米黑毛霉UV-LiCl-6凝乳酶酶學(xué)特性研究
2017-07-25 09:33:12朱建寧張衛(wèi)兵張忠明楊敏文鵬程宋雪梅張丹妮
衛(wèi)生職業(yè)教育 2017年13期
關(guān)鍵詞:研究
朱建寧,張衛(wèi)兵,張忠明,楊敏,文鵬程,宋雪梅,張丹妮
(1.甘肅省食品藥品監(jiān)督管理局,甘肅蘭州730000;2.甘肅農(nóng)業(yè)大學(xué)食品科學(xué)與工程學(xué)院,甘肅蘭州730070)
米黑毛霉UV-LiCl-6凝乳酶酶學(xué)特性研究
朱建寧1,張衛(wèi)兵2*,張忠明2,楊敏2,文鵬程2,宋雪梅2,張丹妮2
(1.甘肅省食品藥品監(jiān)督管理局,甘肅蘭州730000;2.甘肅農(nóng)業(yè)大學(xué)食品科學(xué)與工程學(xué)院,甘肅蘭州730070)
以米黑毛霉凝乳酶為研究對象,研究反應(yīng)溫度、pH值、脫脂乳濃度、金屬離子對凝乳酶活力的影響。結(jié)果表明,米黑毛霉凝乳酶最適反應(yīng)溫度為75℃,在35℃~45℃穩(wěn)定性較好;米黑毛霉凝乳酶的最適pH值為6.0,在pH值為7.0~10.0時較為穩(wěn)定;脫脂乳濃度為20.0%時,相對凝乳酶活力達(dá)到最大值;Mn2+、Mg2+、Na+對凝乳酶的凝乳活力有促進(jìn)作用,K+、Sn2+、Cu2+對凝乳酶的凝乳活力有抑制作用。
凝乳酶;米黑毛霉;酶學(xué)特性
凝乳酶是一種最早在未斷奶的小牛胃中發(fā)現(xiàn)的天門冬氨酸蛋白酶,可專一地切割乳中κ-酪蛋白的Phe105-Met106之間的肽鍵,破壞酪蛋白膠束使牛奶凝結(jié),凝乳酶的凝乳能力及蛋白水解能力使其成為干酪生產(chǎn)中形成質(zhì)構(gòu)和特殊風(fēng)味的關(guān)鍵性酶,被廣泛應(yīng)用于奶酪制作。動物凝乳酶最早被用于干酪生產(chǎn),應(yīng)用也最為廣泛,其產(chǎn)值占總量的70%。據(jù)統(tǒng)計,全世界每年要宰殺5 000萬頭小牛,以獲得凝乳酶,造成全球性小牛短缺,由于原料缺乏,凝乳酶價格昂貴。植物凝乳酶雖然來源廣泛,但其蛋白水解力強(qiáng),并且受時間、地點(diǎn)等限制難以發(fā)展[1]。微生物由于生長周期短,產(chǎn)量大,受氣候、地域、時間限制小,用其生產(chǎn)凝乳酶成本較低、提取方便、經(jīng)濟(jì)效益高,可以節(jié)約動、植物資源,所以微生物凝乳酶成為凝乳酶研究的熱點(diǎn)[2]。
國產(chǎn)凝乳酶的種質(zhì)資源主要包括毛霉類、根霉類和曲霉類[3]。不同來源微生物凝乳酶的酶學(xué)特性差異很大。高維東等[4]對微小毛霉凝乳酶的酶學(xué)性質(zhì)進(jìn)行研究,結(jié)果表明酶的最適溫度為60℃,最適pH值為5.5,在65℃保溫5min活力損失95%;鈣離子是酶的激活劑,其含量與酶活力正相關(guān),而鉀離子則對酶有抑制作用,氯化鈉對酶活力的影響不大。潘道東等[5]研究表明,根霉凝乳酶最適pH為5.7,在pH值為5.0~7.5時酶活力保持穩(wěn)定;最適反應(yīng)溫度45℃,60℃保持20min,則完全失活。
本試驗(yàn)以米黑毛霉UV-LiCl-6凝乳酶為對象,研究反應(yīng)溫度、pH值、脫脂乳濃度、金屬離子對凝乳酶活力的影響,以期為凝乳酶的研究與應(yīng)用提供參考。
1 材料與方法
1.1 材料與儀器
1.1.1 試驗(yàn)材料米黑毛霉菌株:菌株為米黑毛霉UV-LiCl-6,由本實(shí)驗(yàn)室選育并保藏。脫脂乳粉:黑龍江完達(dá)山乳業(yè)股份有限公司。麩皮,馬鈴薯。所用試劑均為市售國產(chǎn)分析純或化學(xué)純。
活化培養(yǎng)基:馬鈴薯20%,葡萄糖2%,瓊脂2%,pH值自然。基礎(chǔ)發(fā)酵培養(yǎng)基:將麩皮與水按1∶0.8(w/v)的比例混合,攪拌均勻,在121℃、0.15 MPa下滅菌20min。發(fā)酵培養(yǎng)基:將麩皮與營養(yǎng)液按1∶0.8(w/v)的比例混合,攪拌均勻,在121℃、0.15MPa下滅菌20min。
營養(yǎng)液:葡萄糖2.5 g,乳清粉2.0 g,硝酸銨1.0 g,以蒸餾水溶解后,鹽酸調(diào)pH值至6.0,定容到100m l。
1.1.2 實(shí)驗(yàn)設(shè)備HWS26型電熱恒溫水浴鍋,上海一恒科技有限公司;AL204電子天平,梅特勒—托利多儀器上海有限公司;GZX-GF101-Ⅱ電熱恒溫鼓風(fēng)干燥箱,上海躍進(jìn)醫(yī)療器械有限公司;NRY-200恒溫?fù)u床,上海南榮實(shí)驗(yàn)室設(shè)備有限公司;TGL-20M高速臺式冷凍離心機(jī),長沙湘儀離心機(jī)儀器有限公司;SW-CJ-ZFD型雙人單面凈化工作臺,蘇州凈化設(shè)備有限公司。
1.2 試驗(yàn)方法
1.2.1 菌株的活化將米黑毛霉菌株UV-LiCl-6接種于斜面培養(yǎng)基進(jìn)行活化,32℃培養(yǎng)4~5 d,待長滿黑色孢子后備用。
1.2.2 孢子懸浮液的制備向生長良好的試管斜面種子中加入10m l無菌生理鹽水,刮下孢子,移入裝有玻璃珠和無菌生理鹽水的三角瓶中振蕩,在搖床上振蕩打散,再用帶脫脂棉的漏斗進(jìn)行過濾,調(diào)整孢子數(shù)為106個/m l。
1.2.3 粗酶液的制備將1 m l米黑毛霉種子液接入發(fā)酵培養(yǎng)基,32℃恒溫培養(yǎng)72 h。培養(yǎng)完成后,加入100 m l蒸餾水,20℃、180 r/min振蕩浸提1 h,浸提液用4層紗布過濾,濾液以10 000 r/min離心20min,上清液即為粗酶液。
1.2.4 酶活力的測定凝乳酶活力測定采用Arima等[6]的方法,用0.01 mol/L氯化鈣液配制10%脫脂奶粉液,配制后在室溫放置40min以上使用,當(dāng)天使用,不宜冰箱放置。取5 m l 10%脫脂奶粉液在35℃保溫10 min,加0.5 m l適當(dāng)稀釋的酶液(35℃保溫),立即搖勻,開始計時(凝乳時間控制在40~90 s),并把試管傾斜45°以上。沿試管周旋轉(zhuǎn),以試管壁出現(xiàn)小顆粒為終點(diǎn),記錄凝乳時間。在上述條件下,將40min凝1 m l 10%脫脂奶粉的酶量定義為一個Soxhlet單位(SU)。

公式中,D為酶液稀釋倍數(shù),t為反應(yīng)時間。
1.2.5 反應(yīng)溫度對凝乳酶活力的影響將作用底物脫脂乳溶液分別在30℃、35℃、40℃、45℃、50℃、55℃、60℃、65℃、70℃、75℃、80℃、85℃下保溫5min,把適當(dāng)稀釋的凝乳酶在35℃下保溫5min待用,分別在相應(yīng)溫度下加入0.5m l酶液測定凝乳酶活力,獲得最高凝乳酶活力為100%(對照),計算相對凝乳酶活力[7]。
1.2.6 凝乳酶的熱穩(wěn)定性將酶液分別在35℃、45℃、55℃、65℃、70℃下保溫0、10、20、30、40、50、60min后,測定殘余凝乳酶活力,以未保溫酶液的凝乳酶活力為100%(對照),計算相對凝乳酶活力。
1.2.7 pH值對凝乳酶活力的影響用0.1 mol/L HCl或0.1 mol/LNaOH將脫脂乳和酶液pH值調(diào)至5.5、6.0、6.5、7.0,測定不同反應(yīng)體系下的凝乳酶活力,以獲得的最高凝乳酶活力為100%(對照)。
1.2.8 凝乳酶的pH穩(wěn)定性分別用0.1 mol/L的甘氨酸-鹽酸緩沖液(pH值2.5~4.0)、檸檬酸-磷酸鹽緩沖液(pH值4.5~ 5.5)、磷酸鈉緩沖液(pH值6.0~8.5)、碳酸鹽緩沖液(pH值9.0~11.0)將酶液的pH值分別調(diào)節(jié)到3.5、4.0、4.5、5.0、5.5、6.0、6.5、7.0、7.5、8.0、8.5、9.0、9.5、10.0、10.5、11.0,在室溫下放置24 h,再將pH值調(diào)到5.5,測定殘余的凝乳酶活力,以獲得最高凝乳酶活力為100%(對照),計算相對凝乳酶活力。
1.2.9 脫脂乳濃度對凝乳酶活力的影響分別調(diào)整脫脂乳的濃度為2.5、5.0、7.5、10.0、12.5、15.0、17.5、20.0 g/100 ml,測定酶液在不同濃度脫脂乳液中的凝乳活力,以獲得的最高凝乳酶活力為100%(對照),計算相對凝乳酶活力。
1.2.10 金屬離子對凝乳酶活力的影響在脫脂乳中分別加入MnSO4、MgSO4、CuSO4、KCl、NaCl、SnCl2,使其終濃度為1mmol/L、5mmol/L、10mmol/L,加酶后在室溫下放置40min,測定凝乳酶活力,以未添加金屬離子酶液的凝乳酶活力為100%(對照),計算相對凝乳酶活力。
2 結(jié)果與分析
2.1 反應(yīng)溫度對凝乳酶活力的影響(見圖1)

圖1 反應(yīng)溫度對凝乳酶活力的影響
從圖1可以看出,隨著溫度的升高,相對凝乳酶活力逐漸增加,溫度達(dá)75℃時,相對凝乳酶活力達(dá)到最高值837.20%,隨著溫度的進(jìn)一步升高,相對凝乳酶活力急劇下降,在80℃時降為91.98%,在85℃時,酶蛋白完全變性,已無凝乳酶活力。所以,凝乳酶的最適溫度是75℃,隨著溫度的升高,活力逐漸減低直至失活。
2.2 凝乳酶的熱穩(wěn)定性(見圖2)

圖2 凝乳酶的熱穩(wěn)定性
從圖2可以看出,在55℃時,隨著保溫時間的延長,相對凝乳酶活力逐漸增大,當(dāng)保溫30min時,相對凝乳酶活力達(dá)到最大值218.22%,隨著保溫時間的延長,相對凝乳酶活力逐漸下降,保溫60 min時相對凝乳酶活力最低;在65℃時,隨著保溫時間的延長,相對凝乳酶活力逐漸增大,當(dāng)保溫10min時,相對凝乳酶活力達(dá)到最大值181.41%,之后隨著保溫時間的延長,相對凝乳酶活力逐漸下降,保溫60 min時降到最低;在70℃時,隨著保溫時間的延長,相對凝乳酶活力持續(xù)下降,保溫20 min時,凝乳酶完全失活;在35℃~45℃,隨著保溫時間的延長,相對凝乳酶活力緩慢增大且較穩(wěn)定。綜上所述,凝乳酶在35℃~45℃穩(wěn)定性較好。
2.3 pH值對凝乳酶活力的影響(見圖3)

圖3 pH值對凝乳酶活力的影響
由圖3可以看出,隨著pH值的增大,相對凝乳酶活力先增大,pH值為6.0時,相對凝乳酶活力達(dá)最大值100%。之后,隨著pH值的增大,相對凝乳酶活力下降。有研究顯示,酸性環(huán)境對凝乳酶起促進(jìn)作用的原因是pH值降低,導(dǎo)致酪蛋白疏水性增加,膠束易發(fā)生解離,從而促進(jìn)酪蛋白聚集產(chǎn)生凝乳[8]。由上可知,凝乳酶的最適pH值為6.0。
2.4 凝乳酶的pH穩(wěn)定性(見圖4)
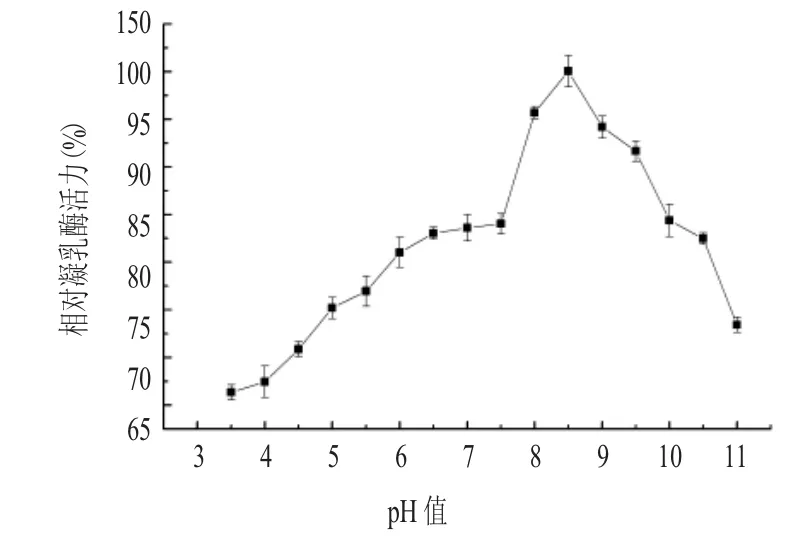
圖4 不同pH值下凝乳酶的穩(wěn)定性
由圖4可以看出,隨著pH值的增大,相對凝乳酶活力逐漸增大,當(dāng)pH值為8.5時,相對凝乳酶活力達(dá)到最大值100.04%,之后隨著pH值的增大,相對凝乳酶活力逐漸下降。在pH值為7.0~10.0時,凝乳活力能保持在80%以上。
2.5 脫脂乳濃度對凝乳酶活力的影響(見圖5)
由圖5可以看出,隨著脫脂乳濃度的增加,凝乳酶活力迅速增大,當(dāng)脫脂乳濃度達(dá)10.0%時,相對凝乳酶活力達(dá)99.17%,之后趨于穩(wěn)定,只有小幅度增加,脫脂乳濃度為20.0%時,凝乳酶活力達(dá)到最大值,為100%。
2.6 金屬離子對凝乳酶活性的影響(見圖6)
由圖6可知,一定濃度的Mn2+、Mg2+、Na+對凝乳酶的凝乳活力有促進(jìn)作用,其中Na+對凝乳酶的凝乳活力有顯著的促進(jìn)作用,K+、Sn2+、Cu2+對凝乳酶的凝乳活力有抑制作用。高巖等[9]研究表明,Mn2+、Mg2+、Ca2+對該酶的凝乳活力有明顯的促進(jìn)作用,而Zn2+、Cu2+對其凝乳活力有抑制作用。
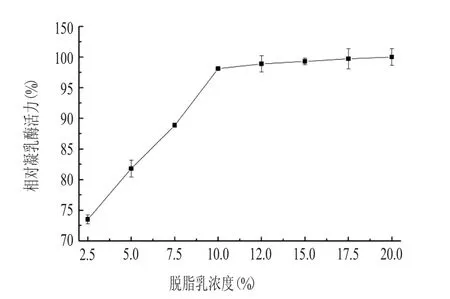
圖5 脫脂乳濃度對凝乳酶活性的影響
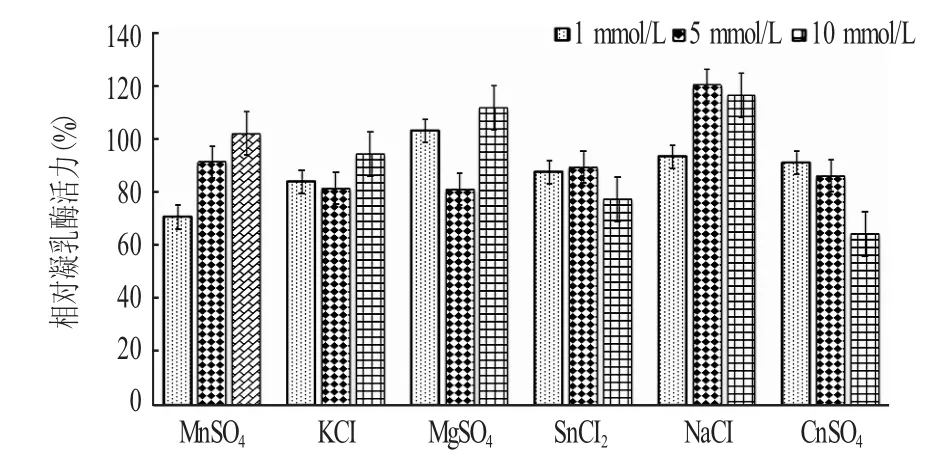
圖6 金屬離子對凝乳酶活性的影響
3 結(jié)論
研究表明,米黑毛霉凝乳酶最適反應(yīng)溫度為75℃;在35℃~45℃時凝乳酶熱穩(wěn)定性較好,70℃時,隨著保溫時間的延長,相對凝乳酶活力逐漸下降,到20 min時完全失活;米黑毛霉凝乳酶的最適pH值為6.0,在pH值為7.0~10.0時,凝乳酶較穩(wěn)定;脫脂乳濃度為20.0%時,凝乳酶活力達(dá)到最大值;Mn2+、Mg2+、Na+對凝乳酶的凝乳活力有促進(jìn)作用,其中Na+對凝乳酶的凝乳活力有顯著的促進(jìn)作用,K+、Sn2+、Cu2+對凝乳酶的凝乳活力有抑制作用。
[1]周俊清,林親錄.微生物源凝乳酶的研究進(jìn)展[J].中國食品添加劑,2004,2(4):6-9.
[2]姜峰,張?zhí)m威.我國凝乳酶特性及其替代品的研究現(xiàn)狀[J].食品研究與開發(fā),2003,24(6):3-6.
[3]甘伯中,高維東,丁福軍,等.微生物凝乳酶固體發(fā)酵條件的研究[J].食品工業(yè)科技,2008,29(4):196-198.
[4]高維東,甘伯中,丁福軍,等.微小毛霉凝乳酶的酶學(xué)性質(zhì)研究[J].食品科學(xué),2010,31(3):185-188.
[5]潘道東,韓玲玲.根霉凝乳酶的分離純化及其酶學(xué)特性研究[J].中國食品學(xué)報,2011,11(2):53-58.
[6]Arima K,Shiniter I,Gakuzo T.Milk-clott ing enzymes from microorganism.part I:screening test and identification of potent fungus[J].Agricultural and Biologi cal Chemistry,1967,31(5):540-545.
[7]錢世鈞,張純青.微小毛霉凝乳酶的純化和性質(zhì)研究[J].微生物學(xué)報,1989,29(4):272-277.
[8]薛璐,陳歷俊,姜鐵民,等.江米酒凝乳酶酶學(xué)特性的研究[J].食品科學(xué),2008,29(4):259-262.
[9]高巖,王景會,李玉秋,等.枯草芽孢桿菌凝乳酶的酶學(xué)性質(zhì)[J].吉林農(nóng)業(yè)大學(xué)學(xué)報,2012,34(4):385-390.
(*通訊作者:張衛(wèi)兵)
G424.31
B
1671-1246(2017)13-0105-03
注:本文系甘肅省自然基金(1606RJZA079);國家自然科學(xué)基金項(xiàng)目(31560442);甘肅農(nóng)業(yè)大學(xué)青年導(dǎo)師扶持基金(GAU-QNDS-201502)
猜你喜歡
體育科技文獻(xiàn)通報(2022年3期)2022-05-23 13:46:54
天津外國語大學(xué)學(xué)報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機(jī)設(shè)計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學(xué)報(2017年2期)2017-07-05 08:13:02
國際商務(wù)財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
