結(jié)縷草肉桂醇脫氫酶基因家族全基因組序列鑒定和表達(dá)分析
2017-06-19 17:07:28史經(jīng)昂張兵肖曉琳馬晶晶楊向陽劉建秀
草業(yè)學(xué)報 2017年6期
關(guān)鍵詞:分析
史經(jīng)昂,張兵,肖曉琳,馬晶晶,楊向陽,劉建秀
(江蘇省中國科學(xué)院植物研究所,江蘇 南京 210014)
?
結(jié)縷草肉桂醇脫氫酶基因家族全基因組序列鑒定和表達(dá)分析
史經(jīng)昂,張兵*,肖曉琳,馬晶晶,楊向陽,劉建秀
(江蘇省中國科學(xué)院植物研究所,江蘇 南京 210014)
肉桂醇脫氫酶(cinnamyl alcohol dehydrogenase,CAD,EC 1.1.1.195)是植物木質(zhì)素合成途徑的關(guān)鍵酶,通過消耗NADPH,將松柏醛等復(fù)雜醛轉(zhuǎn)化成相應(yīng)的醇單體,在植物生長發(fā)育過程和逆境應(yīng)答中起到重要作用。本研究從結(jié)縷草基因組中鑒定獲得了16個肉桂醇脫氫酶(ZjCAD)基因,對其進(jìn)行了系統(tǒng)進(jìn)化與基因結(jié)構(gòu)的生物信息學(xué)分析。半定量PCR分析顯示5個ZjCAD基因在葉片中特異表達(dá),木質(zhì)素染色結(jié)合不同種源材料ZjCAD基因的表達(dá)分析進(jìn)一步表明ZjCAD6的表達(dá)與葉片木質(zhì)素含量正相關(guān),提示ZjCAD6可能為控制結(jié)縷草葉片木質(zhì)素合成的關(guān)鍵基因。ZjCAD6基因全長2736 bp,有3個內(nèi)含子和4個外顯子,開放閱讀框?yàn)?074 bp,編碼357個氨基酸。ZjCAD6蛋白含有兩個Zn2+結(jié)合位點(diǎn)與一個NADP(H)結(jié)合結(jié)構(gòu)域,與已報道的其他植物具催化活性的CAD蛋白特征一致。上述結(jié)果為進(jìn)一步研究結(jié)縷草葉片木質(zhì)素含量及其機(jī)械強(qiáng)度等性狀形成的分子機(jī)理提供了重要的參考。
結(jié)縷草;木質(zhì)素;肉桂醇脫氫酶;生物信息學(xué)分析;基因表達(dá)
結(jié)縷草(Zoysiajaponica)是一種優(yōu)良的多年生暖季型草坪草,具有發(fā)達(dá)的根狀莖和匍匐莖,被廣泛應(yīng)用于庭院草坪、運(yùn)動場草坪、保土草坪等各種草坪的建植中。在幾種暖季型草坪草中,結(jié)縷草具有優(yōu)良的耐踐踏能力,葉片近革質(zhì),其木質(zhì)素含量顯著高于雜交狗牙根(Cynodondactylonvar.dactylon×C.transvaalensiscv.)和海雀稗(Paspalumvaginatum)[1]。
木質(zhì)素是高等植物細(xì)胞壁的重要組成成分,主要存在于次生細(xì)胞壁中,可以提高細(xì)胞壁的強(qiáng)度,為植物組織提供機(jī)械支撐。木質(zhì)素還和植物長距離的運(yùn)輸營養(yǎng)和水分、抵御病蟲害等生理功能密切相關(guān)。在被子植物中,木質(zhì)素主要由松柏醇衍化的愈創(chuàng)木基(G)型單體、芥子醇衍化的紫丁香基(S)單體以及對香豆醇衍化的對羥基苯基(H)單體構(gòu)成,3種單體通過β-O-4醚鍵、C-C鍵、聯(lián)苯醚鍵聚合成復(fù)雜的高分子聚合物[2-4]。
苯丙氨酸途徑是植物木質(zhì)素生物合成的主要途徑,這個過程中涉及多種酶:苯丙氨酸解氨酶(phenylalanine ammonia-lyase,PAL)[5]、肉桂酸-4-羥基化酶(cinnamic acid 4-hydroxylase,C4H)[6]、4-香豆酸輔酶A連接酶(4-coumarate:coenzyme A ligase,4CL)[7]、3-羥基肉桂酰轉(zhuǎn)移酶(p-coumarate 3-hydroxylase,C3H)[8]、咖啡酸-O-甲基轉(zhuǎn)移酶(caffeic acid O-methyltransferase,COMT)[9]、阿魏酸-5-羥基化酶(ferulate-5-hydroxylase,F5H)[10]、肉桂醇脫氫酶(cinnamyl alcohol dehydrogenase,CAD)[11-12]等。CAD是木質(zhì)素生物合成途徑的關(guān)鍵限速酶之一,在單體合成反應(yīng)的最后一步將不同醛單體催化成相應(yīng)的醇單體[13]。CAD基因以多基因家族形式存在植物體中,自第一個CAD基因從煙草(Nicotianatabacum)莖段中獲得以來[14],多種植物的CAD基因均被廣泛研究。在擬南芥(Arabidopsisthaliana)中發(fā)現(xiàn)的9個CAD基因中,CAD4、CAD5被發(fā)現(xiàn)是控制木質(zhì)素生物合成的主要基因[15-16],AtCAD1則與擬南芥莖延伸過程中的木質(zhì)化相關(guān)[17]。水稻(Oryzasativa) 12個CAD基因中,OsCAD2在木質(zhì)素合成途徑中起最主要的作用,表達(dá)譜分析發(fā)現(xiàn)其在節(jié)間、葉鞘、根中高表達(dá)[14]。玉米(Zeamays) bm1突變體中木質(zhì)素含量和結(jié)構(gòu)都發(fā)生了改變,并使葉脈與莖的厚壁組織變成紅褐色,這一突變最終被定位在CAD基因上[18]。CAD-RNAi導(dǎo)致玉米植株總木質(zhì)素減少,但并未影響植株的生長發(fā)育[19]。其他多種植物如柳枝稷(Panicumvirgatum)[11]、煙草[2,14]、白楊(Populustomentosa)[20]、高粱(Sorghumbicolor)[21]等的CAD基因和木質(zhì)素合成的關(guān)系也得到了初步的研究。然而,結(jié)縷草等暖季型草坪草的CAD基因的研究卻鮮有報道。
在本研究中,我們首次對結(jié)縷草CAD基因家族16個ZjCAD基因的系統(tǒng)進(jìn)化關(guān)系、基因結(jié)構(gòu)和組織表達(dá)特異性進(jìn)行了比較分析,發(fā)現(xiàn)5個ZjCAD基因在結(jié)縷草葉片中優(yōu)勢表達(dá)。在不同種源材料中,ZjCAD6基因的表達(dá)豐度和木質(zhì)素含量呈正相關(guān),提示ZjCAD6是決定結(jié)縷草葉片木質(zhì)素含量的關(guān)鍵基因。這一研究結(jié)果為解析結(jié)縷草葉片機(jī)械強(qiáng)度和耐踐踏性狀形成的分子機(jī)理提供了重要線索。
1 材料與方法
1.1 植物材料
結(jié)縷草品種(Zenith,Meyer),野生種源材料(Z103、Z112)種植于南京中山植物園試驗(yàn)地(32°02′ N,118°28′ E;海拔30 m)。采用標(biāo)準(zhǔn)的施肥、灌溉、修剪、除蟲等養(yǎng)護(hù)措施。取直立枝展開葉第二葉位的葉片用于RNA提取和木質(zhì)素染色,材料來源見表1。
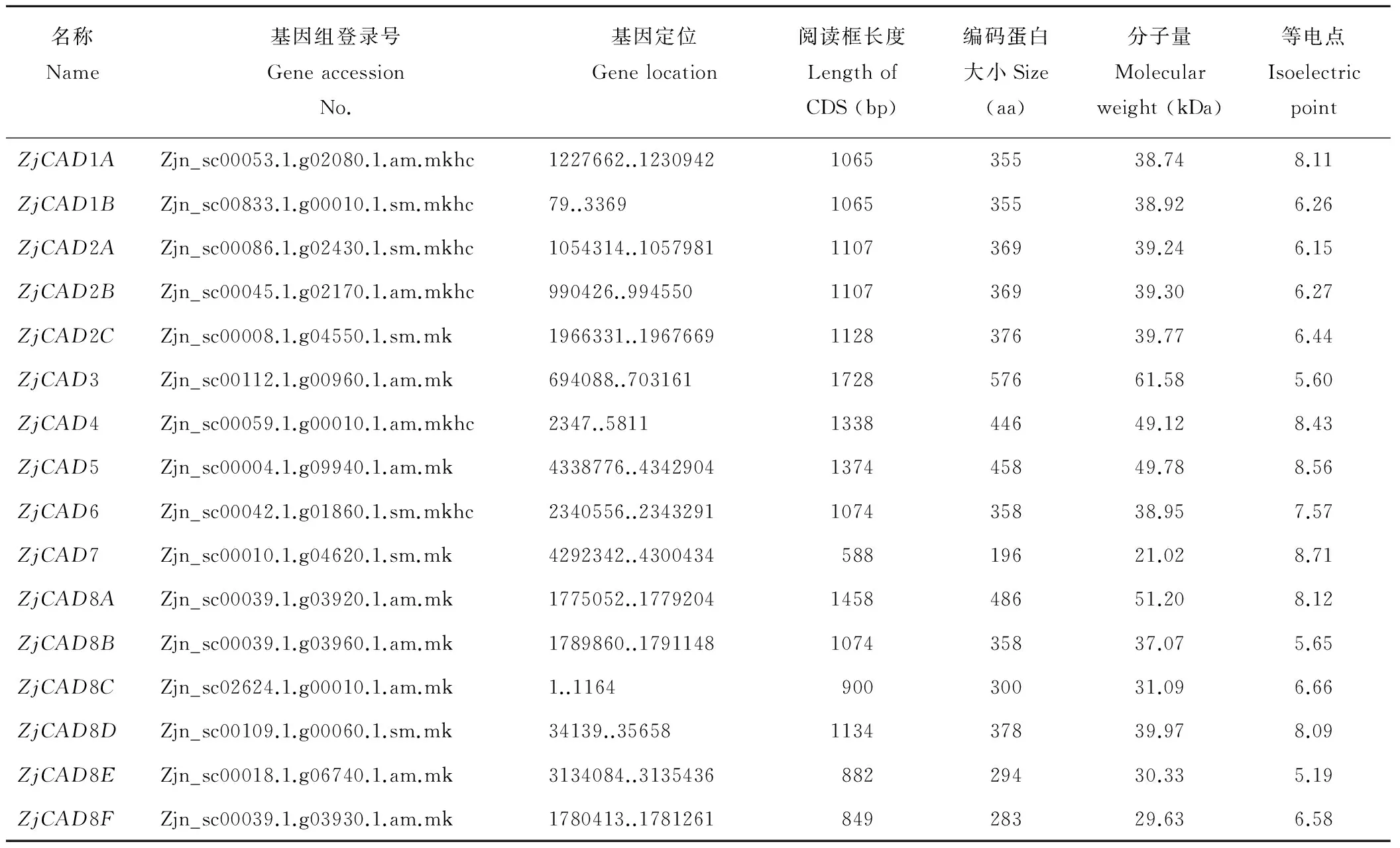
1.2CAD基因的序列獲取與生物信息學(xué)分析
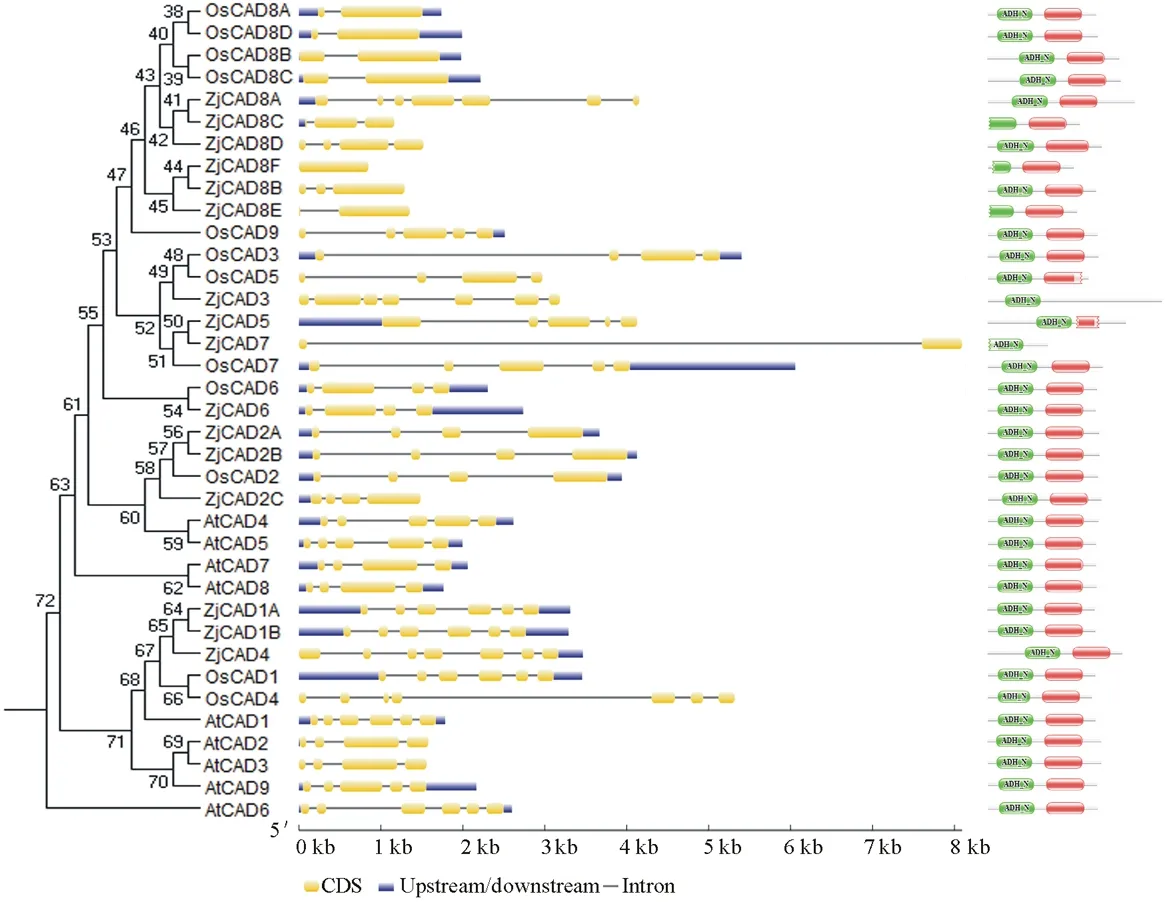
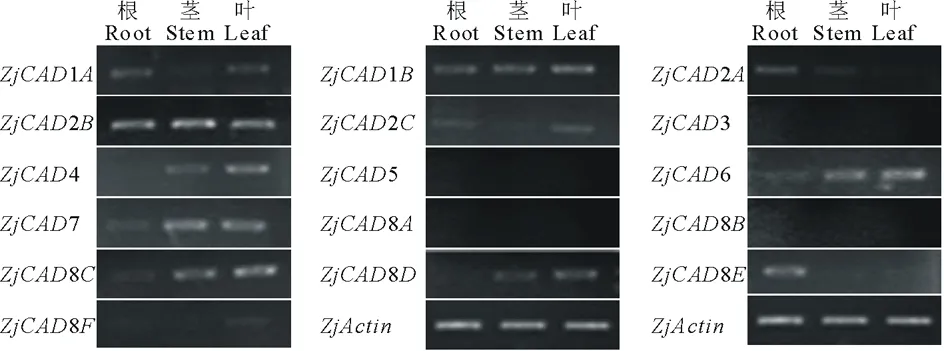
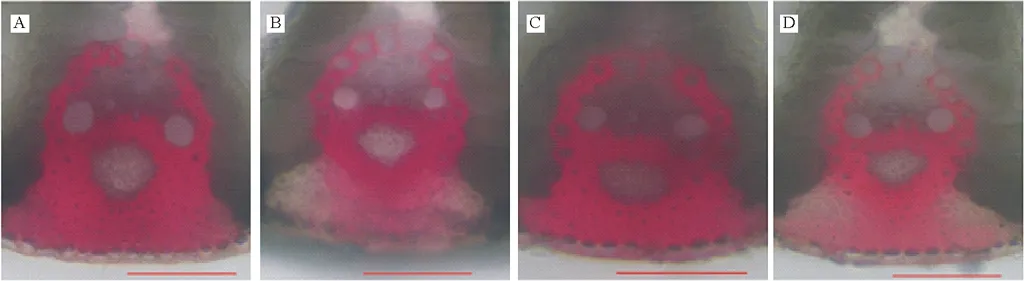
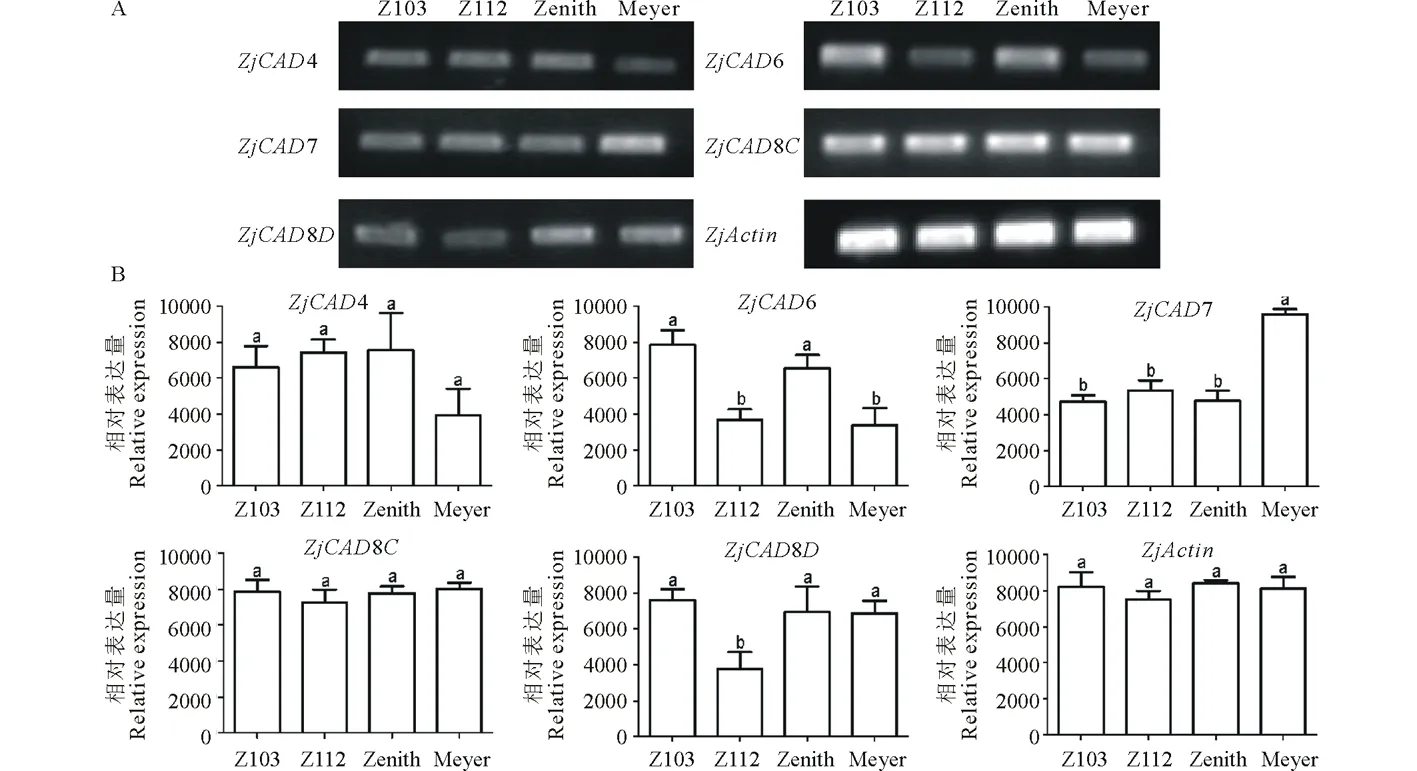
以已報道的擬南芥和水稻CAD基因?yàn)樘结榌18],通過本地同源BLAST搜索[22]結(jié)縷草基因組數(shù)據(jù)庫(http://zoysia.kazusa.or.jp/)[23],E值設(shè)定為 將獲取的結(jié)縷草ZjCAD基因與擬南芥AtCAD基因、水稻OsCAD基因進(jìn)行生物信息學(xué)分析比較。系統(tǒng)進(jìn)化樹采用Megalign軟件進(jìn)行、蛋白結(jié)構(gòu)域通過Pfam分析獲得、內(nèi)含子外顯子分析通過GSDS2.0(http://gsds.cbi.pku.edu.cn/)進(jìn)行。 1.3 RNA提取與cDNA合成 使用植物RNA提取試劑盒(Omega,USA)提取結(jié)縷草不同組織RNA,提取方法參照試劑盒說明書進(jìn)行。使用RNase-Free DNase Set(BBI,China)消化基因組DNA。使用PrimeScriptTMII 1st Strand cDNA Synthesis Kit(Takara,Japan)逆轉(zhuǎn)錄合成cDNA。 1.4ZjCAD基因在結(jié)縷草不同組織和不同種源材料的表達(dá)分析 以cDNA為模板,用特異性引物進(jìn)行PCR擴(kuò)增(引物設(shè)計見表2),以結(jié)縷草肌動蛋白actin基因?yàn)閮?nèi)參,分析不同ZjCAD基因在不同組織和不同種源材料的表達(dá)情況。 表2 ZjCAD基因表達(dá)引物Table 2 Set of primers for ZjCAD genes expression 1.5 木質(zhì)素組織化學(xué)染色 取展開葉第二葉位葉片,于葉基部1 cm處用雙面刀片切取厚度約100 μm的薄片。染色方法采用Wiesner染色法,將超薄切片置于2%間苯三酚(無水乙醇溶解)中染色2 min,置于載玻片上滴加1滴25%鹽酸,立即封片鏡檢。 2.1ZjCAD基因家族生物信息學(xué)分析 如表3所示,通過本地blast分析和Pfam結(jié)構(gòu)域確認(rèn),我們總共在結(jié)縷草基因組中發(fā)現(xiàn)了16個CAD基因,其編碼蛋白氨基酸殘基數(shù)范圍為196~576個,與玉米ZmCAD2基因相似度在42%~92%之間。為了更好地了解結(jié)縷草CAD基因家族的序列變異情況,我們將16個ZjCAD基因與12個水稻OsCAD基因和9個擬南芥AtCAD基因進(jìn)行了進(jìn)化樹的分析。如圖1所示,系統(tǒng)進(jìn)化樹分析結(jié)果表明結(jié)縷草CAD基因存在高度的重復(fù)擴(kuò)增現(xiàn)象,如AtCAD1基因在水稻中有OsCAD1和OsCAD4兩個同源基因,而在結(jié)縷草中其同源基因數(shù)量進(jìn)一步增加至3個。基因結(jié)構(gòu)分析顯示結(jié)縷草CAD基因家族不同成員內(nèi)含子數(shù)量差異巨大,所含內(nèi)含子數(shù)量從0到6均有分布。蛋白結(jié)構(gòu)域分析則進(jìn)一步表明,除ZjCAD3和ZjCAD7外所有ZjCAD基因編碼蛋白均含有1個醇脫氫酶結(jié)構(gòu)域和1個輔分子伴侶結(jié)構(gòu)。 表3 結(jié)縷草CAD基因家族信息Table 3 The information of CAD gene family in Z.japonica 2.2ZjCAD基因在不同結(jié)縷草種質(zhì)不同組織中的表達(dá)特征 利用RT-PCR方法,我們進(jìn)一步對結(jié)縷草CAD基因家族不同成員的組織表達(dá)模式進(jìn)行了分析,結(jié)果如圖2所示。在16個ZjCAD基因中,除ZjCAD3、ZjCAD5、ZjCAD8A和ZjCAD8B 4個基因外,其他12個基因均在一個或多個組織中表達(dá)。12個基因中,ZjCAD1B、ZjCAD2B為持家基因,在根、莖、葉中均高表達(dá)。ZjCAD8E在根中特異性表達(dá),ZjCAD2A在根和莖中優(yōu)勢表達(dá),而ZjCAD4、ZjCAD6、ZjCAD7、ZjCAD8C、ZjCAD8D則均在葉中有著較高表達(dá)。 2.3 不同結(jié)縷草種源材料葉片木質(zhì)素含量與CAD基因表達(dá)相關(guān)性 使用 winser 染色法,我們對4份結(jié)縷草種源材料的葉片進(jìn)行了木質(zhì)素的組織化學(xué)染色分析,如圖3所示,4份種源材料中,Z103與Zenith染色較深,Z112與Meyer染色較淺,且染色面積較小,說明Z103與Zenith木質(zhì)素含量高于Z112與Meyer。如圖4所示,對上述分析發(fā)現(xiàn)的5個葉片中高表達(dá)的CAD基因進(jìn)行RT-PCR分析發(fā)現(xiàn),僅ZjCAD6的表達(dá)模式和木質(zhì)素染色結(jié)果一致,即Z103與Zenith兩份種源材料的表達(dá)量高于Z112與Meyer,而其余4個CAD基因的表達(dá)則無這種相關(guān)性。這一結(jié)果提示ZjCAD6基因可能是決定不同結(jié)縷草種源材料葉片木質(zhì)素含量的關(guān)鍵基因。 圖1 結(jié)縷草ZjCAD基因家族進(jìn)化樹,基因結(jié)構(gòu)和保守結(jié)構(gòu)域分析Fig.1 Phylogenetic tree,gene structure and conserved motif analysis of ZjCAD genes 圖2 ZjCAD基因在不同組織中的表達(dá)情況Fig.2 The expression pattern of ZjCAD genes in different tissues 圖3 木質(zhì)素組織化學(xué)染色Fig.3 Histochemical analysis of ligninA:Z103;B:Z112;C:Zenith;D:Meyer.Scale bar=50 μm. 圖4 5個ZjCAD基因在4份材料中的差異表達(dá)Fig.4 Expression analysis of 5 ZjCAD genes in 4 zoysiagrassesA:代表性半定量RT-PCR電泳結(jié)果Representative electrophoresis result of RT-PCR;B:3次重復(fù)實(shí)驗(yàn)統(tǒng)計學(xué)分析Statistical analysis of 3 replicate experiments. 圖5 ZjCAD6基因與其他物種CAD基因氨基酸序列一致性比對Fig.5 Alignment of amino acid sequence of ZjCAD6 with CADs from other plant species 2.4 ZjCAD6蛋白序列保守性分析 ZjCAD6基因編碼蛋白ZjCAD6為有催化活性的功能蛋白。為了進(jìn)一步確定ZjCAD6基因與結(jié)縷草葉片木質(zhì)素合成的相關(guān)性,我們將ZjCAD6蛋白與其他植物具催化活性的CAD蛋白進(jìn)行了氨基酸序列比對分析。如圖5所示,所有具催化活性的CAD蛋白均含一個與Zn2+催化相關(guān)的GHE(X)2G(X)5G(X)2V結(jié)構(gòu)域,一個與Zn2+結(jié)合的GD(X)10C(X)2C(X)2C(X)7C結(jié)構(gòu)域和一個與NADP(H)結(jié)合相關(guān)的富含甘氨酸的GLGGV(L)G保守結(jié)構(gòu)域[25]。這3個結(jié)構(gòu)域在ZjCAD6蛋白中均同樣存在,提示ZjCAD6基因編碼蛋白ZjCAD6為有催化活性的功能蛋白。 肉桂醇脫氫酶以基因家族的形式存在于多個植物中,在擬南芥中有9個CAD基因,水稻中有12個CAD基因,二穗短柄草(Brachypodiumdistachyum)和楊樹(Populustrichocarpa)中則分別有7和15個CAD基因[20,26]。在結(jié)縷草中,我們共發(fā)現(xiàn)了16個CAD基因。這16個CAD基因具有非常大的序列和基因結(jié)構(gòu)差異,最短的ZjCAD8F僅849 bp,最長的ZjCAD7則長達(dá)8093 bp,ZjCAD8F無內(nèi)含子,而ZjCAD3、ZjCAD4、ZjCAD8A則擁有多達(dá)6個的內(nèi)含子。這一結(jié)果和擬南芥、水稻中CAD基因家族不同成員的序列差異度是不同的[27],說明結(jié)縷草CAD基因家族的不同成員具有較大的基因序列差異性[26]。擬南芥中,AtCAD4(AtCAD-D)、AtCAD5(AtCAD-C)為控制木質(zhì)素合成的主效基因,AtCAD4在花和根中高表達(dá)。AtCAD5在木質(zhì)的根中高表達(dá)[28]。在二穗短柄草中,BdCAD5則在根和莖中高表達(dá)。本研究同樣發(fā)現(xiàn)不同結(jié)縷草CAD基因具有不同的組織表達(dá)特異性,值得一提的是ZjCAD4、ZjCAD6、ZjCAD7、ZjCAD8C、ZjCAD8D5個基因在葉片中均高表達(dá),提示這5個CAD基因參與了結(jié)縷草葉片木質(zhì)素的合成。然而,在擬南芥中,AtCAD1、AtCAD7、AtCAD8等基因也被證明與木質(zhì)素的合成相關(guān)[17,20,28],說明CAD基因家族不同成員可能存在功能冗余的現(xiàn)象。因此,葉片中高表達(dá)的5個ZjCAD基因的具體功能作用尚需進(jìn)一步探究和驗(yàn)證。 Wiesner反應(yīng)利用間苯三酚在酸性條件下與肉桂醛基團(tuán)發(fā)生顯色反應(yīng)獲得紅色或粉色的化合物,是檢測植物木質(zhì)素含量的最直觀、高效的方法[29]。本研究中,Z103與Zenith兩份結(jié)縷草種源材料Wiesner染色強(qiáng)度高于Z112與Meyer,說明Z103與Zenith葉片木質(zhì)素含量和木質(zhì)化程度較高,提示不同種源材料機(jī)械強(qiáng)度和耐踐踏能力差異可能與其木質(zhì)化程度密切相關(guān)。 不同結(jié)縷草種源材料中ZjCAD6基因的表達(dá)和葉片木質(zhì)素含量正相關(guān),提示其為決定結(jié)縷草葉片木質(zhì)素含量的關(guān)鍵CAD基因。在幾種模式植物中,AtCAD4、AtCAD5[16]、ZmCAD2[18]、OsCAD2[13]基因分別為控制擬南芥、玉米、水稻木質(zhì)素合成的主效基因,序列分析發(fā)現(xiàn)ZjCAD6基因與這些具催化活性的CAD基因具有46%~47%的序列相似性,尤其是與催化活性相關(guān)的酶結(jié)構(gòu)域高度保守,說明ZjCAD6蛋白是具有CAD酶活性的活性蛋白。在進(jìn)化樹的比對中,ZjCAD6與OsCAD6相似度最高,盡管OsCAD6未被證明直接參與木質(zhì)素的合成過程,但其特有的過氧化物酶結(jié)合位點(diǎn)揭示其可能參與了其他代謝途徑。而且,OsCAD6與楊樹中芥子醇脫氫酶(sinapyl alcohol dehydrogenase,SAD)有59.4%的相似度,SAD表現(xiàn)出對底物芥子醛特異的親和性[27]。因此,在今后的研究中,可進(jìn)一步對ZjCAD6基因進(jìn)行表達(dá)、純化與活性測定,分析其催化特征和調(diào)控機(jī)制,從而可以更深入的解析結(jié)縷草葉片木質(zhì)素合成的分子機(jī)理。 本研究首次獲得了結(jié)縷草16個CAD基因家族成員。16個結(jié)縷草CAD基因中,有12個基因在根、莖、葉不同組織中表達(dá),5個基因在葉片中優(yōu)勢表達(dá)。5個基因中,僅ZjCAD6基因的表達(dá)與不同結(jié)縷草種源材料木質(zhì)素含量正相關(guān),提示ZjCAD6基因是控制結(jié)縷草葉片木質(zhì)素合成的關(guān)鍵基因。本研究結(jié)果為解析結(jié)縷草機(jī)械強(qiáng)度和耐踐踏的分子機(jī)理提供了重要信息。 References: [1] Lulli F,Guglielminetti L,Grossi N,etal.Physiological and morphological factors influencing leaf,rhizome and stolon tensile strength in C4turfgrass species.Functional Plant Biology,2011,38(11):919-926. [2] Ralph J,Lapierre C,Marita J M,etal.Elucidation of new structures in lignins of CAD- and COMT-deficient plants by NMR.Phytochemistry,2001,57(6):993-1003. [3] Barrière Y,Riboulet C,Méchin V,etal.Genetics and genomics of lignification in grass cell walls based on maize as model species.Genes Genomes Genomics,2007,1(2):133-156. [4] Harrington M J,Mutwil M,Barrière Y,etal.Molecular biology of lignification in grasses.Advances in Botanical Research,2012,61:77-112. [5] Huang J,Gu M,Lai Z,etal.Functional analysis of theArabidopsisfunctional analysis of theArabidopsisPAL gene family in plant growth,development,and response to environmental stress.Plant Physiology,2010,153(4):1526-1538. [6] Schilmiller A L,Stout J,Weng J K,etal.Mutations in the cinnamate 4-hydroxylase gene impact metabolism,growth and development inArabidopsis.The Plant Journal,2009,60(5):771-782. [7] Xu B,Escamilla-Trevio L L,Sathitsuksanoh N,etal.Silencing of 4-coumarate:coenzyme A ligase in switchgrass leads to reduced lignin content and improved fermentable sugar yields for biofuel production.New Phytologist,2011,192(3):611-625. [8] Franke R,Hemm M R,Denault J W,etal.Changes in secondary metabolism and deposition of an unusual lignin in the ref8 mutant ofArabidopsis.The Plant Journal,2002,30(1):47-59. [9] Pichon M,Deswartes C,Gerentes D,etal.Variation in lignin and cell wall digestibility in caffeic acid O-methyltransferase down-regulated maize half-sib progenies in field experiments.Molecular Breeding,2006,18(3):253-261. [10] Ruegger M,Meyer K,Cusumano J C,etal.Regulation of ferulate-5-hydroxylase expression inArabidopsisin the context of sinapate ester biosynthesis.Plant Physiology,1999,119(1):101-110. [11] Saathoff A J,Sarath G,Chow E K,etal.Downregulation of cinnamyl-alcohol dehydrogenase in switchgrass by RNA silencing results in enhanced glucose release after cellulase treatment.PloS one,2011,6(1):e16416. [12] Ookawa T,Inoue K,Matsuoka M,etal.Increased lodging resistance in long-culm,low-lignin gh2 rice for improved feed and bioenergy production.Scientific Reports,2014,4:6567. [13] Hirano K,Aya K,Kondo M,etal.OsCAD2 is the majorCADgene responsible for monolignol biosynthesis in rice culm.Plant Cell Reports,2012,31(1):91-101. [14] Knight M E,Halpin C,Schuch W.Identification and characterization of cDNA clones encoding cinnamyl alcohol dehydrogenase from tobacco.Plant Molecular Biology,1992,19(5):793-801. [15] Sibout R,Eudes A,Pollet B,etal.Expression pattern of two paralogs encoding cinnamyl alcohol dehydrogenases inArabidopsis.Isolation and characterization of the corresponding mutants.Plant Physiology,2003,132(2):848-860. [16] Sibout R,Eudes A,Mouille G,etal.Cinnamyl alcohol dehydrogenase-C and-D are the primary genes involved in lignin biosynthesis in the floral stem ofArabidopsis.The Plant Cell,2005,17(7):2059-2076. [17] Eudes A,Pollet B,Sibout R,etal.Evidence for a role of AtCAD 1 in lignification of elongating stems ofArabidopsisthaliana.Planta,2006,225(1):23-39. [18] Halpin C,Holt K,Chojecki J,etal.Brown-midrib maize (bm1)-a mutation affecting the cinnamyl alcohol dehydrogenase gene.The Plant Journal,1998,14(5):545-553. [19] Fornalé S,Capellades M,Encina A,etal.Altered lignin biosynthesis improves cellulosic bioethanol production in transgenic maize plants down-regulated for cinnamyl alcohol dehydrogenase.Molecular Plant,2012,5(4):817-830. [20] Barakat A,Bagniewska-Zadworna A,Choi A,etal.The cinnamyl alcohol dehydrogenase gene family inPopulus:phylogeny,organization,and expression.BMC Plant Biology,2009,9(1):26. [21] Sattler S E,Saathoff A J,Haas E J,etal.A nonsense mutation in a cinnamyl alcohol dehydrogenase gene is responsible for the sorghum brown midrib6 phenotype.Plant Physiology,2009,150(2):584-595. [22] Altschul S F,Gish W,Miller W,etal.Basic local aligment search tool.Journal of Molecular Biology,1990,215(3):403-410. [23] Tanaka H,Hirakawa H,Kosugi S,etal.Sequencing and comparative analyses of the genomes of zoysiagrasses.DNA Research,2016,23(2):171-180. [24] Xie Q,Niu J,Xu X,etal.Corrigendum:De novo assembly of the Japanese lawngrass (ZoysiajaponicaSteud.) root transcriptome and identification of candidate unigenes related to early responses under salt stress.Frontiers in Plant Science,2015,6:610. [25] Chao N,Liu S X,Liu B M,etal.Molecular cloning and functional analysis of nine cinnamyl alcohol dehydrogenase family members inPopulustomentosa.Planta,2014,240(5):1097-1112. [26] Bukh C,Nord-Larsen P H,Rasmussen S K.Phylogeny and structure of the cinnamyl alcohol dehydrogenase gene family inBrachypodiumdistachyon.Journal of Experimental Botany,2012,63(17):6223-6236. [27] Tobias C M,Chow E K.Structure of the cinnamyl-alcohol dehydrogenase gene family in rice and promoter activity of a member associated with lignification.Planta,2005,220(5):678-688. [28] Tronchet M,Balagué C,Kroj T,etal.Cinnamyl alcohol dehydrogenases-C and D,key enzymes in lignin biosynthesis,play an essential role in disease resistance inArabidopsis.Molecular Plant Pathology,2010,11(1):83-92. [29] Vallet C,Chabbert B,Czaninski Y,etal.Histochemistry of lignin deposition during sclerenchyma differentiation in alfalfa stems.Annals of Botany,1996,78(5):625-632. Genome-wide identification and characterization of the cinnamyl alcohol dehydrogenase gene family inZoysiajaponica SHI Jing-Ang,ZHANG Bing*,XIAO Xiao-Lin,MA Jing-Jing,YANG Xiang-Yang,LIU Jian-Xiu InstituteofBotany,JiangsuProvinceandChineseAcademyofSciences,Nanjing210014,China Cinnamyl alcohol dehydrogenase (CAD,EC1.1.1.195) is a key enzyme in the lignin biosynthesis pathway.By using NADPH as a cofactor,CAD catalyzes cinnamyl aldehydes to alcohols and plays an important role in plant development and stress responses.This study identified 16 CAD genes inZoysiajaponica.Phylogenetic and structural analyses of these genes were conducted.Semi-quantitative RT-PCR indicated that 5ZjCADgenes were specifically expressed in leaves.Lignin histochemical staining and expression analyses ofZjCADgenes in different cultivars and a wild accession further showed that the expression pattern ofZjCAD6 was positively correlated with leaf lignin contents.These results indicate thatZjCAD6 may be the key gene that regulates lignin biosynthesis in the leaves ofZ.japonica.The full length of theZjCAD6 gene is 2736 bp,and is comprised of three introns and four exons.The 1074 bp open reading frame ofZjCAD6 encodes 357 amino acids.ZjCAD6 protein contains two Zn2+-binding motifs and a NADP(H)-binding motif,which is vital for its catalytic activity.Our results provide an important reference for further study of both lignin content's molecular mechanisms and the tensile strength ofZ.japonicaleaves. Zoysiajaponica;lignin;cinnamyl alcohol dehydrogenase;bioinformatics analysis;gene expression 10.11686/cyxb2016442 http://cyxb.lzu.edu.cn 史經(jīng)昂,張兵,肖曉琳,馬晶晶,楊向陽,劉建秀.結(jié)縷草肉桂醇脫氫酶基因家族全基因組序列鑒定和表達(dá)分析.草業(yè)學(xué)報,2017,26(6):111-119. SHI Jing-Ang,ZHANG Bing,XIAO Xiao-Lin,MA Jing-Jing,YANG Xiang-Yang,LIU Jian-Xiu.Genome-wide identification and characterization of the cinnamyl alcohol dehydrogenase gene family inZoysiajaponica.Acta Prataculturae Sinica,2017,26(6):111-119. 2016-11-19;改回日期:2017-02-14 江蘇省公益院所能力提升項目-草坪種質(zhì)創(chuàng)新(BM2015019-1),江蘇省體育產(chǎn)業(yè)引導(dǎo)項目“江蘇省運(yùn)動草坪研究中心建設(shè)暨運(yùn)動草坪標(biāo)準(zhǔn)化項目”和江蘇省體育產(chǎn)業(yè)引導(dǎo)項目“天然草坪耐踐踏性關(guān)鍵技術(shù)體系研發(fā)與集成示范”資助。 史經(jīng)昂(1992-),男,江蘇徐州人,在讀碩士。E-mail:shion1992@outlook.com *通信作者Corresponding author.E-mail:bingkeith@gmail.com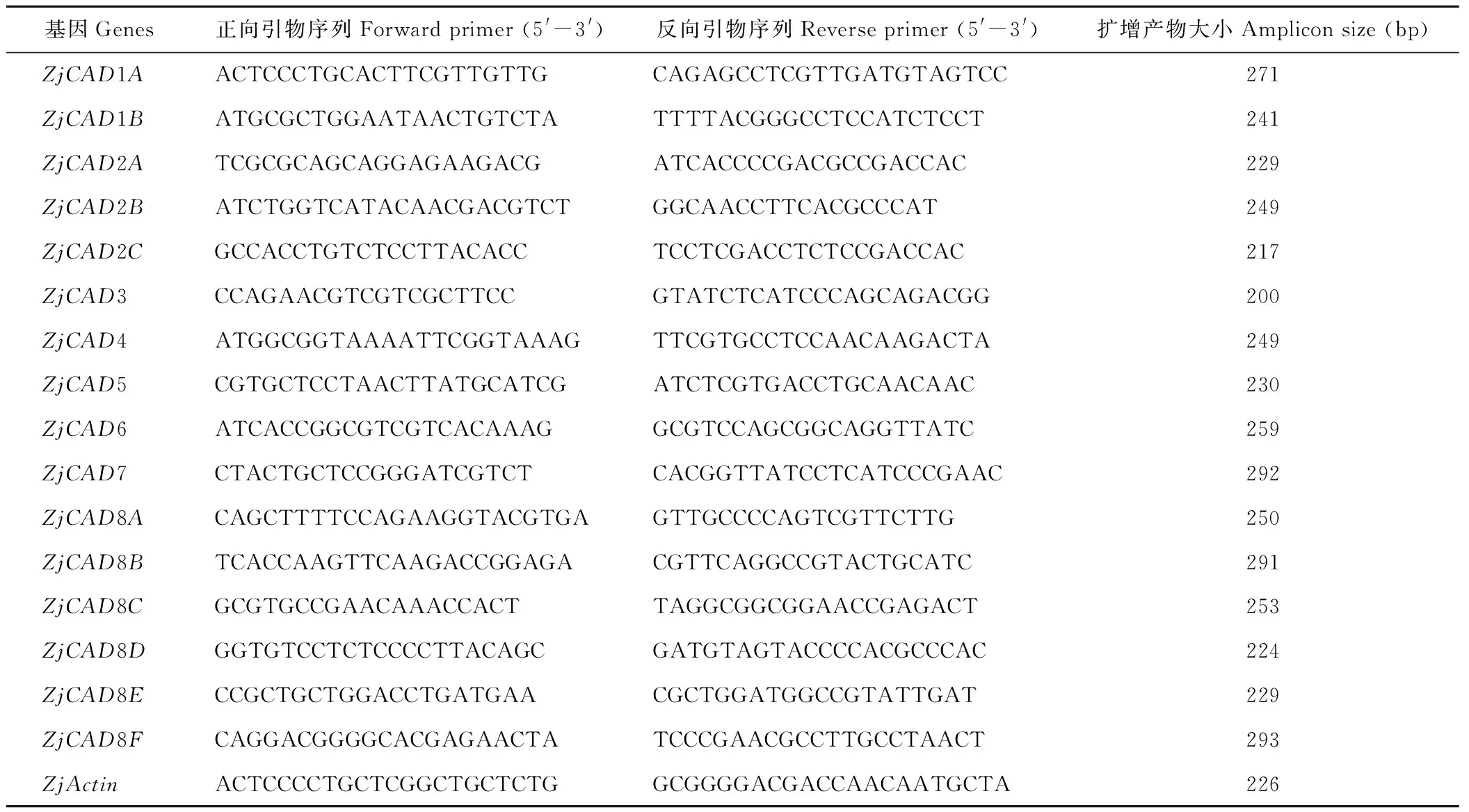
2 結(jié)果與分析

3 討論
4 結(jié)論
猜你喜歡
現(xiàn)代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機(jī)設(shè)計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
當(dāng)代經(jīng)濟(jì)研究(2016年5期)2016-12-01 03:12:05
現(xiàn)代農(nóng)業(yè)(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫(yī)藥現(xiàn)代遠(yuǎn)程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學(xué)學(xué)報(社會科學(xué)版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
