代謝工程改造大腸桿菌合成β-丙氨酸
2017-06-21 15:10:24梁姍姍周麗張斌周哲敏
食品與發(fā)酵工業(yè) 2017年5期
關(guān)鍵詞:優(yōu)化
梁姍姍,周麗,張斌,周哲敏
(江南大學(xué) 生物工程學(xué)院,工業(yè)生物技術(shù)教育部重點(diǎn)實(shí)驗(yàn)室,江蘇 無錫,214122)
代謝工程改造大腸桿菌合成β-丙氨酸
梁姍姍,周麗,張斌,周哲敏*
(江南大學(xué) 生物工程學(xué)院,工業(yè)生物技術(shù)教育部重點(diǎn)實(shí)驗(yàn)室,江蘇 無錫,214122)
β-丙氨酸是天然存在的唯一一種β型氨基酸,在醫(yī)藥、食品、化工等領(lǐng)域有重要應(yīng)用。該研究考察了以重組Escherichiacoli發(fā)酵合成β-丙氨酸的可能性。在敲除副產(chǎn)物乙酸、甲酸、乙醇、琥珀酸、乳酸合成途徑編碼基因的EscherichiacoliCICIM B0016-050 (ackA-ptapflBadhEfrdAldhA)菌株中,考察疊加敲除β-丙氨酸的競爭途徑天冬氨酸激酶、泛酸合成酶和葡萄糖轉(zhuǎn)運(yùn)蛋白(EⅡCBGlc)的編碼基因(lysC、panC、ptsG)以及過表達(dá)來源于Corynebacteriumglutamicum的panD基因?qū)铣搔?丙氨酸的影響。結(jié)果表明,疊加敲除上述基因后,β-丙氨酸的合成量依次提高了12.5%、39.3%和13.3%;過量表達(dá)panD基因,β-丙氨酸合成量提高了86.2倍;經(jīng)發(fā)酵條件優(yōu)化,菌株 B0016-080BB/pPL-panD搖瓶發(fā)酵可合成(5.0±0.2) g/L β-丙氨酸,體積生產(chǎn)強(qiáng)度達(dá)到(0.12±0.01) g/(L·h)。該結(jié)果為發(fā)酵法合成β-丙氨酸奠定了基礎(chǔ)。
β-丙氨酸;大腸桿菌;L-天冬氨酸-α-脫羧酶;代謝工程;基因敲除
β-丙氨酸即3-氨基丙酸,是自然界中唯一存在的β型氨基酸,近年來,在醫(yī)藥、化工、食品、環(huán)境等領(lǐng)域的作用日漸突出[1]。在醫(yī)藥領(lǐng)域,主要用于合成泛酸以及泛酸鈣,此外,β-丙氨酸還是合成肌肽(camosine)、帕米酸鈉(pamidronate,第二代抗骨溶解藥)、巴柳氮(balsalazide)等藥物的原料;在化工領(lǐng)域,用于藥劑制備中的沉淀劑,電鍍緩蝕劑,鉛中毒解毒劑等;在食品領(lǐng)域,可用作調(diào)味品;在環(huán)境領(lǐng)域用于水的凈化絮凝劑等[2]。有報道稱β-丙氨酸是未來全球12種最具開發(fā)潛力的三碳化工產(chǎn)品之一[3]。
國內(nèi)外生產(chǎn)β-丙氨酸主要采用化學(xué)合成法,包括丙烯酸法、丙烯腈法、β-氨基丙腈法、β-氨基丙醇法等,這些方法合成工藝流程長,且需高溫、高壓,能耗大、生產(chǎn)過程對環(huán)境有一定的毒害作用。β-丙氨酸生物生產(chǎn)主要是采用微生物如產(chǎn)堿桿菌屬Alcaligenessp. OMT-MY14、氨基桿菌屬AminobacteraminobranceATCC 23314和紅球菌Rhodococcussp. G20[4],將β-氨基丙腈進(jìn)行全細(xì)胞生物轉(zhuǎn)化;KONST等人還利用L-天門冬氨酸-α-脫羧酶的酶轉(zhuǎn)化法制備β-丙氨酸[5-6],成為近年來研究的熱點(diǎn)[6],但其生產(chǎn)需要昂貴的前體和大量酶,還存在著高濃度底物抑制菌生長、產(chǎn)率較低等問題,不適合大規(guī)模工業(yè)化生產(chǎn)。Chan Woo Song等[7]構(gòu)建了菌株E.coliCWF4NA2 (ΔiclRΔfumCΔfumAΔfumBΔptsGΔlacIPaspA::Ptrc Pacs::Ptrc),首次采用發(fā)酵法在E.coli中合成β-丙氨酸,搖瓶發(fā)酵產(chǎn)量達(dá)3.94 g/L,但并未敲除有機(jī)酸、醇等副產(chǎn)物的合成途徑,這些副產(chǎn)物的積累會導(dǎo)致β-丙氨酸得率降低,同時影響產(chǎn)物的化學(xué)純度;此外,也未考察β-丙氨酸競爭代謝途徑(lysC)和分解代謝途徑(panC)的敲除對β-丙氨酸合成的影響;另外,以葡萄糖為碳源時,敲除ptsG[8]可顯著增強(qiáng)丙酮酸激酶(Pck)和磷酸烯醇式丙酮酸羧化酶(Ppc)的活性,從而提高草酰乙酸的積累量,而該基因在其它碳源存在時對β-丙氨酸合成的影響也未有報道。ZHOU等用PL啟動子成功地對乳酸脫氫酶[9]和丙氨酸脫氫酶[10]進(jìn)行開關(guān)控制,僅通過溫控調(diào)節(jié),在重組E.coli中實(shí)現(xiàn)了D-乳酸和L-丙氨酸的高產(chǎn),將該方法應(yīng)用于β-丙氨酸合成重組菌株,也有望大幅度提高β-丙氨酸的合成水平。
本文在實(shí)驗(yàn)室已有菌株E.coliB0016-050[11](敲除副產(chǎn)物乙酸、甲酸、乙醇、琥珀酸、乳酸合成途徑編碼基因ackA-pta、pflB、adhE、frdA、ldhA)的基礎(chǔ)上,敲除lysC、panC和ptsG基因,并將來源于Corynebacteriumglutamicum的panD基因克隆于溫控強(qiáng)啟動子PL下游,在上述重組菌株中進(jìn)行高效表達(dá),考察這些代謝途徑的改造對發(fā)酵甘油合成β-丙氨酸的影響。

圖1 E.coli菌株β-丙氨酸合成相關(guān)代謝途徑及改造策略Fig.1 Metabolic pathway and strategy for β- alanine production in E.coli注:基因及其編碼的酶: pflB, 丙酮酸甲酸裂解酶; ldhA, NAD依賴發(fā)酵型D-乳酸脫氫酶; pta, 磷酸轉(zhuǎn)乙酰酶; ackA, 乙酸激酶; adhE, 乙醇脫氫酶; frdA, 富馬酸還原酶; aspA, 天冬氨酸氨裂解酶; lysC, 天冬氨酸激酶; panC, 泛酸合成酶; ptsG, 主要的葡萄糖轉(zhuǎn)運(yùn)蛋白IICB(Glc); ppc, 磷酸烯醇丙酮酸羧化酶; aspC, 天冬氨酸轉(zhuǎn)氨酶; panD, 天冬氨酸-α-脫羧酶·黑點(diǎn)代表本研究所刪除的基因
1 材料與方法
1.1 菌株與引物
1.1.1 菌株和質(zhì)粒
本文所用到的菌株、質(zhì)粒如表1所示。
E.coliB0016-050菌株為在野生型菌株E.coliCICIM B0016中敲除了基因ackA-pta、pflB、adhE、frdA、ldhA的重組菌株[11],由本實(shí)驗(yàn)室構(gòu)建、保藏。其他菌株由本文構(gòu)建。
1.1.2 引物
本文所用到的引物如表2所示。
1.2 培養(yǎng)基
(1)LB培養(yǎng)基(g/L):胰蛋白胨10;酵母浸出粉5;NaCl 10(固體培養(yǎng)基,瓊脂15)。
(2)M9-1液體培養(yǎng)基(g/L):NaHPO4·12H2O 15.1;KH2PO43.0;NH4Cl 1.0;NaCl 0.5;(NH4)2SO413.21,補(bǔ)加50% (v/v)甘油 10 mL,200 g/L泛酸1 mL,0.1% (v/v) MgSO41 mol/L和0.1% (v/v)的微量元素母液。微量元素成分為(g/L):FeCl3·6H2O 2.4;CoCl2·6H2O 0.3;CuCl2·2H2O 0.15;ZnCl20.3;Na2MO4·2H2O 0.3;H3BO30.075;MnCl2·4H2O 0.495。
1.3 基因敲除方法
用 DATSENKO 等[12]報道的Red 重組系統(tǒng)基因敲除方法依次敲除B0016-050菌株染色體上的lysC、panC、和ptsG基因。其具體過程為: 以pKD13質(zhì)粒為模板,分別用引物對LysC-PKD13R/LysC-PKD13F、PanC-PKD13R/PanC-PKD13F和PtsG -PKD13R/PtsG-PKD13F進(jìn)行PCR 擴(kuò)增,獲得兩端各具有50 bp 同源臂序列、中部為FRT-Kan-FRT 的突變盒片段。借助質(zhì)粒pKD46 上的Red重組系統(tǒng)將突變盒整合于待突變菌株的染色體上,并用相應(yīng)的驗(yàn)證引物對(YlysCF/YlysCR,YpanCF/YpanCR,YptsGF/YptsGR)進(jìn)行菌落PCR 驗(yàn)證,最后用pCP20質(zhì)粒去除重組菌中的卡那抗性基因,并進(jìn)一步用相應(yīng)驗(yàn)證引物進(jìn)行驗(yàn)證。

表1 本研究中所使用的菌株和質(zhì)粒
注:a:江南大學(xué)中國高校工業(yè)微生物資源和信息中心;b:耶魯大學(xué)基因保藏中心。

表2 本研究中所使用的引物
注a:小寫字母表示與pKD13質(zhì)粒同源堿基序列。
1.4 搖瓶發(fā)酵培養(yǎng)條件
種子培養(yǎng):取-80 ℃甘油保藏的菌種,在氨芐青霉素抗性的LB平板劃線,于33 ℃ 恒溫培養(yǎng)24 h后,轉(zhuǎn)接到 50 mL含有氨芐青霉素的LB液體培養(yǎng)基中,于33 ℃搖床200 r/min振蕩培養(yǎng)至OD600=2.5。
發(fā)酵培養(yǎng):將LB種子培養(yǎng)基轉(zhuǎn)接至50 mL M9-1培養(yǎng)基中,使OD600終濃度為0.05,于33 ℃、200 r/min振蕩培養(yǎng)至OD600=2.5(優(yōu)化時OD600取:0、0.5、1、1.5、2、2.5、3、3.5、4),補(bǔ)料1.5 mL 50% (v/v)甘油并用100 g/L的NaHCO3調(diào)節(jié)pH=7,同時調(diào)節(jié)溫度至42 ℃誘導(dǎo),繼續(xù)200 r/min振蕩培養(yǎng),每6 h補(bǔ)加1 mL 50% (v/v)甘油,并用100 g/L的NaHCO3調(diào)節(jié)pH=7,48 h結(jié)束搖瓶發(fā)酵,每次試驗(yàn)采用3個平行并計(jì)算其平均值。
1.5 β-丙氨酸含量測定
發(fā)酵液預(yù)處理方法:發(fā)酵液樣品10 000 r/min離心2 min,取上清液加入等體積100 g/L三氯乙酸,避光放置大于3 h,適當(dāng)稀釋后用0.45 μm濾膜過膜。利用鄰苯二甲醛(OPA)試劑在線衍生β-丙氨酸,用高效液相色譜(HPLC)分析β-丙氨酸的含量。色譜柱為Diamonsil C18柱(250 cm×4.6 mm, 5 μm),流動相A為:無水醋酸鈉(分析純)4.52 g,先溶于水,再加三乙胺 200 μL,四氫呋喃5 mL,水定容至1 L,醋酸調(diào)節(jié)pH 7.2;流動相B為:無水醋酸鈉4.52 g 水定容為200 mL,醋酸調(diào)節(jié)pH 7.2,加400 mL甲醇和400 mL乙腈。采用梯度洗脫:0~27 min,B液由8%上升到60%,流速為1 mL/min;20~31.5 min,B液由60%上升到100%,流速為1 mL/min ;32~34 min,B液梯度不變,流速為1.2 mL/min。34~35 min,B液由100%下降到8%,流速為1 mL/min。柱溫為40 ℃,檢測波長為338 nm;進(jìn)樣量為10 μL。
2 結(jié)果和分析
2.1lysC、panC和ptsG基因的敲除
用 DATSENDO 等[12]報道的基因敲除方法,在菌株E.coliB0016-050中依次疊加敲除天冬氨酸激酶、泛酸合成酶和葡萄糖轉(zhuǎn)運(yùn)蛋白(EⅡCBGlc)的編碼基因lysC、panC和ptsG(圖1),依次獲得了B0016-060A、B0016-070B和B0016-080BB菌株(表1)。用驗(yàn)證引物對上述突變進(jìn)行驗(yàn)證,結(jié)果如圖2所示,lysC敲除前基因大小為1 714 bp,敲除后為469 bp;panC敲除前基因大小為1 334 bp,敲除后為587 bp;ptsG敲除前基因大小為1 770 bp,敲除后為340 bp。表明上述基因已成功刪除。
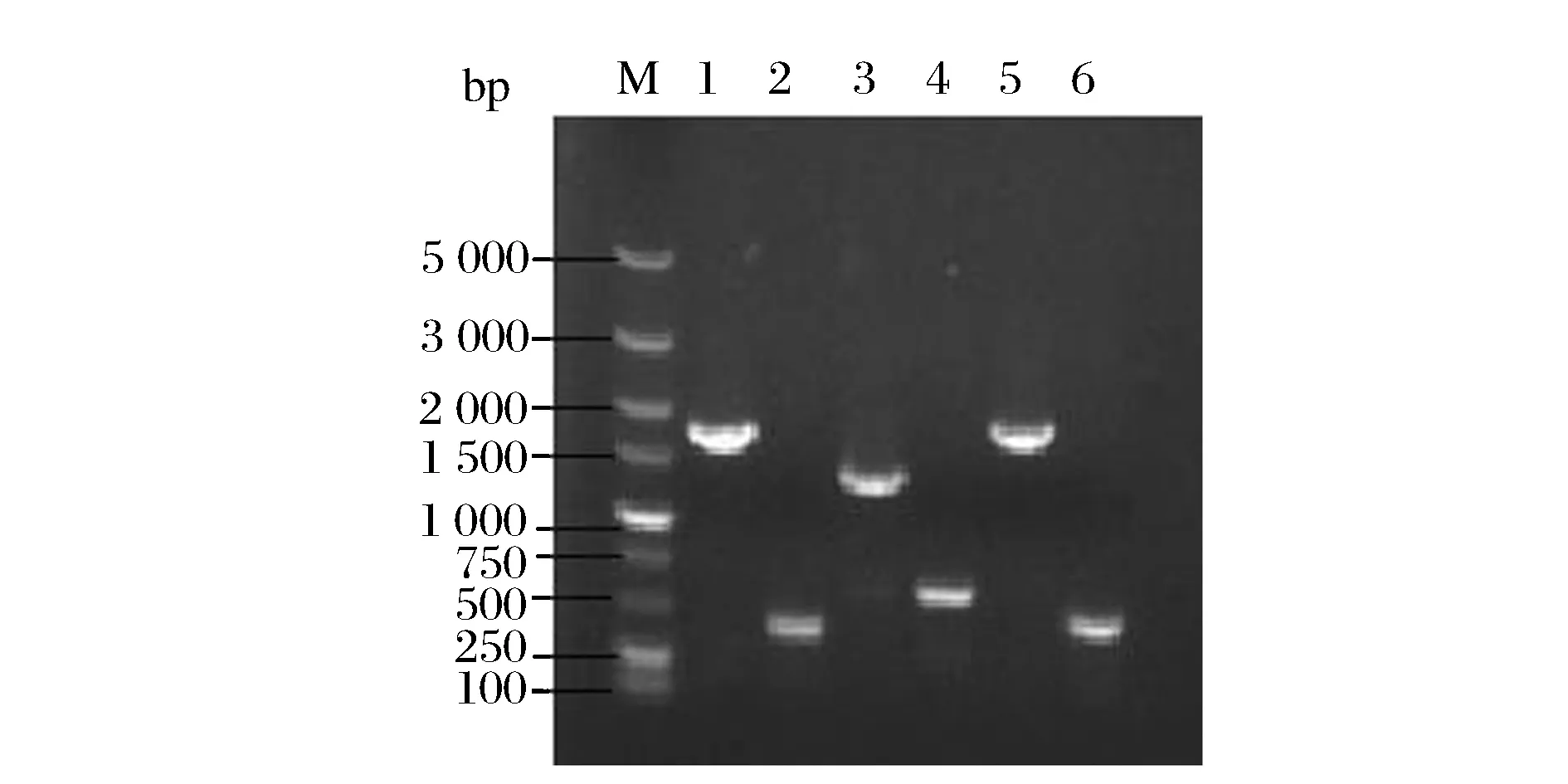
M-DL 5 000 DNA maker;1-lysC敲除前;2-lysC敲除后;3-panC敲除前;4-panC敲除后;5-ptsG敲除前;6-ptsG敲除后圖2 基因敲除PCR驗(yàn)證電泳圖Fig.2 PCR verification of gene knockouts
2.2 pPL-panD重組質(zhì)粒的構(gòu)建
本實(shí)驗(yàn)室前期構(gòu)建的質(zhì)粒pET28a-panD[14]用BglⅡ和EcoR I進(jìn)行雙酶切,膠回收0.5 kb片段,克隆于pPL451質(zhì)粒載體的BamH I和EcoR I酶切位點(diǎn),獲得重組質(zhì)粒pPL-panD,其物理圖譜如圖3A所示。重組質(zhì)粒pPL-panD用EcoR I進(jìn)行酶切,電泳獲得了4.8 kb 條帶(圖3B),表明質(zhì)粒構(gòu)建正確。

M-DL 10 000 DNA maker;1- pPL-panD digested by EcoR I圖3 pPL-panD重組質(zhì)粒的物理圖譜(A)和酶切驗(yàn)證電泳圖譜(B)Fig.3 The map of the recombinant plasmid pPL- panD (A) and confirmation of pPL-panD by restriction digestion (B)
2.3 泛酸添加量的優(yōu)化
敲除panC基因的B0016-070B菌株為營養(yǎng)缺陷型,不能在無泛酸的基本培養(yǎng)基上生長。向M9-1培養(yǎng)基中添加不同濃度的泛酸(0、10-1、10-2、10-3、10-4、10-5g/L),菌體生長狀況如圖4所示。在不添加泛酸時,B0016-070B菌株基本不生長;添加量為10-4、10-5g/L時,菌體生長受到一定程度的抑制;泛酸添加量為10-2g/L時,B0016-070B菌體生長情況與未敲除panC基因的B0016-060A菌株的菌體生長情況相似,且再提高泛酸濃度,菌體生長情況不變。因此,后續(xù)實(shí)驗(yàn)在培養(yǎng)敲除了panC基因的菌株時,基本培養(yǎng)基中添加10-2g/L泛酸。

圖4 B0016-070B 菌株泛酸添加量的優(yōu)化Fig.4 Optimization of pantothenic acid concentration in B0016-070B
2.4 代謝途徑敲除對β-丙氨酸合成的影響
通過搖瓶發(fā)酵實(shí)驗(yàn),比較B0016-060A、B0016-070B和B0016-080BB菌株的β-丙氨酸合成水平。結(jié)果如圖5所示,B0016-050菌株的β-丙氨酸產(chǎn)量為5.2 mg/L,B0016-060A菌株的β-丙氨酸合成量較B0016-050提高了35.1%,說明敲除lysC基因有助于L-天冬氨酸的積累,從而提高β-丙氨酸的產(chǎn)量;B0016-070B菌株的β-丙氨酸合成量較B0016-060A提高了595.8%,表明敲除panC基因阻斷了β-丙氨酸的分解,從而使其合成量大幅度提高;B0016-080BB菌株的β-丙氨酸產(chǎn)量為49.3 mg/L,較B0016-070B提高了1.7%,說明敲除ptsG基因?qū)Ζ?丙氨酸的合成也有一定影響。

圖5 基因敲除對β-丙氨酸合成的影響Fig.5 Effects of gene deletions on β-alanine synthesis
2.5panD基因的異源過表達(dá)對β-丙氨酸合成的影響
將pPL-panD質(zhì)粒分別轉(zhuǎn)化入菌株B0016-050、B0016-060A、B0016-070B、B0016-080BB,獲得B0016-050/pPL-panD、B0016-060A/pPL-panD、B0016-070B/pPL-panD、B0016-080BB/pPL-panD重組株,進(jìn)一步進(jìn)行發(fā)酵實(shí)驗(yàn)檢測其β-丙氨酸合成性能。
如圖6所示,過量表達(dá)panD基因顯著提高了β-丙氨酸合成量,B0016-050/pPL-panD菌株的β-丙氨酸合成量達(dá)2.4 g/L,比B0016-050重組菌株提高了464.1倍;B0016-080BB/pPL-panD重組菌β-丙氨酸合成量最高,達(dá)4.3 g/L,比B0016-080BB重組菌β-丙氨酸合成量提高了86.2倍。敲除lysC、panC和ptsG基因的B0016-060A/pPL-panD、B0016-070B/pPL-panD和B0016-080BB/pPL-panD菌株的β-丙氨酸合成量依次升高12.5%、39.3%和13.3%。可見,敲除天冬氨酸的分解途徑關(guān)鍵基因lysC和β-丙氨酸的分解途徑關(guān)鍵基因panC顯著提高了β-丙氨酸的合成量;此外,也證明了在以甘油為碳源時,敲除ptsG基因同樣可將增加β-丙氨酸代謝流。

圖6 panD基因的過量表達(dá)對不同菌株發(fā)酵合成β-丙氨酸的影響Fig.6 Effect of over expression of panD on the synthesis of β-alanine in different strains
2.6 B0016-080BB/pPL-panD菌株搖瓶發(fā)酵優(yōu)化
2.6.1 誘導(dǎo)時機(jī)的優(yōu)化
ZHOU等優(yōu)化了PL啟動子調(diào)控的發(fā)酵過程,其最佳的菌體生長溫度為33 ℃,而最佳的產(chǎn)物合成溫度為42 ℃[9-10]。本文進(jìn)一步對菌株B0016-080BB/pPL-panDβ-丙氨酸發(fā)酵過程中,這兩個溫度的轉(zhuǎn)折時機(jī)進(jìn)行優(yōu)化,結(jié)果如圖7所示。當(dāng)菌體生長至OD600為2.5時,將發(fā)酵溫度由33 ℃切換為42 ℃,β-丙氨酸合成量最高,達(dá)到4.9 g/L。因此,最佳溫度誘導(dǎo)時機(jī)為OD600值達(dá)到2.5。

圖7 B0016-080BB/pPL-panD菌株溫度誘導(dǎo)時機(jī)的優(yōu)化Fig.7 Optimization of thermo-induction of strain B0016-080BB/pPL-panD
2.6.2 發(fā)酵周期的優(yōu)化
進(jìn)一步對菌株B0016-080BB/pPL-panD的β-丙氨酸發(fā)酵周期進(jìn)行優(yōu)化。測定發(fā)酵液中的β-丙氨酸產(chǎn)量和菌體濃度,結(jié)果如圖8所示。當(dāng)搖瓶發(fā)酵42 h時,β-丙氨酸合成量最高,達(dá)到5.0±0.2 g/L,體積生產(chǎn)強(qiáng)度達(dá)到(0.12±0.005) g/(L·h)。因此,菌株B0016-080BB/pPL-panD的最佳搖瓶發(fā)酵周期為42 h。

圖8 B0016-080BB/pPL-panD菌株搖瓶發(fā)酵時間的優(yōu)化Fig.8 Optimization of fermentation time of strain B0016-080BB/pPL-panD
3 結(jié)論
本文在去除了有機(jī)酸、醇副產(chǎn)物的重組E.coliB0016-050菌株中,刪除β-丙氨酸競爭代謝途徑(lysC)和分解途徑基因(panC),可顯著提高β-丙氨酸的積累量;并證明了在以甘油為碳源時,敲除ptsG基因同樣可提高β-丙氨酸合成量。此外,用溫控型PL強(qiáng)啟動子調(diào)控谷氨酸棒桿菌(Corynebacteriumglutamicum)來源的L-天冬氨酸α-脫羧酶的表達(dá),可顯著提高重組E.coli的β-丙氨酸合成水平。優(yōu)化后B0016-080BB/pPL-panD菌株β-丙氨酸合成量達(dá)到(5.0±0.2) g/L,體積生產(chǎn)強(qiáng)度達(dá)到(0.12±0.005) g/(L·h)。
[1] JANG Y S,KIM B,SHIN J H,et al.Bio-based production of C2-C6 platform chemicals[J].Biotechnology and Bioengineering, 2012,109(10):2 437-2 459.
[2] LEE J W,NA D,PARK J M,et al.Systems metabolic engineering of microorganisms for natural and non-natural chemicals[J].Nature Chemical Biology,2012,8(6):536-546.
[3] STEUNENBERGP,K NST P M,SCOTT E L,et al.Polymerisation of β-alanine through catalytic ester-amide exchange[J].European Polymer Journal,2013,49(49):1 773-1 781.
[4] LIANG Lu-yi, ZHENG Yu-guo, SHEN Yin-chun.Optimization of β-alanine production from β-aminopropionitrile by resting cells ofRhodococcussp. G20 in a bubble column reactor using response surface methodology[J].Process Biochemistry,2008,43(7):758-764.
[5] K NST P M,FRANSSEN M C R,SCOTT E L,et al.A study on the applicability ofL-aspartate α-decarboxylase in the biobased production of nitrogen containing chemicals[J]. Green Chemistry,2009,11(10):1 646-1 652.
[6] BORODINA I,KILDEGAARD K R,JENSEN N B,et al.Establishing a synthetic pathway for high-level production of 3-hydroxypropionic acid inSaccharomycescerevisiaevia β-alanine[J].Metabolic Engineering,2014,27:57-64.
[7] SONG C W,LEE J,KO Y S,et al.Metabolic engineering ofEscherichiacolifor the production of 3-aminopropionic acid.[J].Metabolic Engineering,2015,30(3):121-129.
[8] 呂忠成.Ptsg敲除對重組大腸桿菌碳氮代謝的影響.[D].西安:西北大學(xué),2015.
[9] ZHOU Li,NIU Dan-dan,TIAN Kang-ming,et al.Genetically switchedD-lactate production inEscherichiacoli[J]. Metabolic Engineering,2012,14(5):560-568.
[10] ZHOU Li,DENG Can,CUI Wen-jing,et al.EfficientL-alanine production by a thermo-regulated switch inEscherichiacoli[J].Applied Biochemistry and Biotechnology,2016,178(2):324-337.
[11] 周麗,鄧璨,崔文璟,等.溫度調(diào)節(jié)基因開關(guān)調(diào)控大腸桿菌發(fā)酵合成L-丙氨[J].微生物學(xué)通報,2015,42(11):2 272-2 281.
[12] DATSENKO K A,WANNER B L.One-step inactivation of chromosomal genes inEscherichiacoliK-12 using PCR products[J].Proceedings of the National Academy of Sciences of the United States of America, 2000,97(12): 6 640-6 645.
[13] SONDEREGGER M,JEPPSSON M,HAHN-H GERDAL B,et al.Molecular basis for anaerobic growth ofSaccharomycescerevisiaeon xylose, investigated by global gene expression and metabolic flux analysis[J].Applied & Environmental Microbiology,2004,70(4):2 307-2 317.
[14] 石增秀,崔文璟,周麗,等.谷氨酸棒桿菌L-天冬氨酸α-脫羧酶基因的克隆及重組酶性質(zhì)研究[J].生物技術(shù)通報,2013(4):110-115.
Metabolic engineering ofEscherichiacolifor the production of β-alanine
LIANG Shan-shan, ZHOU Li,ZHANG Bin, ZHOU Zhe-min*
(Schoool of Biotechnology and the Key Laboratory of Industrial Biotechnology, Jiangnan University, Wuxi 214122, China)
β-Alanine, the only kind of β amino acid which exists naturally, has important applications in medicine, food and chemical industry. In this study, the possibility of metabolic engineering ofEscherichiacolifor the production of β-alanine was investigated. TheEscherichiacoliCICIM B0016-050 (ackA-ptapflBadhEfrdAldhA) strain with deletions on synthetic pathways of acetic acid, formic acid, ethanol, succinic acid and lactic acid, was used as the starting strain. Effects of gene knockouts oflysC(encoding aspartokinase),panC(encoding pantothenate synthetase) andptsG(encoding the major glucose transporter IICB (Glc)) and overexpression of theCorynebacteriumglutamicumpanDgene (encodingL-aspartate-α-decarboxylase) on synthesis of β-alanine in B0016-050 were investigated. The synthesis of β-alanine was improved by 12.5%, 39.3% and 13.3%, respectively, after combined gene knockouts, and was increased by more than 100.6 times after overexpressing thepanDgene. After optimization of fermentation conditions, the yield of β-alanine in the shake flask fermentation of strain B0016-080BB/pPL-panDreached 5.0±0.2 g/L, and the volume productivity was 0.12±0.01 g/(L·h). The results laid the foundation for fermentation of β-alanine.
β-alanine;Escherichiacoli;L-aspartate-α-decarboxylase;metabolic engineering;gene knockout
10.13995/j.cnki.11-1802/ts.201705003
碩士研究生(周哲敏教授為通訊作者,E-mail:zhmzhou@jiangnan.edu.cn)。
國家自然科學(xué)基金項(xiàng)目(31300087);基本科研-重點(diǎn)項(xiàng)目(JUSRP51611A)
2017-01-05,改回日期:2017-02-09
猜你喜歡
房地產(chǎn)導(dǎo)刊(2022年5期)2022-06-01 06:20:14
能源工程(2022年1期)2022-03-29 01:06:28
建材發(fā)展導(dǎo)向(2021年12期)2021-07-22 08:06:48
建材發(fā)展導(dǎo)向(2021年7期)2021-07-16 07:07:52
中學(xué)生數(shù)理化(高中版.高二數(shù)學(xué))(2021年12期)2021-04-26 07:43:48
中學(xué)生數(shù)理化(高中版.高考數(shù)學(xué))(2021年12期)2021-03-08 01:28:50
今日農(nóng)業(yè)(2020年16期)2020-12-14 15:04:59
消費(fèi)導(dǎo)刊(2018年8期)2018-05-25 13:20:08
家庭影院技術(shù)(2018年4期)2018-05-09 07:07:41
電子制作(2017年20期)2017-04-26 06:57:45
