貝母素甲對(duì)多藥耐藥人白血病細(xì)胞活力和凋亡的影響及機(jī)制
2017-08-09 01:38:38齊彥廖斌徐成波皇甫真萍陳佳薇福建中醫(yī)藥大學(xué)附屬人民醫(yī)院福州350004
山東醫(yī)藥 2017年26期
齊彥,廖斌,徐成波,皇甫真萍,陳佳薇(福建中醫(yī)藥大學(xué)附屬人民醫(yī)院,福州350004)
?
貝母素甲對(duì)多藥耐藥人白血病細(xì)胞活力和凋亡的影響及機(jī)制
齊彥,廖斌,徐成波,皇甫真萍,陳佳薇
(福建中醫(yī)藥大學(xué)附屬人民醫(yī)院,福州350004)
目的 觀察貝母素甲對(duì)多藥耐藥白血病人細(xì)胞K562/A02活力與細(xì)胞凋亡的影響,探討活性氧(ROS)在此過程中的作用。方法 將K562/A02細(xì)胞成3組,藥物組分別加入終濃度為100、200、400 μmol/L的貝母素甲;NAC預(yù)處理組采用5 mmlo/L ROS清除劑N-乙酰-L-半胱氨酸(NAC)預(yù)處理后,再加入終濃度為400 μmol/L的貝母素甲;對(duì)照組加等體積的藥物溶解介質(zhì)RPMI1640培養(yǎng)基。采用MTT法檢測各組細(xì)胞活力,流式細(xì)胞術(shù)檢測細(xì)胞凋亡情況,紫外分光光度法檢測細(xì)胞內(nèi)ROS、GSH水平。結(jié)果 與對(duì)照組比較,藥物組細(xì)胞活力均降低(P<0.05或<0.01)、細(xì)胞內(nèi)ROS水平增高(P均<0.01)、GSH水平下降(P均<0.01)、細(xì)胞凋亡率上升(P均<0.05),NAC預(yù)處理組的細(xì)胞活力、細(xì)胞凋亡率和ROS水平無明顯變化(P均>0.05)。結(jié)論 貝母素甲可抑制多藥耐藥人白血病細(xì)胞K562/A02的細(xì)胞活力、誘導(dǎo)其凋亡;其機(jī)制是通過誘導(dǎo)細(xì)胞ROS爆發(fā),引起GSH水平下降,從而逆轉(zhuǎn)多藥耐藥的發(fā)生。
貝母素甲;慢性粒細(xì)胞白血病;K562/A02;多藥耐藥;活性氧;細(xì)胞活力;細(xì)胞凋亡
慢性粒細(xì)胞白血病(CML)是由Bcr-Abl酪氨酸激酶持續(xù)激活引起的血液系統(tǒng)惡性腫瘤。腫瘤細(xì)胞的多藥耐藥現(xiàn)象(MDR)是導(dǎo)致CML化療失敗主要原因之一[1]。浙貝母為百合科植物浙貝母的干燥鱗莖,具有清熱化痰、散結(jié)解毒的功效[2]。現(xiàn)代藥理研究顯示,浙貝母的主要成分貝母素甲可抑制多種腫瘤細(xì)胞生長,逆轉(zhuǎn)腫瘤細(xì)胞的MDR[3~5],但具體作用機(jī)制的研究不多。活性氧(ROS)是細(xì)胞內(nèi)一類重要信號(hào)分子,參與細(xì)胞增殖、凋亡、多藥耐藥產(chǎn)生等諸多事件[6,7]。一些化療藥物如Chaetominine、小白菊內(nèi)酯、小檗胺等能調(diào)節(jié)多藥耐藥CML細(xì)胞內(nèi)的ROS水平,誘導(dǎo)細(xì)胞凋亡,抑制細(xì)胞增殖[8~10]。2015~2016年,我們采用多藥耐藥CML細(xì)胞株K562/A02細(xì)胞[11]進(jìn)行體外研究,觀察貝母素甲對(duì)多藥耐藥CML細(xì)胞體外增殖和凋亡的影響,探討ROS的變化及其機(jī)制,為貝母素甲治療CML耐藥提供實(shí)驗(yàn)依據(jù)。
1 材料與方法
1.1 材料 人CML多藥耐藥細(xì)胞株K562/A02由福建省血液病研究所惠贈(zèng)。RPMI1640培養(yǎng)基和胎牛血清(FBS)購自美國Gibco公司;貝母素甲(純度>98%)和鹽酸阿霉素購自美國阿拉丁工業(yè)公司;MTT和二甲基亞砜(DMSO)購自美國Amresco公司;ROS清除劑N-乙酰-L-半胱氨酸(NAC)、Annexin V-FITC細(xì)胞凋亡檢測試劑盒,ROS檢測試劑盒和GSH檢測試劑盒購自碧云天生物技術(shù)研究所。細(xì)胞培養(yǎng)瓶、培養(yǎng)板購自美國Corning公司;IX70倒置相差顯微鏡及數(shù)碼攝像裝置產(chǎn)自日本Olympus公司;二氧化碳培養(yǎng)箱購自德國Heraeus公司,超凈工作臺(tái)為蘇州凈化設(shè)備有限公司產(chǎn)品,ELX800全自動(dòng)酶標(biāo)儀產(chǎn)自BioTek公司,全波長多功能微孔板檢測儀Infinite 200 PRO產(chǎn)自瑞士TECAN公司。
1.2 細(xì)胞培養(yǎng)與分組 取K562/A02接種于含10% FBS的RPMI1640培養(yǎng)基,置于37 ℃、飽和濕度、5% CO2培養(yǎng)箱中培養(yǎng),2~3 d傳代1次。培養(yǎng)基中加入鹽酸阿霉素1 μg/mL維持細(xì)胞的耐藥性。實(shí)驗(yàn)前1周更換不含鹽酸阿霉素的培養(yǎng)基,取對(duì)數(shù)生長期細(xì)胞用于實(shí)驗(yàn)。將細(xì)胞分為藥物組、對(duì)照組和NAC預(yù)處理組,各藥物組分別加入終濃度為100、200、400 μmol/L的貝母素甲;對(duì)照組加等體積的藥物溶解介質(zhì)于RPMI1640培養(yǎng)基;NAC預(yù)處理組先加入5 mmol/LNAC預(yù)處理1 h,再加入終濃度為400 μmol/L的貝母素甲。
1.3 細(xì)胞活力觀察 取對(duì)數(shù)生長期細(xì)胞,按4×104/mL接種于96孔板,每孔細(xì)胞懸液體積為100 μL。常規(guī)培養(yǎng)過夜后,按照1.2方法進(jìn)行分組處理,每組設(shè)6個(gè)平行孔。培養(yǎng)24 h,用常規(guī)MTT法檢測。每孔分別加入5 mg/mL MTT溶液15 μL,培養(yǎng)3 h,離心棄上清,加入DMSO振蕩溶解沉淀。選擇570 mn為主波長,630 nm為參比波長,在酶標(biāo)儀上測定各孔的光吸收值(A值)。實(shí)驗(yàn)重復(fù)3次。
1.4 細(xì)胞凋亡觀察 按照1.2方法將細(xì)胞分組處理,常規(guī)培養(yǎng)24 h。離心收集細(xì)胞,用預(yù)冷的PBS洗滌,取適量細(xì)胞加入195 μL Annexin V-FITC結(jié)合液,再加入5 μL Annexin V-FITC和10 μL碘化丙啶染色液,室溫避光孵育20 min,上流式細(xì)胞儀檢測。每份樣品檢測104個(gè)細(xì)胞,計(jì)數(shù)處于細(xì)胞周期早期、晚期凋亡的細(xì)胞數(shù),計(jì)算早期調(diào)亡率、晚期凋亡率。雙參數(shù)結(jié)果以二維散點(diǎn)圖顯示,數(shù)據(jù)由FCM計(jì)算機(jī)軟件分析輸出。實(shí)驗(yàn)重復(fù)3次。
1.5 細(xì)胞內(nèi)ROS檢測 藥物組加入終濃度為200 μmol/L的貝母素甲后,分別于培養(yǎng)0、3、6、9、12、24 h時(shí)檢測細(xì)胞內(nèi)ROS表達(dá)。取ROS表達(dá)最高的時(shí)點(diǎn),按1.2方法將細(xì)胞分組處理,檢測各組ROS表達(dá)。各組細(xì)胞分別加入10 μmol/L DCFH-CA檢測探針,于37 ℃細(xì)胞培養(yǎng)箱中避光孵育30 min,離心收集細(xì)胞,用無血清細(xì)胞培養(yǎng)液洗滌3次。選擇488 nm為激發(fā)波長,525 nm為發(fā)射波長,于全波長多功能微孔板檢測儀檢測。ROS表達(dá)以相對(duì)于對(duì)照組的倍數(shù)(fold)表示。實(shí)驗(yàn)重復(fù)3次。
1.6 細(xì)胞內(nèi)GSH檢測 按照1.2方法將細(xì)胞分為藥物組和對(duì)照組,常規(guī)培養(yǎng)24 h,計(jì)數(shù)并收集細(xì)胞,用PBS洗滌1次,加入細(xì)胞沉淀體積3倍量的蛋白去除試劑溶液,充分振蕩;利用液氮和37 ℃溫水快速凍融2次,4 ℃離心10 min,取上清。取適量待測上清液加入檢測工作液,25 ℃孵育45 min。加入0.5 mg/mL的NADPH 50 μL,于405 nm處檢測光吸收值,根據(jù)標(biāo)準(zhǔn)曲線計(jì)算總GSH含量。同時(shí)取部分上述準(zhǔn)備好的待測上清液,加入5%體積的GSH清除輔助液和1%體積的GSH清除工作液,25 ℃反應(yīng)60 min清除GSH,然后加入檢測工作液,參照總GSH的測定方法,檢測細(xì)胞內(nèi)氧化型谷胱苷肽(GSSG)含量。將測定得到的總GSH含量減去GSSG含量,計(jì)算得出細(xì)胞中GSH含量。實(shí)驗(yàn)重復(fù)3次。
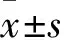
2 結(jié)果
2.1 各組細(xì)胞活力比較 藥物組加入100、200、400 μmol/L貝母素甲后,細(xì)胞活力A值分別為0.392±0.040、0.314±0.022、0.161±0.033,對(duì)照組和NAC預(yù)處理組細(xì)胞活力A值分別為0.462±0.012、0.497±0.028。藥物組細(xì)胞活力較對(duì)照組均降低(P均<0.05或<0.01),NAC預(yù)處理組細(xì)胞活力與對(duì)照組比較無統(tǒng)計(jì)學(xué)差異(P>0.05)。
2.2 各組細(xì)胞凋亡率比較 見表1。藥物組加入100、200、400 μmol/L貝母素甲后細(xì)胞凋亡率較對(duì)照組上升(P均<0.05),NAC預(yù)處理組細(xì)胞凋亡率與對(duì)照組比較無統(tǒng)計(jì)學(xué)差異(P>0.05)。

表1 各組細(xì)胞凋亡率比較
注:與對(duì)照組比較,*P<0.05。
2.3 各組細(xì)胞ROS水平比較 藥物組加入200 μmol/L貝母素甲處理0、3、6、9、12、24 h時(shí)的細(xì)胞ROS水平分別為對(duì)照組的1.00、1.10±0.09、1.34±0.06、1.56±0.06、1.21±0.08、1.04±0.06倍,培養(yǎng)3 h時(shí)ROS水平開始上升,9 h時(shí)達(dá)到峰值,然后隨時(shí)間延長開始下降,24 h時(shí)與對(duì)照組無統(tǒng)計(jì)學(xué)差異。采用100、200、400 μmol/L貝母素甲處理細(xì)胞9 h,細(xì)胞ROS水平分別為對(duì)照組的1.22±0.01、1.64±0.09、1.91±0.14倍,均高于對(duì)照組(P均<0.01);NAC預(yù)處理組培養(yǎng)9 h時(shí)細(xì)胞ROS水平為對(duì)照組的1.06±0.08倍,與對(duì)照組比較無統(tǒng)計(jì)學(xué)差異(P>0.05)。
2.4 各組細(xì)胞GSH水平比較 藥物組加入100、200、400 μmol/L貝母素甲后細(xì)胞GSH水平分別為(4.44±0.17)、(3.87±0.31)、(3.06±0.14)μmol/L,與對(duì)照組的(5.68±0.06)μmol/L比較,P均<0.01。
3 討論
CML是一組由多潛能造血干細(xì)胞異常增殖引起的惡性骨髓性疾病,發(fā)病率為1/10萬~1.5/10萬[12]。雖然以伊馬替尼為代表的酪氨酸激酶抑制劑已成功用于CML的一線治療,大幅度提高了患者的存活率[13],但仍有眾多患者在治療中產(chǎn)生對(duì)化療藥物的MDR,導(dǎo)致化療失敗。逆轉(zhuǎn)MDR是CML治療的重點(diǎn)之一[3]。產(chǎn)生MDR的機(jī)制很多,除了P-糖蛋白(P-gp)過表達(dá)及其對(duì)底物的外排作用之外,ROS信號(hào)亦參與其中[14]。在阿霉素誘導(dǎo)的K562多藥耐藥性細(xì)胞中,GSH等抗氧化系統(tǒng)活性的上升和ROS生成的抑制是細(xì)胞產(chǎn)生耐藥性的關(guān)鍵因素之一[7]。
中藥貝母首載于《本草正義》,其性寒,味大苦,具有清熱化痰、散結(jié)解毒的功效,是臨床常用的化痰藥物。近年研究發(fā)現(xiàn),貝母屬植物提取物對(duì)炎癥、高血壓、腫瘤等多種疾病具有治療潛力[15]。貝母素甲是中藥浙貝母的主要成分之一,具有抗腫瘤活性[3~5,16,17]。胡凱文等[5]率先利用MTT法檢測到貝母素甲對(duì)多藥耐藥CML細(xì)胞K562/A02具有體外細(xì)胞毒作用,但具體作用機(jī)制未明。因此,研究貝母素甲對(duì)多藥耐藥CML細(xì)胞增殖抑制的作用機(jī)制具有重要意義。
本研究顯示貝母素甲可顯著抑制K562/A02細(xì)胞活力,引起細(xì)胞內(nèi)ROS上升,GSH含量下降。貝母素甲對(duì)K562/A02細(xì)胞ROS的誘導(dǎo)作用具有劑量與時(shí)間依賴性,可被ROS清除劑NAC所抑制。而NAC預(yù)處理幾乎完全消除了貝母素甲對(duì)K562/A02細(xì)胞活力的抑制和細(xì)胞凋亡的誘導(dǎo)。
許多研究表明,大量的ROS可以和細(xì)胞內(nèi)脂質(zhì)、蛋白質(zhì)、DNA等分子相互作用,觸發(fā)相關(guān)信號(hào)通路,導(dǎo)致細(xì)胞氧化損傷乃至死亡。細(xì)胞內(nèi)的抗氧化防護(hù)系統(tǒng)(如GSH)能清除過多ROS,防護(hù)ROS造成的氧化損傷。細(xì)胞中GSH含量下降常伴隨著細(xì)胞凋亡率上升[18]。一些藥物可通過引起K562細(xì)胞及其多藥耐藥細(xì)胞中ROS的爆發(fā)和GSH含量的下降,進(jìn)而抑制細(xì)胞活力、誘導(dǎo)細(xì)胞凋亡,殺傷白血病細(xì)胞[1,19]。鄭智等[20]研究表明,復(fù)方浙貝母顆粒協(xié)同阿霉素可有效降低K562/A02移植瘤組織細(xì)胞中GSH含量,抑制抗氧化酶谷胱甘肽-S-轉(zhuǎn)移酶的表達(dá),提高難治耐藥性白血病臨床緩解率。因此,ROS爆發(fā)與GSH含量下降導(dǎo)致的氧化-還原平衡破壞可能是貝母素甲抑制K562/A02細(xì)胞活力、誘導(dǎo)細(xì)胞凋亡的重要機(jī)制,ROS在這一過程中扮演著重要角色。
細(xì)胞凋亡常常是細(xì)胞ROS爆發(fā)的一個(gè)后繼事件。本研究發(fā)現(xiàn),貝母素甲處理在3 h時(shí)開始誘導(dǎo)ROS生成增加,9 h左右達(dá)到峰值,隨后開始下降并恢復(fù)至與對(duì)照組相當(dāng)?shù)乃剑砻髫惸杆丶卓蓜┝恳蕾囆缘卣T導(dǎo)K562/A02細(xì)胞凋亡,提示貝母素甲誘導(dǎo)的ROS升高是一個(gè)相對(duì)短期的過程,在對(duì)細(xì)胞產(chǎn)生氧化損傷的同時(shí),可能具有信號(hào)效應(yīng),能激活相應(yīng)的信號(hào)分子而引起細(xì)胞凋亡。隨著細(xì)胞凋亡數(shù)量增多,特別是晚期凋亡的細(xì)胞因功能受損導(dǎo)致ROS生成能力下降,藥物引起的ROS產(chǎn)生隨之減少。這亦從一個(gè)側(cè)面提示貝母素甲刺激細(xì)胞產(chǎn)生ROS進(jìn)而誘導(dǎo)細(xì)胞凋亡可能是其抗腫瘤機(jī)制之一。Wang等研究發(fā)現(xiàn),平貝母和川貝母中提取的生物堿可激活Caspase-3,激活Caspase依賴性凋亡[15,21,22]。有報(bào)道表明刺激K562細(xì)胞ROS的產(chǎn)生可促進(jìn)Caspase-3表達(dá),誘導(dǎo)細(xì)胞凋亡[23,24]。因此,貝母素甲在刺激K562/A02細(xì)胞產(chǎn)生ROS、引起細(xì)胞凋亡的過程中是否激活了Caspase-3相關(guān)通路有待于后續(xù)研究。
綜上所述,貝母素甲可通過抑制細(xì)胞活力、誘導(dǎo)細(xì)胞凋亡來抑制多藥耐藥白血病細(xì)胞K562/A02的生長。在這一過程中,貝母素甲誘導(dǎo)細(xì)胞ROS爆發(fā),引起GSH含量下降。ROS清除劑NAC預(yù)處理可消除貝母素甲對(duì)K562/A02生長的抑制作用,提示ROS在貝母素甲抑制多藥耐藥白血病細(xì)胞K562/A02生長的過程中起關(guān)鍵作用。
[1] Wang L, Deng Q, Wang J, et al. Effect of CIK on multidrug-resistance reversal and increasing the sensitivity of ADR in K562/ADR cells[J]. Oncol Lett, 2014,8(4):1778-1782.
[2] 王忠華,史姍姍,陳雨,等.浙貝母生物堿提取工藝研究[J].中藥材,2010,33(1):128-132.
[3] 諶海燕,陳信義.貝母素甲抑制人乳腺癌細(xì)胞MCF-7/TAM增殖及其對(duì)細(xì)胞凋亡的影響[J].中醫(yī)藥學(xué)報(bào),2012,40(4):12-15.
[4] 唐曉勇,唐迎雪.浙貝母堿對(duì)肺癌A549/DDP細(xì)胞多藥耐藥的逆轉(zhuǎn)作用觀察及機(jī)制探討[J].山東醫(yī)藥,2012,52(18):4-6.
[5] 胡凱文,鄭洪霞,齊靜,等.浙貝母堿逆轉(zhuǎn)白血病細(xì)胞多藥耐藥的研究[J]. 中華血液學(xué)雜志,1999,20(12):33-34.
[6] Cen JA, Qi Y, Tao YF, et al. HZ08, a great regulator to reverse multidrug resistance via cycle arrest and apoptosis sensitization in MCF-7/ADM[J]. Eur J Pharmacol, 2010,647(1-3):21-30.
[7] Kalinina E, Novichkova M, Scherbak NP, et al. GSH-dependent redox regulation and antioxidant enzymes in the formation of resistance to doxorubicin in K562 human erythroleukemia cells[J]. Adv Exp Med Biol, 2001,500:241-244.
[8] Yao JY, Wei X, Lu YH. Chaetominine reduces MRP1-mediated drug resistance via inhibiting PI3K/Akt/Nrf2 signaling pathway in K562/Adr human leukemia cells[J]. Biochem Bioph Res Co, 2016,473(4):867-873.
[9] 石倩倩,丁亞輝,祁瑞哲,等.小白菊內(nèi)酯誘導(dǎo)耐藥白血病細(xì)胞K562/ADR凋亡的研究[J]. 中國藥理學(xué)通報(bào),2012,28(12):1736-1740.
[10] 鄒凌琳,周圓,楊銘,等.鈣調(diào)素拮抗劑O-(4-乙氧基-丁基)-小檗胺對(duì)人白血病細(xì)胞K562和K562/A02生長的抑制作用[J]. 中國藥理學(xué)通報(bào),2009,25(10):1313-1317.
[11] Zhou QX, Chen YL, Chen X, et al. In Vitro Antileukemia Activity of ZSTK474 on K562 and Multidrug Resistant K562/A02 Cells[J]. Int J Biol Sci, 2016,12(6):631-638.
[12] Rumjanek VM, Vidal RS, Maia RC. Multidrug resistance in chronic myeloid leukaemia: how much can we learn from MDR-CML cell lines[J]. Biosci Rep, 2013,33(6):875-888.
[13] Mathisen MS, Kantarjian HM, Cortes J, et al. Practical issuessurrounding the explosion of tyrosine kinase inhibitors for the management of chronic myeloid leukemia[J]. Blood Rev, 2014,28(5):179-187.
[14] 董憲喆,畢明剛.活性氧對(duì)P-糖蛋白調(diào)節(jié)作用的研究進(jìn)展[J].中國藥理學(xué)通報(bào),2010,26(10):1386-1390.
[15] Wang D, Jiang Y, Wu K, et al. Evaluation of antitumor property of extracts and steroidal alkaloids from the cultivated Bulbus Fritillariae ussuriensis and preliminary investigation of its mechanism of action[J]. BMC complementary and alternative medicine, 2015,15:29.
[16] 李冬云,陳信義,姜靖雯.復(fù)方浙貝顆粒研究現(xiàn)狀與應(yīng)用前景分析[J].中國藥物與臨床,2009,9(2):85-87.
[17] 李偉,胡凱文,蘇偉,等.浙貝母散劑逆轉(zhuǎn)急性白血病多藥耐藥的臨床研究[J].北京中醫(yī)藥大學(xué)學(xué)報(bào),2004,27(1):63-65.
[18] Park YK, Do YR, Jang BC. Apoptosis of K562 leukemia cells by Abnobaviscum F (R), a European mistletoe extract[J]. Oncol Rep, 2012,28(6):2227-2232.
[19] Zhang J, Cao M, Yang WW, et al. Inhibition of Glucose-6-Phosphate Dehydrogenase Could Enhance 1,4-Benzoquinone-Induced Oxidative Damage in K562 Cells[J]. Oxid Med Cell Longev, 2016,21:1-11.
[20] 鄭智,侯麗,許亞梅,等.復(fù)方浙貝顆粒聯(lián)合阿霉素影響K562/A02移植瘤細(xì)胞耐藥相關(guān)酶表達(dá)研究[J]. 醫(yī)學(xué)研究雜志,2009,38(12):29-31,146.
[21] Wang DD, Feng Y, Li Z, et al. In vitro andin vivo antitumor activity of Bulbus Fritillariae Cirrhosae and preliminaryinvestigation of its mechanism[J]. Nutr Cancer, 2014,66(3):441-452.
[22] Wang DD, Wang S, Feng Y, et al. Antitumor effects of Bulbus Fritillariae cirrhosae on Lewis lung carcinoma cells in vitro and in vivo[J]. Ind Crops Prod, 2014,54:92-101.
[23] Wang XD, Li CY, Jiang MM, et al. Induction of apoptosis in human leukemia cells through an intrinsic pathway by cathachunine, a unique alkaloid isolated from Catharanthus roseus[J]. Phytomedicine, 2016,23(6):641-653.
[24] Luo J, Kong W, Yang M. HJC, A new arylnaphthalene lignan isolated from Justicia procumbens, causes apoptosis and caspase activation in K562 leukemia cells[J]. J Pharmacol Sci, 2014,125(4):355-363.
Effects of peimine on cell viability and apoptosis of multidrug resistant human leukemia cells
QIYan,LIAOBin,XUChengbo,HUANGFUZhenping,CHENJiawei
(People′sHospitalAffiliatedtoFujianUniversityofTraditionalChineseMedicine,Fuzhou350004,China)
Objective To investigate the effects of peimine on cell viability and apoptosis of multidrug resistant human leukemia K562/A02 cells and the function of reactive oxygen species (ROS) in this progress. Methods The K562/A02 cells were divided into three groups. Cells in the drug group were treated with peimine with the final concentrations of 100, 200, and 400 μmol/L, respectively. Cells in the N acetyl cysteine (NAC) pretreatment group were treated with 5 mmol/L antioxidant NAC, followed by 400μmol/L peimine. Cells in the control group were treated with an equal volume of dissolution medium RPMI1640. The cell viability of each group was detected by MTT, the apoptosis was detected by flow cytometry, and the levels of ROS and glutathione (GSH) in the cells were detected by UV spectrophotometry. Results Compared with the control group, the cell viability of the drug group decreased (P<0.05 orP<0.01), intracellular ROS level increased (P<0.01), GSH concentration decreased (P<0.01), and the apoptosis rate increased (P<0.05). However, the cell viability, the apoptosis rate, and the ROS level in NAC pretreatment group showed no obvious change (allP>0.05). Conclusion Peimine inhibits the cell viability and induces apoptosis of multidrug resistant leukemia K562/A02 cells through inducing the ROS outbreak and thus increasing the GSH content.
peimine; chronic myeloid leukemia; K562/A02; multidrug resistance; reactive oxygen species; cell viability; apoptosis
福建省教育廳中青年教師教育科研項(xiàng)目(JA14161)。
齊彥(1977-),女,副主任醫(yī)師,碩士,主要研究方向?yàn)橹形麽t(yī)結(jié)合治療血液病。E-mail: 3362313804@qq.com
廖斌(1961-),男,主任醫(yī)師,碩士,主要研究方向?yàn)橹形麽t(yī)結(jié)合治療血液病。E-mail: liaobin1961@qq.com
10.3969/j.issn.1002-266X.2017.26.005
R733.7
A
1002-266X(2017)26-0017-04
2017-03-30)
猜你喜歡
保健醫(yī)苑(2022年5期)2022-06-10 07:46:38
現(xiàn)代臨床醫(yī)學(xué)(2022年3期)2022-06-06 07:59:40
昆明醫(yī)科大學(xué)學(xué)報(bào)(2022年1期)2022-02-28 07:43:40
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
科學(xué)大眾(2020年12期)2020-08-13 03:22:22
海峽科技與產(chǎn)業(yè)(2016年3期)2016-05-17 04:32:12
