衛紅,等.塔里木河下游植被群落分布與衰退演替趨勢分析[J].生態學報,2004,24(2):379-383.
[27] 宋冬梅,肖篤寧,馬明國,等.民勤湖區地下水資源時空變化對灌區景觀生態安全的影響研究[J].應用生態學報,2004,15(10):1815-1820.
[28] 袁國富,張佩,雪沙沙,等.沙丘多枝檉柳灌叢根系土壤含水量變化特征與根系水力提升證據[J].植物生態學報,2012,36(10):1033-1042.
[29] 付愛紅,陳亞寧,李衛紅.中國黑河下游荒漠河岸林植物群落水分利用策略研究[J].中國科學(地球科學),2014,44(4):693-705.
[30] Dawson T E.Hydraulic lift and water use by plants:implications for water balance,performance and plant-plant interactions[J].Oecologia,1993,95:565-574.
[31] Moreira M Z,Scholz F G,Bucci S J,etal.Hydraulic lift in a neotropical savanna[J].Functional Ecology,2003,17:573-581.
[32] 徐海量,宋郁東,王強,等.塔里木河中下游地區不同地下水位對植被的影響[J].植物生態學報,2004,25(3):400-405.
[33] 宋郁東,樊自立,雷志棟,等.塔里木河水資源與生態研究[M].烏魯木齊:新疆人民出版社,2000:249-264.
[34] 馮起,程國棟,譚志剛.荒漠綠洲植被生長與生態地下水位的研究[J].中國沙漠,1998,18(增刊1):106-109.
[35] 楊維康,張道遠,張立運,等.新疆主要檉柳屬植物的生態類型劃分與生境相似性研究[J].2004,27(2):186-192.
[36] 張宏峰,李衛紅,葛洪濤,等.塔里木河下游地下水位與水化學成分關聯度排序分析[J].干旱區地理,2003,26(3):260-263.
[37] 肖生春,肖紅浪,周茅先,等.近百年來西居延海湖泊水位變化的湖岸林樹輪記錄[J].冰川凍土,2004,26(5):557-562.
[38] 周洪華,陳亞寧,李衛紅.塔里木河下游綠洲-荒漠過渡帶植物多樣性特征及其優勢種群分布格局[J].中國沙漠,2009,29(4):688-696.
[39] 趙雪,張小由,李啟森,等.額濟納綠洲沙漠化對檉柳群落影響的研究[J].中國沙漠,2004,24(4):467-472.
[40] 張元禧.河西走廊地下水開采的環境影響及其對策[M]∥地下水開發利用研究與實踐.北京:中國農業科技出版社,1999.
Impact of groundwater level on growth and species diversity ofTamarixramosissimacommunity around oasis fringe in Minqin
XU Gao-xin1,2,WANG Li1,XU Xian-ying1,2,JIN Hong-xi2,CHAI Cheng-wu2
(1.ForestryCollegeofGansuAgricultureUniversity,Lanzhou730070,China;2.GansuDesertControlResearchInstitute,StateKeyLaboratoryBreedingBaseofDesertificationandAeolianSandDisasterCombating,Lanzhou730070,China)
The relationship between groundwater level changes andTamarixramosissimacommunity was studied in order to offer the scientific evidences for the recover of degradedT.ramosissimacommunity in the oasis fringe in Minqin.The results showed thatT.ramosissimacommunity succeeded gradually toNitrariatangutorumandReaumuriasoongoricacommunities along with the decrease of groundwater level.The depths of 2 to 3 m,4 m and 10 m of groundwater level were the optimum,warning and death critical levels individually.The species number was the highest while the groundwater depth was around 10 m.The degraded speed of theT.ramosissimacommunity on fixed sand dune was faster than that on semi-fixed sand dune when the groundwater level dropped less than 10 m.
groundwater depth;Tamarixramosissimacommunity;growth and species diversity;oasis fringe,Minqin
2016-11-14;
2016-12-05
國家科技支撐計劃(2012BAD16B0203);國家重點基礎研究發展計劃(2012CB723203)資助
徐高興(1993-),女,甘肅武威人,碩士研究生。 E-mail:2910824960@qq.com
S 793.5;P641.2
A
1009-5500(2017)02-0049-08
王立為通訊作者。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
汽車觀察(2018年10期)2018-11-06 07:05:26
