幾種抗逆增強(qiáng)劑對(duì)蘋果花與幼果抗寒力的效果
2017-07-15 15:31:42王來平薛曉敏路超聶佩顯王
江蘇農(nóng)業(yè)科學(xué) 2017年9期
王來平++薛曉敏++路超++聶佩顯++王金政

摘要:以7年生天紅2號(hào)/SH40/八棱海棠蘋果為試材,花芽膨大期噴施去離子水(CK)、18 mg/L ABA、200 mg/L甜菜堿、40 mg/L水楊酸、60 mg/L蕓薹素、200 mg/L山梨醇等外源物質(zhì),分別于花蕾期、盛花期、幼果期采集樣品,進(jìn)行0、-2、-4、-6、-8 ℃梯度的低溫脅迫,測(cè)定抗寒相關(guān)生理指標(biāo),研究不同外源物質(zhì)對(duì)蘋果花器官不同發(fā)育階段抗寒力的影響效果。結(jié)果發(fā)現(xiàn),隨著環(huán)境溫度的升高,花器官不同發(fā)育階段半致死溫度升高,抗寒力降低,即抗寒力花蕾期>盛花期>幼果期;施用外源物質(zhì)降低了花器官不同發(fā)育階段半致死溫度,提高了花器官的抗寒力;5種外源物質(zhì)均是通過提高低溫脅迫下花器官可溶性糖含量及抗氧化酶SOD、POD活性,降低了細(xì)胞MDA含量和電解質(zhì)外滲率,從而提高了蘋果花器官抗寒力。本試驗(yàn)表明施用18 mg/L ABA效果最好,其次是40 mg/L水楊酸。
關(guān)鍵詞:抗寒力;蘋果花期;蕓薹素;水楊酸;甜菜堿;山梨醇;ABA
中圖分類號(hào): S661.101文獻(xiàn)標(biāo)志碼: A
文章編號(hào):1002-1302(2017)09-0109-03
蘋果為我國重要的果樹樹種,山東省、陜西省、甘肅省等蘋果主產(chǎn)區(qū)花期受寒流侵襲,易遭受低溫凍害,導(dǎo)致減產(chǎn),給生產(chǎn)帶來巨大損失。目前,普遍采用施放煙霧、花期噴水等傳統(tǒng)方法防止蘋果花期凍害,這些方法耗時(shí)費(fèi)力,收效不大。因此,篩選能夠有效增強(qiáng)蘋果抗寒力的保護(hù)劑,不僅能夠解決生產(chǎn)實(shí)際問題,對(duì)于研究低溫脅迫基礎(chǔ)理論也具有重要意義。蕓薹素、水楊酸、甜菜堿、山梨醇、ABA作為植物抗逆增強(qiáng)劑,在調(diào)節(jié)植物抗旱、耐鹽堿等方面效用明顯,調(diào)節(jié)機(jī)理不盡相同,其施用效果也有所差別。有研究表明,ABA是抗寒基因表達(dá)的啟動(dòng)因子,對(duì)植物抗寒力的調(diào)控起著非常重要的作用[1-3],甜菜堿作為一種相容性物質(zhì)對(duì)植物的抗逆性起著調(diào)節(jié)作用。外施ABA、甜菜堿、水楊酸等能夠有效提高植物的抗寒力,這已在番茄、水稻、芝麻等植物上多見報(bào)道[4-6],但這些物質(zhì)對(duì)提高蘋果花期抗寒力的效果怎樣,有什么樣的差別,還未見報(bào)道。本研究以7年生天紅2號(hào)蘋果為材料,探討了外施蕓薹素、水楊酸、甜菜堿、山梨醇、ABA對(duì)蘋果花期抗寒力及相關(guān)生理指標(biāo)的影響,以期為選擇適宜于蘋果的抗逆增強(qiáng)劑提供參考,并為揭示蘋果花期抗寒生理特性和提高防凍技術(shù)提供理論依據(jù)。
1材料與方法
1.1材料
試驗(yàn)在山東省果樹研究所泰東基地進(jìn)行,試材為天紅2號(hào)/SH40/八棱海棠,7年生,生長良好,長勢(shì)一致,管理水平較高。
1.2試劑與儀器
外源蕓薹素、水楊酸、甜菜堿、山梨醇、ABA由北京鼎國生物技術(shù)有限責(zé)任公司生產(chǎn)。DDS-11C型電導(dǎo)儀由上海光學(xué)儀器廠生產(chǎn)。
1.3方法
于2016年3月中旬花芽膨大期噴施18 mg/L ABA、200 mg/L 甜菜堿、40 mg/L水楊酸、60 mg/L蕓薹素、200 mg/L 山梨醇(為多次試驗(yàn)篩選濃度)。試驗(yàn)按3株每小區(qū),3次重復(fù),隨機(jī)區(qū)組設(shè)計(jì)。噴施后在顯蕾期、盛花期和幼果期分別采集花蕾、花朵和幼果作為試樣,在低溫冰箱中進(jìn)行低溫脅迫處理,設(shè)置0、-2、-4、-6、-8 ℃梯度的脅迫。冰箱放入材料時(shí)溫度為室溫,然后進(jìn)行降溫,降溫速度2 ℃/h,達(dá)到所需處理溫度后維持8 h,之后,切斷電源,冰箱溫度自然回升至室溫,取出材料,將材料室內(nèi)靜置10 h,進(jìn)行各生理指標(biāo)測(cè)定。
采用楊向娜的方法[7]進(jìn)行電解質(zhì)滲出率測(cè)定,電解質(zhì)滲出率=(煮沸前的電導(dǎo)值/煮沸后的電導(dǎo)值)×100%。對(duì)電解質(zhì)滲出率配以Logistic方程y=k/(1+ae-bx),用擬合的 Logistic 方程的拐點(diǎn)溫度表示組織的半致死溫度LT50[8]。式中,y表示電導(dǎo)率,x表示處理溫度,k、a、b為常數(shù),e為自然對(duì)數(shù)。可溶性糖含量用蒽酮比色法測(cè)定[9];丙二醛含量用硫代巴比妥酸TBA顯色法測(cè)定;SOD酶活性用氮蘭四唑(NBT)顯色法測(cè)定;POD酶活性用愈創(chuàng)木酚比色法測(cè)定。
2結(jié)果與分析
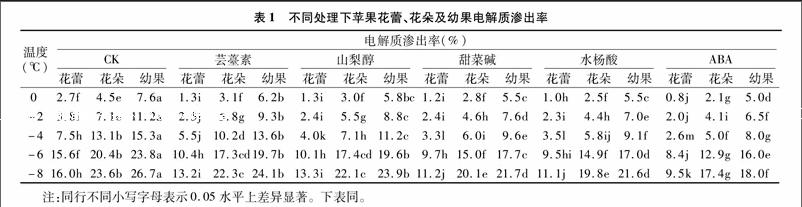
2.1不同處理下蘋果花、幼果電解質(zhì)滲出率
在不同程度低溫脅迫下,蘋果花蕾、花朵和幼果的電解質(zhì)滲出率變化趨勢(shì)一致,均隨溫度的下降而上升,說明隨著低溫脅迫的加劇,低溫對(duì)花器官不同發(fā)育時(shí)期細(xì)胞的傷害逐漸增加(表1)。噴施各調(diào)節(jié)劑后,蘋果花蕾、花朵和幼果電解質(zhì)滲出率均顯著小于對(duì)照,說明各調(diào)節(jié)劑能緩解低溫對(duì)蘋果細(xì)胞造成的傷害。花器官不同發(fā)育階段,低溫下電解質(zhì)滲透率不同,花蕾期<盛花期<幼果期,說明花器官在不同的發(fā)育階段抗寒力花蕾期>盛花期>幼果期。比較各調(diào)節(jié)劑處理下電解質(zhì)滲透率發(fā)現(xiàn)ABA處理的滲透率最低,說明ABA在提高蘋果花器官抗寒力方面效果最好,在0 ℃到-6 ℃的低溫脅迫下水楊酸處理效果優(yōu)于蕓薹素、甜菜堿、山梨醇,在-8 ℃低溫脅迫下水楊酸和甜菜堿效果沒有顯著差別,二者均優(yōu)于山梨醇和蕓薹素。
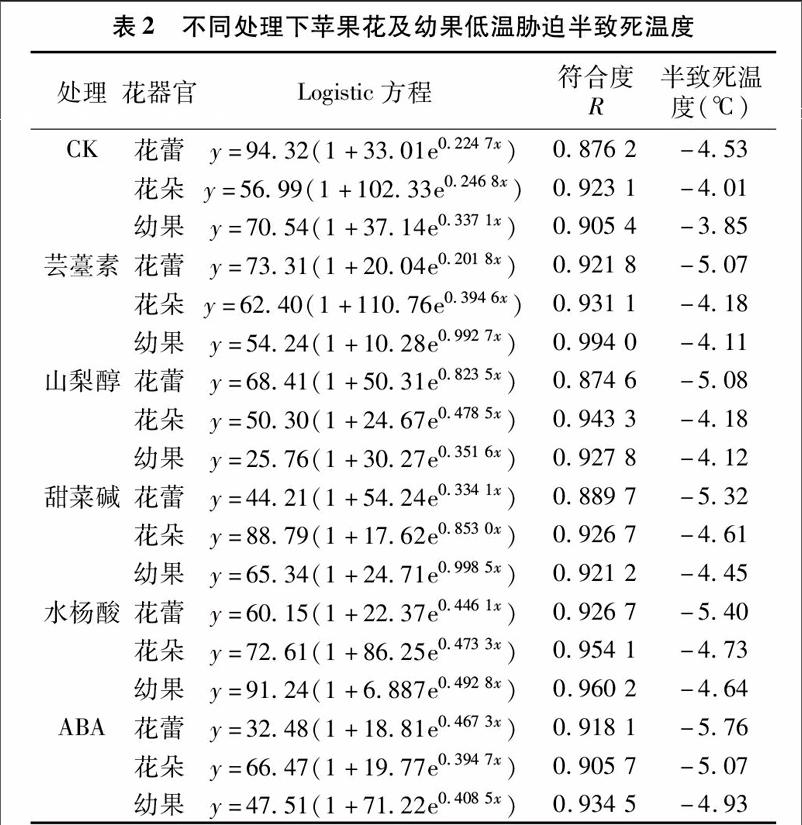
2.2不同處理對(duì)蘋果花、幼果半致死溫度的影響
采用電解質(zhì)滲出率及Logistic回歸方程,求得蘋果花蕾、花朵和幼果的半致死溫度(LT50)(表2)。從表2可以看出,不同處理下花和幼果的半致死溫度大小為花蕾<花朵<幼果,表明蘋果在器官發(fā)育過程中,隨著環(huán)境溫度的上升,器官不同發(fā)育時(shí)期半致死溫度逐漸升高,抗寒性逐漸降低。噴施調(diào)節(jié)劑后,蘋果花蕾、花朵和幼果的半致死溫度較對(duì)照均明顯降低(α=0.05),表明外源蕓薹素、水楊酸、甜菜堿、山梨醇、ABA均能提高蘋果的抗寒力,其中ABA處理施用效果最佳,其次是水楊酸。
2.3不同處理對(duì)蘋果及幼果MDA含量的影響
MDA是衡量逆境條件下植物細(xì)胞膜膜脂過氧化程度的指標(biāo)。在不同程度低溫脅迫下,蘋果花蕾、花朵和幼果的MDA含量變化趨勢(shì)一致,均隨溫度的下降而上升(表3)。說明隨著低溫脅迫的加劇,蘋果花不同發(fā)育階段細(xì)胞膜膜脂過氧化程度增加,細(xì)胞傷害逐漸加重。噴施調(diào)節(jié)劑后,蘋果花蕾、花朵和幼果MDA含量均顯著小于對(duì)照,說明調(diào)節(jié)劑能緩解低溫對(duì)植物細(xì)胞造成的傷害,低溫脅迫下MDA含量依次為花蕾期<盛花期<幼果期。比較各調(diào)節(jié)劑處理下MDA含量,外源ABA處理MDA含量最低,說明施用效果最好。
2.4不同處理對(duì)蘋果花、幼果可溶性糖含量的影響
可溶性糖是植物重要的滲透調(diào)節(jié)物質(zhì),是衡量植物抗逆性強(qiáng)弱的重要指標(biāo)之一。表4表明,在低溫脅迫下,蘋果花蕾、花朵和幼果的可溶性糖含量均隨溫度的下降而上升,說明隨著低溫脅迫的加劇,蘋果滲透調(diào)節(jié)能力逐漸增強(qiáng)。噴施各調(diào)節(jié)劑后,蘋果花蕾、花朵和幼果可溶性糖含量均高于對(duì)照,說明各調(diào)節(jié)劑能增強(qiáng)低溫脅迫下花和幼果滲透調(diào)節(jié)能力。花和幼果低溫下滲透調(diào)節(jié)能力不同,其中花蕾期>盛花期>幼果期。各調(diào)節(jié)劑處理下以外源ABA處理可溶性糖含量最高,說明其施用效果最好。
2.5不同處理對(duì)蘋果花及幼果抗氧化酶活性的影響
逆境條件下,植物抗氧化酶活性增強(qiáng),清除由于逆境而產(chǎn)生的過多的活性氧(ROS),防止活性氧對(duì)細(xì)胞產(chǎn)生傷害。表5和表6表明,在低溫脅迫下,蘋果花蕾、花朵和幼果的SOD和POD活性均隨溫度的下降而呈現(xiàn)先上升后下降的趨勢(shì),說明低程度的低溫脅迫下,蘋果抗氧化酶活性逐漸增強(qiáng),隨著脅迫程度加重,酶蛋白遭到破壞,活性降低。噴施各調(diào)節(jié)劑后,蘋果花蕾、花朵和幼果SOD,POD活性均高于對(duì)照, 說明各調(diào)節(jié)劑均能增強(qiáng)低溫脅迫下蘋果花器官抗氧化酶活性,提高其抗寒力。比較各調(diào)節(jié)劑處理下抗氧化酶活性,施用ABA處理抗氧化酶活性顯著高于其他處理,施用效果最好。在 0 ℃ 到-2 ℃的低溫脅迫下水楊酸處理抗氧化酶SOD、POD活性高于蕓薹素、甜菜堿、山梨醇處理,但是在-2 ℃到 -8 ℃ 的低溫脅迫下,甜菜堿處理的抗氧化酶SOD、POD活性顯著高于水楊酸、蕓薹素和山梨醇。這可能是因?yàn)樘鸩藟A具有維持蛋白結(jié)構(gòu)穩(wěn)定的功能[10],低溫脅迫下能更好維持抗氧化酶功能。
3討論
低溫脅迫下植物細(xì)胞膜的結(jié)構(gòu)遭受破壞,且破壞程度隨脅迫程度的增加而增大。細(xì)胞內(nèi)電解質(zhì)外滲率是衡量細(xì)胞膜結(jié)構(gòu)破壞程度的指標(biāo)之一,比較相同脅迫水平下電解質(zhì)的外滲率,是評(píng)價(jià)植物抗寒力的一種有效手段[11]。用電解質(zhì)滲出率及Logistic方程求得半致死溫(LT50),是鑒定植物抗寒性強(qiáng)弱的常用方法,已被廣泛應(yīng)用[12]。植物處于逆境條件下,細(xì)內(nèi)自由基清除與產(chǎn)生的平衡被打破,活性氧自由基逐漸積累,對(duì)細(xì)胞結(jié)構(gòu)產(chǎn)生傷害。細(xì)胞膜系統(tǒng)首先受傷害,造成膜內(nèi)離子滲出和膜脂的過氧化。而作為膜脂過氧化的中間產(chǎn)物之一的MDA,使生物膜受到的傷害進(jìn)一步加劇[13]。SOD和POD是防御膜脂過氧化作用的重要保護(hù)被,是清除逆境下產(chǎn)生的活性氧自由基的關(guān)鍵酶。提高逆境下植物體內(nèi)清除活性氧自由基的SOD和POD等保護(hù)酶活性,就可以減輕膜脂的過氧化程度,降低活性氧自由基對(duì)細(xì)胞結(jié)構(gòu)的傷害,提高植物的抗逆性[14]。本試驗(yàn)中,施用外源蕓薹素、水楊酸、甜菜堿、山梨醇、ABA降低了蘋果花蕾、花朵和幼果的電解質(zhì)滲出率,SOD和POD酶的活性增強(qiáng),減緩了MDA的積累,增加了可溶性糖的含量,降低了半致死溫度,提高了蘋果的抗寒性。這在番茄、芝麻、水稻、香蕉等植物上的研究結(jié)論基本一致[15-17]。
本研究還發(fā)現(xiàn),外源蕓薹素、水楊酸、甜菜堿、山梨醇、ABA對(duì)于提高蘋果抗寒性效果有所差異,在本試驗(yàn)中,施用外源ABA處理效果最佳,其次是水楊酸。施用甜菜堿處理對(duì)于抗氧化酶保護(hù)有著顯著的作用,顯著優(yōu)于蕓薹素、水楊酸、山梨醇,可能是因?yàn)樘鸩藟A具有維持蛋白結(jié)構(gòu)穩(wěn)定的功能[10],低溫脅迫下能更好地維持抗氧化酶功能。因此,生產(chǎn)中施用調(diào)節(jié)劑可以有選擇地進(jìn)行復(fù)配,以提高施用效果。
參考文獻(xiàn):
[1]彭艷華,黃永秀. 脫落酸應(yīng)答基因的結(jié)構(gòu),表達(dá)調(diào)控及信號(hào)轉(zhuǎn)導(dǎo)[J]. 植物生理學(xué)通訊,1996,32(2):155-158.
[2]謝吉容,向鄧云,梅虎,等. 南方紅豆杉抗寒性的變化與內(nèi)源激素的關(guān)系[J]. 西南師范大學(xué)學(xué)報(bào)(自然科學(xué)版),2002,27(2):231-234.
[3]郭鳳領(lǐng),盧育華,李寶光. 外源ABA對(duì)番茄苗期和開花期抗冷特性的影響[J]. 山東農(nóng)業(yè)大學(xué)學(xué)報(bào)(自然科學(xué)版),2000,31(4):357-362.
[4]黨秋玲,余超,王禎麗. ABA處理種子對(duì)加工番茄幼苗抗寒力及相關(guān)生理指的影響[J]. 石河子大學(xué)學(xué)報(bào)(自然科學(xué)版),2005,23(3):349-351.
[5]曾韶西,王以柔,李美茹,等. 冷鍛煉和ABA誘導(dǎo)水稻幼苗提高抗冷性期間膜保護(hù)系統(tǒng)的變化[J]. 熱帶亞熱帶植物學(xué)報(bào),1994,2(1):44-50.
[6]李亞男,陳大清,胡培麗. ABA和6-BA對(duì)不同溫度條件下芝麻幼苗某些生理指標(biāo)的影響[J]. 長江大學(xué)學(xué)報(bào)(自然科學(xué)版),2005,2(8):67-68,75.
[7]楊向娜,魏安智,楊途熙,等. 仁用杏3個(gè)生理指標(biāo)與抗寒性的關(guān)系研究[J]. 西北林學(xué)院學(xué)報(bào),2006,21(3):30-33.
[8]龔月樺,周永學(xué),樊軍鋒,等. 美國黃松、班克松和油松的抗寒性比較[J]. 應(yīng)用生態(tài)學(xué)報(bào),2006,17(8):1389-1392.
[9]高俊鳳. 植物生理學(xué)實(shí)驗(yàn)技術(shù)[M]. 西安:世界圖書出版公司,2000.
[10]侯彩霞,於新建,李榮,等. 甜菜堿穩(wěn)定PSⅡ放氧中心外周多肽機(jī)理[J]. 中國科學(xué):C輯,1998,28(4):355-361.
[11]聶慶娟,孟朝,梁海永,等. 低溫脅迫對(duì)4種常綠闊葉植物膜脂過氧化及保護(hù)酶活性的影響[J]. 植物研究,2007,27(5):578-581.
[12]趙習(xí)平,劉鐵錚,付雅麗. 杏樹花期霜凍危害及其抗寒性研究進(jìn)展[J]. 江西農(nóng)業(yè)學(xué)報(bào),2007,19(11):33-35.
[13]和紅云,田麗萍,薛琳. 植物抗寒性生理生化研究進(jìn)展[J]. 天津農(nóng)業(yè)科學(xué),2007,13(2):10-13.
[14]王燕凌,廖康,劉君,等. 越冬前低溫鍛煉期間不同品種葡萄枝條中滲透性物質(zhì)和保護(hù)酶活性的變化[J]. 果樹學(xué)報(bào),2006,23(3):375-378.
[15]白潔,蔣衛(wèi)杰,余宏軍,等. 外源ABA、Put和BR對(duì)亞適溫條件下番茄幼苗葉片保護(hù)酶活性的影響[J]. 中國農(nóng)學(xué)通報(bào),2007,23(6):317-320.
[16]詹嘉紅,藍(lán)宗輝. 外源ABA對(duì)低溫脅迫水稻幼苗酯酶同工酶的影響[J]. 生物技術(shù),2000,10(4):19-21.
[17]周玉萍,鄭燕玲,田長恩,等. 脫落酸、多效唑和油菜素內(nèi)酯對(duì)低溫期間香蕉過氧化物酶和電導(dǎo)率的影響[J]. 廣西植物,2002,22(5):444-448.
