水稻矮化劍葉卷曲突變體dcfl1的鑒定與基因精細定位
2017-06-05 14:16:19張孝波謝佳張曉瓊田維江何沛龍劉思岑何光華鐘秉強桑賢春
中國農業科學 2017年9期
關鍵詞:水稻
張孝波,謝佳,張曉瓊,田維江,何沛龍,劉思岑,何光華,鐘秉強,桑賢春
(西南大學水稻研究所/轉基因植物與安全控制重慶市重點實驗室,重慶 400715)
水稻矮化劍葉卷曲突變體dcfl1的鑒定與基因精細定位
張孝波,謝佳,張曉瓊,田維江,何沛龍,劉思岑,何光華,鐘秉強,桑賢春
(西南大學水稻研究所/轉基因植物與安全控制重慶市重點實驗室,重慶 400715)
【目的】對一個水稻矮化劍葉卷曲突變體進行鑒定與基因定位,為水稻等禾谷類作物劍葉形態發育及分子改良奠定基礎。【方法】在秈型水稻恢復系縉恢10號的甲基磺酸乙酯(EMS)突變庫中篩選到一個隱性矮化劍葉卷曲突變體,命名為dcfl1(dwarf and curled flag leaf 1)。田間小區種植,全生育期內觀察dcfl1和野生型的株型變化。苗期利用掃描電鏡觀察葉鞘內表皮細胞大小;孕穗期和抽穗期利用石蠟切片觀察劍葉基部形態;開花期測定劍葉、倒2葉和倒3葉的葉綠素含量;成熟期考查株高、有效穗數、穗實粒數、結實率和千粒重等主要農藝性狀。配制西農1A/dcfl1雜交組合,利用F1和F2群體進行遺傳分析,并利用F2隱性群體進行基因定位。【結果】生育期內,突變體dcfl1都表現出矮化性狀。dcfl1葉鞘內表皮細胞長度明顯比野生型要短,達到了極顯著水平。與野生型相比,穗長、倒1節間和倒2節間均顯著變短,倒3節間和倒4節間無顯著變化。抽穗期dcfl1劍葉的葉片和葉鞘連接處硬化,劍葉基部展開受阻,半邊葉片向內卷曲,劍葉上部和中部正常,其他葉片也正常。農藝性狀調查發現,dcfl1的有效穗數為14.24,極顯著高于野生型的11.62,穗粒數、實粒數、結實率和千粒重等則無顯著變化。此外,dcfl1的葉色略深,劍葉、倒2葉和倒3葉的葉綠素a含量均極顯著高于野生型,類胡蘿卜素含量也略有升高,但僅劍葉達到極顯著差異水平,葉綠素b的含量則無顯著變化。西農1A/dcfl1的F1群體中,株高和劍葉表型與野生型一致。F2群體中分離出正常和突變兩種表型,突變表型與dcfl1類似,植株株高變矮,劍葉基部特異卷曲,說明矮化和劍葉基部特異卷曲是一對共分離性狀。且兩種表型分離比符合3﹕1,表明dcfl1突變型受1對隱性核基因控制。利用620株F2隱性單株,最終將DCFL1精細定位在第3染色體短臂InDel標記Ind03-11和Ind03-6之間78 kb的物理范圍內,包含15個注釋基因,為DCFL1的克隆和水稻劍葉形態發育機理研究奠定了基礎。【結論】dcfl1是一個水稻矮化劍葉基部特異卷曲突變體,基因精細定位在第3染色體78 kb的物理范圍內。
水稻;矮化;劍葉卷曲;遺傳分析;精細定位
0 引言
【研究意義】葉片形態、大小和顏色直接決定了群體光能利用效率,進而影響作物的產量和品質,受到育種和分子生物學家的廣泛關注[1]。劍葉是水稻最重要的功能葉,葉片大小和形態與單株產量、單穗重、穗粒數均呈極顯著正相關,同時,劍葉葉型也是水稻理想株型的重要構成部分[2-3]。因此,闡明劍葉發育的分子機理對水稻生產具有重要意義。【前人研究進展】目前,利用突變體已在水稻中克隆了一系列調控葉片大小和形態發育的基因,如窄葉基因 NAL1[4]、NAL2/NAL3[5]、NAL7[6],窄卷葉基因NRL1[7-8],卷葉基因 CFL1[9]、SRL1[10]和 RL14[11]等,為闡釋水稻葉片發育的分子機理奠定了基礎。然而,這些突變體多表現為階段性或全生育期所有葉片發育缺陷,劍葉特異異常發育的突變體還鮮有報道。目前普遍認為劍葉發育屬于復雜性狀,已利用 F2分離群體、雙單倍體(DH)、重組自交系(RILs)和單片段代換系(CSSLs)等材料,在水稻12條染色體上定位了系列控制劍葉大小的QTL。YAN等[12]利用IR64/Azucena衍生DH群體在第 4染色體上定位到一個控制劍葉大小的QTL。KOBAYASHI等[13]利用 Milyang23/Akihikari組合衍生的RIL群體(191個家系)定位到7個控制劍葉長度和5個控制劍葉寬度的QTL。近來,BIAN等[14]利用CSSLs群體鑒定了4個控制水稻劍葉寬度、1個控制劍葉面積和 2個控制劍葉角度的 QTL;ZHANG等[3]則利用 RIL群體在海南和杭州兩地種植,檢測到9個控制劍葉長度和14個控制劍葉寬度的QTL,其中僅7個為先前鑒定的QTL;CHEN等[15]利用 RIL群體也鑒定到 5個控制劍葉葉寬發育的QTL,并認為主效QTL qFLW4是NAL1的同義突變,其通過選擇性剪切調控劍葉寬度的發育。【本研究切入點】盡管目前已經鑒定到大量控制劍葉發育的QTL,但這些研究主要集中在葉片大小上,對劍葉形態發育還鮮有報道。利用EMS誘變秈型水稻恢復系縉恢10號,從其后代鑒定到一個矮化,劍葉基部特異卷曲突變體,暫命名為dcfl1(dwarf and curled flag leaf 1)。【擬解決的關鍵問題】本研究對突變體dcfl1進行了形態鑒定、細胞學觀察和基因精細定位等研究,為DCFL1的克隆和功能研究奠定了基礎,有利于水稻劍葉形態發育的分子機理的闡釋。此外,劍葉基部卷曲導致葉片直立,有利于群體的通透性,dcfl1在水稻育種中也具有重要的應用價值,是一類新型種質資源。
1 材料與方法
1.1 試驗材料
突變體 dcfl1來自西南大學水稻所培育的晚秈恢復系縉恢10號EMS誘變庫,經過多世代連續種植,突變性狀已穩定遺傳。配置西農1A/ dcfl1雜交組合,調查西農1A、dcfl1及其雜交組合F1和F2的株高及劍葉形態,進行遺傳分析,利用F2群體隱性單株進行基因定位。西農1A是西南大學水稻所選育的不育系,整個生育期株高及葉片均正常。
1.2 主要農藝性狀鑒定
田間小區種植dcfl1和野生型縉恢10號,成熟期分別測量10株材料的株高和節間長并調查穗長、有效穗數、穗粒數、穗實粒數、結實率和千粒重等主要農藝性狀。
1.3 光合色素含量測定
開花期,參照文獻[16]描述的方法測定dcfl1和野生型的光合色素含量,測定部位為劍葉、倒2葉和倒3葉的葉片中部。
1.4 石蠟切片分析
孕穗期和抽穗期分別選取未全展的劍葉基部和全展劍葉基部,根據文獻[17]略有改動。用FAA固定液固定后,依次進行乙醇脫水、二甲苯透明、石蠟包埋、切片、番紅固綠染色等步驟。并在體視鏡下觀察葉片形態。
1.5 掃描電鏡觀察
田間種植三葉期秧苗,移至掃描電鏡室,利用日立 SU350型掃描電鏡在-20℃冷凍條件下觀察第二片葉的葉鞘內表皮細胞。
1.6 DNA的提取
在西農1A/ dcfl1的F2群體中分別選取正常和突變單株各10株,構成正常基因池和突變基因池。采取改良的CTAB法[18]提取基因池DNA,采用堿煮法[19]提取定位群體單株基因組DNA。
1.7 SSR分析
RM系列SSR引物來源于http://www.gramene.org/網站。根據縉恢10號和西農1A的DNA序列差異,利用Vector 10軟件設計InDel標記。SSR引物及InDel標記均由上海英俊生物公司合成。PCR反應總體積12.5 μL,包括1.25 μL的10×PCR緩沖液、0.65 μL的25 mmol·L-1MgC12、0.5 μL 2.5 mmol·L-1dNTPs、8.0 μL的ddH2O、1.0 μL的10 μmol·L-1引物、1.0 μL的模板DNA和0.1 μL的5 U·μL-1Taq酶。PCR反應程序為94℃ 5 min;94℃ 20 s,55℃ 20 s,72℃ 20 s,35個循環; 72℃ 10 min。PCR產物用10%的非變性聚丙烯酰胺凝膠電泳后,0.1%AgNO3染色10 min,去離子水漂洗2次,1%質量濃度的氫氧化鈉和0.1%甲醛混合液顯色,觀察照相。
1.8 遺傳圖譜構建
西農 1A/dcfl1雜交組合的 F2群體中,具有西農1A帶型的單株記為A,具有dcfl1帶型的單株記為B,具有 F1雜合體帶型的單株記為 H。利用公式[(H+2A)/2n]×100%計算遺傳重組率,其中,H代表定位群體中雜合體帶型單株的數量,A代表正常株帶型單株的數量,n表示F2群體隱性單株總株數。
2 結果
2.1 dcfl1的形態鑒定
突變體dcfl1整個生育期都表現出矮化性狀。播種后5 d的dcfl1籽苗地上部長度只有野生型的一半,二葉期也明顯矮于野生型。dcfl1的倒1節間、倒2節間和穗長顯著短于野生型,進而導致株高半矮化(圖1)。掃描電鏡觀察野生型和 dcfl1葉鞘內表皮細胞,發現dcfl1的細胞長度明顯比野生型短,達到了極顯著水平,而細胞寬度并無顯著差異(圖 2),暗示突變體的矮化性狀可能是由于細胞長度變短而導致的。農藝性狀分析(表1)發現,dcfl1的有效穗數為14.24,極顯著高于野生型的11.62,穗粒數、實粒數、結實率和千粒重等則無顯著變化。此外,dcfl1的葉片呈深綠色(圖1-D),光合色素測定表明劍葉、倒2葉和倒3葉的葉綠素a含量均極顯著增加,葉綠素b含量無顯著變化,進而導致dcfl1的葉綠素a/b值極顯著增加;dcfl1的類胡蘿卜素含量雖略有升高,但僅劍葉中的含量極顯著高于野生型。
抽穗期dcfl1劍葉的葉片和葉鞘連接處硬化,葉基部展開受阻,半邊葉片向內卷曲(圖3-A),而劍葉上部和中部無卷曲現象(圖3-B)。為進一步觀察劍葉基部形態,將未展開的劍葉和全展開時期的劍葉基部進行石蠟切片。發現在劍葉發育前期,野生型的劍葉有規律地向內卷曲排列(圖3-C),而dcfl1的內卷葉排列不規則(圖3-D)。劍葉全展開后,野生型的劍葉基部呈向外展開狀(圖3-E),而dcfl1的劍葉基部展開受阻,一半葉片呈向外展開狀,但內卷葉始終無法向外展開(圖3-F),從而形成了劍葉基部半邊葉片向內卷曲這一突變表型。盡管目前已經報道了許多矮化突變體,但劍葉基部特異卷曲的水稻矮化突變體還沒有報道。
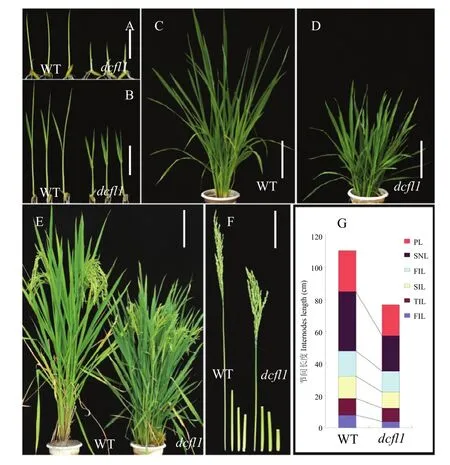
圖1 突變體dcfl1和野生型(WT)縉恢10號形態鑒定Fig. 1 Morphology identification of the dcfl1 and the wild type

表1 突變體dcfl1與野生型的農藝性狀Table 1 Agronomic traits of dcfl1 mutant and the wild type
2.2 遺傳分析
西農 1A/dcfl1雜交組合的 F1代植株劍葉基部均無卷曲現象,且株高正常。F2群體中分離出正常和突變2種類型,正常型1 940株,突變型620株,突變型植株也表現為劍葉基部卷曲和矮化現象,與dcfl1類似,表明劍葉基部卷曲和矮化是1對共分離性狀。χ2測驗顯示正常植株與突變植株的分離比符合 3﹕1(χ2=0.79<χ20.05=3.84),暗示dcfl1的突變性狀受1對隱性核基因控制。

圖2 突變體dcfl1和野生型(WT)縉恢10號細胞大小比較Fig. 2 Comparison of cell length and width between the dcfl1 and the wild type
2.3 基因定位
利用平均分布在水稻12條染色體上、在西農1A和野生型縉恢10號間呈多態性的96對SSR標記篩選正常基因池和突變基因池,發現第3染色體上的SSR標記RM6297、RM14347、RM5474和RM5955在基因池間呈現多態性,暗示可能與 DCFL1連鎖。利用140個 F2隱性單株進行驗證,確定了連鎖關系并將DCFL1初步限定在RM14347和RM5474之間,遺傳距離分別為4.32和3.81 cM。
為進一步確定DCFL1的物理位置,在DCFL1初步定位區間內設計開發了20對InDel標記,多態性篩選發現 4對在親本之間具有多態性,分別命名為Ind03-8、Ind03-11、Ind03-6和Ind03-4(表2)。利用這些多態性標記對620株西農1A/dcfl1雜交組合的F2隱性單株進行分析,結果表明,Ind03-8、Ind03-11、Ind03-6和Ind03-4的交換株分別為9、2、3和7個,且前 2個標記的交換株不同于后者,從而將 DCFL1精細定位在InDel標記Ind03-11和Ind03-6之間,物理距離約為78 kb(圖4)。

圖3 突變體dcfl1和野生型(WT)縉恢10號的劍葉形態Fig. 3 Flag leaf phenotype of the dcfl1 and the wild type
根據Gramene(http://ensembl.gramene.org/ Oryza_ sativa/Info/Index)和 Rice genome annotation project(http://rice.plantbiology.msu.edu/)提供的信息,發現在DCFL1精細定位區間內,包含15個注釋基因(表3),5個編碼逆轉錄轉座子蛋白,6個編碼表達蛋白,4個為功能基因,分別編碼細胞色素P450蛋白、60S核糖體蛋白L21-2、β-淀粉酶和RNA基序識別蛋白。

表2 基因定位InDel引物Table 2 Primers of InDel markers for gene mapping
3 討論

圖4 DCFL1在水稻第3染色體上的分子定位Fig. 4 Molecular mapping of DCFL1 gene on rice chromosome 3
在水稻生產中,葉片適度卷曲有利于葉的挺直,從而改善了群體結構、提高了光能利用率,在水稻高產育種中具有重要的應用價值。因此,揭示水稻葉片卷曲的遺傳機制不僅有利于葉片發育的分子機理闡釋,也為水稻株型育種提供了基礎材料和理論支撐。目前,在水稻中至少報道了20多份卷葉突變體,這些突變體的形成多受植物葉片極性建成、泡狀細胞大小和數量變化以及環境因素的影響。蔥狀卷曲突變體sll1[20]是由于葉片遠軸面厚壁組織細胞發育異常而表現出卷曲。位于第3染色體上的突變體srl2[21]表現出葉片半卷,葉片變窄,株高降低。類似于 sll1,srl2也是由于遠軸面厚壁組織細胞異常導致葉片卷曲。突變體adl1[22]表現為下表皮泡狀細胞異位發育,從而導致葉片向遠軸面卷曲。另一個外卷突變體oul1[23]來源于水稻最外層細胞特異基因 Roc5的敲除,表現出近軸側泡狀細胞體積變大而引起葉片外卷。SRL1編碼糖基磷脂酰肌醇固定蛋白,調控葉片上表皮泡狀細胞數量從而控制葉片卷曲[10]。突變體rl14[11]卷葉表型是由于近軸面泡狀細胞萎縮引起,RL14通過調控次生細胞壁組分合成從而影響葉片水分運輸,并進一步影響泡狀細胞形態。環境誘導型卷葉突變體rl15(t)葉片卷曲行為受環境誘導,濕度是誘導突變體卷曲的主要因素[25]。值得注意的是突變體cfl1[9],由于劍葉卷曲,所以該突變體被命名為 curly flag leaf1,但其他葉片也受到影響,因此,cfl1也屬于所有葉片全卷突變體。此外,rl12(t)葉片卷曲特性隨著發育進程而發生變化,卷曲表型主要發生在葉片中上部1/3處,中下部正常,劍葉亦是如此[24]。本文報道的 dcfl1突變體,僅劍葉基部卷曲,明顯不同于已報道的水稻卷葉突變體,也不同于劍葉發育缺陷相關突變體,因此,dcfl1是一類新型劍葉基部特異卷曲突變體。

表3 定位區間內注釋基因Table 3 Annotated genes of the gene mapping range
目前報道的葉片發育調控基因,多具有“一因多效”性。如,Ghd7調控抽穗期和產量性狀,同時也影響劍葉的葉面積[26-27];dtl1是一個矮化突變體,同時也表現葉片卷曲、分蘗減少和不育等性狀[28]。dcf1并不是傳統的葉卷曲突變體,只是劍葉基部特異卷曲,劍葉中上部和其他葉片均正常,因此它的育性并沒有受影響。另外,盡管dcfl1的穗長變短了,但dcfl1的穗型更緊湊,使dcfl1穗粒數無明顯變化,而且有效穗增加了,從而暗示該突變體的產量有所提高。突變體dcfl1除劍葉特異卷曲外,還表現植株的矮化和葉片顏色深綠。這可能是由于 DCFL1的多效性造成的,引起這些突變表型的原因還有待進一步研究。
利用SSR等分子標記最終將DCFL1定位在第3染色體InDel標記Ind03-11和Ind03-6之間78 kb物理距離內,包含15個注釋基因,其中4個為功能基因。細胞色素 P450編碼基因已克隆,可能通過脂類代謝途徑調控細胞伸長,其突變體oscyp96b4主要表現為植株半矮化和育性降低。它的矮化表型并不受激素的調控,而是通過轉錄劑量的方式來降低水稻株高[29]。ZHANG等[30]鑒定到一個 oscyp96b4的等位突變體sd37,除全生育期植株矮化外,還表現稻穗和花軸變短,籽粒變小。另一個等位突變體dss1萌發以及早期生長均延遲,它的矮化表型也不受外源激素影響。但內源ABA的積累和GA的缺陷可能是dss1矮化的原因,而且其耐旱性增強[31]。此外,WANG等[32]鑒定了oscyp96b4第三個等位突變體bsh1,其表現出株高、千粒重以及每株產量均顯著降低,葉鞘角質層蠟質含量降低,暗示BSH1可能參與蠟質生物合成。其他3個基因則沒有克隆:β-淀粉酶是一個非生物脅迫蛋白,響應磷和鉀的缺失[33-34];60S核糖體蛋白L21-2編碼基因在不育系和保持系之間具有表達差異[35];RNA基序識別蛋白則尚沒有描述。從表型和定位結果推測,DCFL1可能是一個調控劍葉發育的新基因。
4 結論
EMS誘變獲得一個矮化和劍葉基部卷曲的新型水稻突變體dcfl1,其表現為分蘗數增多、葉色深綠。產量性狀除有效穗極顯著升高外,其他無明顯變化。dcfl1的葉綠素a含量極顯著高于野生型,導致葉綠素a/b比值極顯著增加。劍葉基部卷曲和植株矮化受同1對隱性核基因調控,利用西農1A和dcfl1雜交組合的F2分離群體,最終將DCFL1定位在水稻第3染色體InDel標記Ind03-11和Ind03-6之間78 kb的物理距離內,包含15個注釋基因。
[1] 徐靜, 王莉, 錢前, 張光恒. 水稻葉片形態建成分子調控機制研究進展. 作物學報, 2013, 39(5): 767-774.
XU J, WANG L, QIAN Q, ZHANG G H. Research advance in molecule regulation mechanism of leaf morphogenesis in rice (Oryza sativa L.). Acta Agronomica Sinica, 2013, 39(5): 767-774. (in Chinese)
[2] YUE B, XUE W Y, LUO L J, XING Y Z. QTL analysis for flag leaf characteristics and their relationships with yield and yield traits in rice. Acta Genetica Sinica, 2006, 33(9): 824-832.
[3] ZHANG B, YE W J, REN D Y, TIAN P, PENG Y L, GAO Y, RUAN B P, WANG L, ZHANG G H, GUO L B, QIAN Q, GAO Z Y. Genetic analysis of flag leaf size and candidate genes determination of a major QTL for flag leaf width in rice. Rice, 2015, 8: 2.
[4] QI J, QIAN Q, BU Q Y, LI S Y, CHEN Q, SUN J Q, LIANG W X, ZHOU Y H, CHU C C, LI X G, REN F G, PALME K, ZHAO B R, CHEN J F, CHEN M S, LI C Y. Mutation of rice Narrow leaf1 gene, which encodes a novel protein, affects vein patterning and polar auxin transport. Plant Physiology, 2008, 147(4): 1947-1959.
[5] CHO S H, YOO S C, ZHANG H, PANDEYA D, KOH H J, HWANG J Y, KIM G T, PAEK N C. The rice narrow leaf2 and narrow leaf3 loci encode WUSCHEL related homeobox 3A (OsWOX3A) and function in leaf, spikelet, tiller and lateral root development. New Phytologist, 2013, 198(4): 1071-1084.
[6] FUJINO K, MATSUDA Y, OZAWA K, NISHIMURA T, KOSHIBA T, W. FRAAIJE M, SEKIGUCHI H. NARROW LEAF 7 controls leaf shape mediated by auxin in rice. Molecular Genetic Genome, 2008, 279(5): 499-507.
[7] HU J, ZHU L, ZENG D, GAO Z, GUO L, FANG Y, ZHANG G, DONG G, YAN M, LIU J, QIAN Q. Identification and characterization of NARROW AND ROLLED LEAF 1, a novel gene regulating leaf morphology and plant architecture in rice. Plant Molecular Biology, 2010, 73(3): 283-292.
[8] WU C, FU Y P, HU G C, SI H M, CHENG S H, LIU W Z. Isolation and characterization of a rice mutant with narrow and rolled leaves. Planta, 2010, 232(2): 313-324.
[9] WU R H, LI S B, HE S, WA?MANN F, YU C H, QIN G J, SCHREIBER L, QU L J, GU H Y. CFL1, a WW domain protein, regulates cuticle development by modulating the function of HDG1, a class IV homeodomain transcription factor, in rice and Arabidopsis. The Plant Cell, 2011, 23(9): 3392-3411.
[10] XIANG J J, ZHANG G H, QIAN Q, XUE H W. Semi-rolled leaf1 encodes a putative glycosylphosphatidylinositol-anchored protein and modulates rice leaf rolling by regulating the formation of bulliform cells. Plant Physiology, 2012, 159(4): 1488-1500.
[11] FANG L K, ZHAO F M, CONG Y F, SANG X C, DU Q, WANG D Z, LI Y F, LING Y H, YANG Z L, HE G H. Rolling-Leaf14 is a 2OG-Fe (II) oxygenase family protein of unknown function that modulates rice leaf rolling by affecting secondary cell wall formation in leaves. Plant Biotechnology Journal, 2012, 10(5): 524-532.
[12] YAN J P, ZHU J, HE C X, BENMOUSSA M, WU P. Molecular marker-assisted dissection of genotype× environment interaction for plant type traits in rice (Oryza sativa L.). Crop Science, 1999, 39(2): 538-544.
[13] KOBAYASHI S, FUKUTA Y, MORITA S, SATO T, OSAKI M, KHUSH G S. Quantitative trait loci affecting flag leaf development in rice (Oryza sativa L.). Breed Science, 2003, 53(3): 255-262.
[14] BIAN J M, HE H H, SHI H, ZHU G Q, LI C J, ZHU C L, PENG X S, YU Q Y, FU J R, HE X P, CHEN X R, HU L F, OUYANG L J. Quantitative trait loci mapping for flag leaf traits in rice using a chromosome segment substitution line population. Plant Breeding, 2014, 133(2): 203-209.
[15] CHEN M L, LUO J, SHAO G N, WEI X J, TANG S Q, SHENG Z H, SONG J, HU P S. Fine mapping of a major QTL for flag leaf width in rice, qFLW4, which might be caused by alternative splicing of NAL1. Plant Cell Reports, 2012, 31(5): 863-872.
[16] LICHTENTHALER H K. Hlorophylls and carotenoids pigments of photosynthetic biomembranes. Methods in Enzymology, 1987, 48: 350-382.
[17] SANG X C, LI Y F, LUO Z K, WANG N, LING Y H, ZHAO H M, YANG Z L, LUO H F, LIU Y S, HE G H. CHIMERIC FLORAL ORGANS 1, encoding a monocot-specific MADS box protein, regulates floral organ identity in rice. Plant Physiology, 2012, 160(2): 788-807.
[18] MURRAY M G, THOMPSON W F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Research, 1980, 8(19): 4321-4325.
[19] 桑賢春, 何光華, 張毅, 楊正林, 裴炎. 水稻PCR擴增模板的快速制備. 遺傳, 2003, 25(6): 705-707.
SANG X C, HE G H, ZHANG Y, YANG Z L, PEI Y. The simple gain of templates of rice genomes DNA for PCR. Hereditas, 2003, 25(6): 705-707. (in Chinese)
[20] ZHANG G H, XU Q, ZHU X D, QIAN Q, XUE H W. SHALLOT-LIKE1 is a KANADI transcription factor that modulates rice leaf rolling by regulating leaf abaxial cell development. The Plant Cell, 2009, 21(3): 719-735.
[21] LIU X F, LI M, LIU K, TANG D, SUN M F, LI Y F, SHEN Y, DU G J, CHENG Z K. Semi-Rolled Leaf 2 modulates rice leaf rolling by regulating abaxial side cell differentiation. Journal of Experimental Botany, 2016, 67(8): 2139-2150.
[22] HIBARA K, OBARA M, HAYASHIDA E, ABE M, ISHIMARU T, SATOH H, ITOH J, NAGATO Y. The ADAXIALIZED LEAF 1 gene functions in leaf and embryonic pattern formation in rice. Developmental Biology, 2009, 334(2): 345-354.
[23] ZOU L P, SUN X H, ZHANG Z G, LIU P, WU J X, TIAN C J, QIU J L, LU T G. Leaf rolling controlled by the homeodomain leucine zipper class IV gene Roc5 in rice. Plant Physiology, 2011, 156(3): 1589-1602.
[24] 羅遠章, 趙芳明, 桑賢春, 凌英華, 楊正林, 何光華. 水稻新型卷葉突變體 rl12(t)的遺傳分析和基因定位. 作物學報, 2009, 35(11): 1967-1972.
LUO Y Z, ZHAO F M, SANG X C, LING Y H, YANG Z L, HE G H. Genetic analysis and gene mapping of a novel rolled leaf mutant rl12(t) in rice. Acta Agronomica Sinica, 2009, 35(11): 1967-1972. (in Chinese)
[25] 張禮霞, 劉合芹, 于新, 王林友, 范宏環, 金慶生, 王建軍. 水稻卷葉突變體 rl15(t)的生理學分析及基因定位. 中國農業科學, 2014, 47(14): 2881-2888.
ZHANG L X, LIU H Q, YU X, WANG L Y, FAN H H, JIN Q S, WANG J J. Molecular mapping and physiological characterization of a novel mutant rl15(t) in rice. Scientia Agricultura Sinica, 2014, 47(14): 2881-2888. (in Chinese)
[26] XUE W Y, XING Y Z, WENG X Y, ZHAO Y, TANG W J, WANG L, ZHOU H J, YU S B, XU C G, LI X H, ZHANG Q F. Natural variation in Ghd7 is an important regulator of heading date and yield potential in rice. Nature Genetics, 2008, 40(6): 761-767.
[27] 談聰, 翁小煜, 鄢文豪, 白旭峰, 邢永忠. 多效性基因Ghd7調控水稻劍葉面積. 遺傳, 2012, 34(7): 901-906.
TAN C, WENG X Y, YAN W H, BAI X F, XING Y Z. Ghd7, a pleiotropic gene controlling flag leaf area in rice. Hereditas, 2012, 34(7): 901-906. (in Chinese)
[28] 張帆濤, 方軍, 孫昌輝, 李潤寶, 羅向東, 謝建坤, 鄧曉建, 儲成才. 水稻矮稈突變體dtl1的分離鑒定及其突變基因的精細定位. 遺傳, 2012, 34(1): 79-86.
ZHANG F T, FANG J, SUN C H, LI R B, LUO X D, XIE J K, DENG X J, CHU C C. Characterization of a rice dwarf and twist leaf 1 (dtl1) mutant and fine mapping of DTL1 gene. Hereditas, 2012, 34(1): 79-86. (in Chinese)
[29] RAMAMOORTHY R, JIANG S Y, RAMACHANDRAN S. Oryza sativa cytochrome P450 family member OsCYP96B4 reduces plant height in a transcript dosage dependent manner. PLoS ONE, 2011, 6(11): e28069.
[30] ZHANG J, LIU X Q, LI S Y, CHENG Z K, LI C Y. The rice semi-dwarf mutant sd37, caused by a mutation in CYP96B4, plays an important role in the fine-tuning of plant growth. PLoS ONE, 2014, 9(2): e88068.
[31] TAMIRU M, UNDAN J R, TAKAGI H, ABE A, YOSHIDA K, UNDAN J Q, NATSUME S, UEMURA A, SAITOH H, MATSUMURA H, URASAKI N, YOKOTA T, TERAUCHI R. A cytochrome P450, OsDSS1, is involved in growth and drought sress responses in rice (Oryza sativa L.). Plant Moecular Biology, 2015, 88(1): 85-99.
[32] WANG X L, CHENG Z J, ZHAO Z C, GAN L, QIN R Z, ZHOU K N, MA W W, ZHANG B C, WANG J L, ZHAI H Q, WAN J M. BRITTLE SHEATH1 encoding OsCYP96B4 is involed in secondary cell wall formation in rice. Plant Cell Reports, 2016, 35: 745-755.
[33] PARK M R, BAEK S H, DE LOS REYES B G, YUN S J, HASENSTEIN K H. Transcriptome profiling characterizes phosphate deficiency effects on carbohydrate metabolism in rice leaves. Journal of Plant Physiology, 2012, 169(2): 193-205.
[34] SHANKAR A, SINGH A, KANWAR P, SRIVASTAVA A K, PANDEY A, SUPRASANNA P, KAPOOR S, PANDEY G K. Gene expression analysis of rice seedling under potassium deprivation reveals major changes in metabolism and signaling components. PLoS ONE, 2013, 8(7): e70321.
[35] YANA J J, TIAN H, WANG S Z, SHAO J Z, ZHENG Y Z, ZHANG H Y, GUO L, DING Y. Pollen developmental defects in ZD-CMS rice line explored by cytological, molecular and proteomic approaches. Journal of Proteomics, 2014, 108: 110-123.
(責任編輯 李莉)
Identification and Gene Mapping of a Dwarf and Curled Flag Leaf Mutant dcfl1 in Rice (Oryza sativa L.)
ZHANG XiaoBo, XIE Jia, ZHANG XiaoQiong, TIAN WeiJiang, HE PeiLong, LIU SiCen, HE GuangHua, ZHONG BingQiang, SANG XianChun
(Rice Research Institute of Southwest University/Chongqing Key Laboratory of Application and Safety Control of Genetically Modified Crops, Chongqing 400715)
【Objective】Leaf blade is an important factor of plant type, which is directly related to leaf photosynthetic area and light energy utilization. Flag leaf is most prominently in the formation of rice production. Study of the genes which regulate flag leaf development in rice is of very significance in rice functional genomics research and molecular breeding. A novel flag leaf mutant has been identified and the results of study will provide a foundation for the research of leaf morphological formation and plant type breeding in Oryza sativa L.【Method】A dwarf and curled flag leaf mutant (dcfl1) was discovered from the progeny of indica restorerline Jinhui10 with seeds treated by ethyl methane sulfonate (EMS) and the traits of dwarf and curled flag leaf base inherited steadily after multi-generations’ self-fertility. The second leaf sheath was observed by scanning electron microscopy (SEM) at the three-leaf stage. The flag leaf base was used for paraffin section at the booting and heading stages. At the blooming stage, the characteristics of chloroplast pigment of the flag, second and third leaf blades were measured. At the maturity stage, agronomic traits such as plant height, panicle length, efficient panicle per plant, seed number per panicle, filled grain number per panicle, seed setting ratio, and 1000-seed weight were measured. The dcfl1 was crossed with indica sterile line Xinong 1A, and the F1and F2generations were used for genetic analysis. Additionally, gene mapping was performed based on the recessive individuals of the F2generation of Xinong 1A/ dcfl1.【Result】The dcfl1 was dwarf in all phases of plant development. The cell length of the 2nd leaf sheath surface of the dcfl1 was significantly shorter than the wild type. The panicle length, the first and the second internode of the dcfl1 were all significantly shorter than those of the wild type. The dcfl1 displayed a severe curl at the base of flag leaf blade after the heading stage, while the upper of flag leaf blade was nearly normal in the flag leaf. Meanwhile, the other leaf blades appeared as normal as the wild type. No significant differences were detected in grain number per panicle, filled grain number per panicle, seed setting rate and 1000-seed weight between the dcfl1 and the wild type. However, the number of the tiller in the dcfl1 was more than the wild type and the efficient panicle per plant was increased significantly than the wild type. Having the dark green leaves, the contents of chlorophyll a and total chlorophyll in the dcfl1 increased significantly compared with those of the wild type for the flag leaves, the second and the third leaves. Genetic analysis indicated that the dwarf and curled flag leaf traits of dcfl1 were controlled by a recessive nuclear gene. Based on the F2population derived from a cross between the dcfl1 and an indica sterile line, Xinong 1A, the gene was fine mapped on chromosome 3 between InDel marker Ind03-11 and Ind03-6 with the physical distance 78 kb, containing fifteen annotated genes.【Conclusion】The dcfl1 is a novel recessive dwarf and curled flag leaf mutant coming from EMS-inducement. The DCFL1 was mapped on chromosome 3 with 78 kb physical distance. These results will provide a foundation for map-based cloning of DCFL1 gene and understanding of the molecular mechanism of the rice flag leaf.
rice (Oryza sativ L.); dwarf; curled flag leaf; genetic analysis; gene mapping
2016-11-09;接受日期:2017-01-04
中央高校基本科研業務費項目(XDJK2013A023)、重慶市研究生科研創新項目(CYS14043)
聯系方式:張孝波,E-mail:zhangxiaobo@163.com。通信作者桑賢春,E-mail:sangxianchun@163.com
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
