一株地衣芽孢桿菌產(chǎn)凝乳酶酶學(xué)性質(zhì)研究
2017-08-22 06:26:53劉圓媛
中國(guó)食物與營(yíng)養(yǎng) 2017年7期
宋 曦,汪 慧,尚 菲,韓 雍,劉圓媛
(1隴東學(xué)院農(nóng)林科技學(xué)院,甘肅慶陽(yáng) 745000;2隴東學(xué)院生命科學(xué)技術(shù)學(xué)院,甘肅慶陽(yáng) 745000)
一株地衣芽孢桿菌產(chǎn)凝乳酶酶學(xué)性質(zhì)研究
宋 曦1,汪 慧1,尚 菲2,韓 雍1,劉圓媛1
(1隴東學(xué)院農(nóng)林科技學(xué)院,甘肅慶陽(yáng) 745000;2隴東學(xué)院生命科學(xué)技術(shù)學(xué)院,甘肅慶陽(yáng) 745000)
目的:對(duì)地衣芽孢桿菌所產(chǎn)凝乳酶的酶學(xué)特性進(jìn)行研究。方法:在不同的溫度、pH值、底物濃度、不同金屬離子等條件下測(cè)定地衣芽孢桿菌產(chǎn)凝乳酶相對(duì)酶活力。結(jié)果:地衣芽孢桿菌所產(chǎn)凝乳酶最適凝乳溫度為70℃;40℃以上熱處理后凝乳活性有不同程度的損失,75℃熱處理10min后凝乳酶活性喪失;pH值為5~8時(shí)凝乳活性隨乳pH值的降低而增強(qiáng),pH值為7時(shí)凝乳酶最穩(wěn)定;Ca2+、Fe2+、Fe3+、Mn2+、Mg2+、Al3+對(duì)酶活性有一定的促進(jìn)作用;Li2+、Na+、Cu2+、Zn2+對(duì)酶活性有抑制作用;底物濃度最適為250 g/L、Ca2+的最適濃度為0.014 g/L。
地衣芽孢桿菌;凝乳酶
凝乳酶在干酪加工過(guò)程中主要作用為專(zhuān)一性地水解乳中κ-酪蛋白第105~106 位苯丙氨酸和甲硫氨酸之間的肽鍵,形成穩(wěn)定的副κ-酪蛋白及親水性的糖巨肽[1-3]。國(guó)際市場(chǎng)上凝乳酶中動(dòng)物性來(lái)源的約占70%、微生物性的占30%、植物性的不到1%。微生物產(chǎn)凝乳酶不受自然條件影響,生產(chǎn)成本低,可大規(guī)模工業(yè)化生產(chǎn),所以成為國(guó)內(nèi)外的研究重點(diǎn)[2-8]。
本試驗(yàn)從甘肅省天祝藏族自治縣牦牛牧區(qū)土壤中分離得到高產(chǎn)凝乳酶菌株1株,對(duì)其進(jìn)行分子鑒定表明該菌株與地衣芽孢桿菌BacilluslicheniformisstrainN8 和BacilluslicheniformisstrainYP1A同源性為99%,標(biāo)記為D3.11。
1 材料與方法
1.1 材料與儀器設(shè)備
菌種:實(shí)驗(yàn)室篩選菌種,經(jīng)16sRNA序列分析比對(duì),確定為地衣芽孢桿菌。
試劑:酪蛋白、牛肉膏、磷酸氫二鈉、氯化鈉、瓊脂、蒸餾水、10%脫脂牛乳、硫酸銨等。
儀器與設(shè)備:冷凍離心機(jī)、電子分析天平、顯微鏡、搖床、恒溫培養(yǎng)箱,超凈工作臺(tái),試管(若干)、接種針、接種槍、酒精燈、玻璃棒、紗布、棉線、鐵架臺(tái)、移液管、三角瓶(若干)、培養(yǎng)皿等。
1.2 方法
1.2.1 測(cè)定方法
(1)硫酸銨鹽析用量條件的確定:低溫條件下向發(fā)酵液中緩慢加入研細(xì)的硫酸銨粉末至20 %~30 %飽和度,靜置3 h,在12 000 r/min、4 ℃下離心10 min,分別收集上清液。同樣條件下,再緩慢向上清液中加入硫酸銨至70%、80%、90%、100%飽和度,沉淀過(guò)夜,在12 000 r/min、4 ℃下離心10 min,分別收集沉淀,溶于pH為6.0的磷酸鹽緩沖溶液中待測(cè)用。
(2)凝乳酶凝乳活性的測(cè)定:采用Arima方法[9],取5 mL 100g/L的脫脂乳,在一定溫度下保溫5 min,加入1 mL酶液,迅速混合均勻,準(zhǔn)確記錄從加入酶液到乳凝固的時(shí)間(s)。把40 min凝固1 mL 100 g/L脫脂乳的酶量定義為一個(gè)索氏單位。

T—凝乳時(shí)間;D—稀釋倍數(shù)

1.2.2 試驗(yàn)方法
(1)酶的最適凝乳溫度:將酶液分別在25、30、35、40、45、50、55、60、65、70 ℃下保溫5 min 后測(cè)定酶活力,以35 ℃時(shí)的酶活力為對(duì)照。
(2)酶的熱穩(wěn)定性:將酶液分別在40、50、60、70 ℃下保溫10、20、30、40、50、60 min后,測(cè)定酶活力,以35 ℃時(shí)的酶活為力為對(duì)照。
(3)脫脂乳的pH值對(duì)酶凝乳活性的影響:用0.1 mol/L HCl和0.1 mol/L NaOH將100 g/L的脫脂乳的pH分別調(diào)節(jié)到5.5、5.8、6.1、6.4、6.7在酶最適宜的凝乳溫度下測(cè)定酶的凝乳活性,以脫脂乳自然pH值(6.5)下酶活力為對(duì)照。
(4)不同pH值下的穩(wěn)定性:用0.1 mol/L HCl和0.1mol/L NaOH將酶液調(diào)節(jié)到pH為2、3、4、5、6、7、8、9、10、11、12,在室溫下放置過(guò)夜后,再將酶液pH調(diào)節(jié)到酶最適pH值,在酶最適溫度下測(cè)定酶活力,以酶液pH自然時(shí)的酶活作為對(duì)照。
(5)Ca2+對(duì)酶凝乳活性的影響:脫脂乳中添加CaCl2,使其含量分別為0.004、0.006、0.008、0.012、0.014、0.016、0.018 g/L,35 ℃下測(cè)定凝乳酶活,以CaCl2添加量為 0.011 g/L的脫脂乳時(shí)的酶活作為對(duì)照。
(6)金屬離子對(duì)酶的凝乳活性的影響:配制KCl,MgSO4、LiSO4、FeCl2、NaCl、MnSO4、Fe(SO4)3、Al(SO4)3、CuSO4、ZnCl含量分別為0.05 mol/L的脫脂乳溶液,在酶的最適作用溫度下測(cè)定酶活力,以未添加金屬離子的10 %脫脂乳時(shí)的酶活為對(duì)照。
(7)底物濃度對(duì)酶的凝乳活性的影響:脫脂奶粉配制成10、50、100、150、200、250 g/L溶液,35 ℃下測(cè)定酶活力,以脫脂乳為10 %的酶活為對(duì)照。
2 結(jié)果與分析
2.1 硫酸銨用量的確定
將收集沉淀溶解于pH 6.8的磷酸緩沖液中,測(cè)定凝乳酶活力。表1可知,酶活在初始硫酸銨濃度為20 %時(shí)最高,其原因可能是30 %時(shí)濃度過(guò)大,而使酶蛋白有一定的沉淀。在同樣條件下緩慢向上清液中加入硫酸銨至70 %、80 %、90 %、100 %飽和度,沉淀過(guò)夜,測(cè)得在加入硫酸銨至濃度為70 %時(shí)酶活力最高。
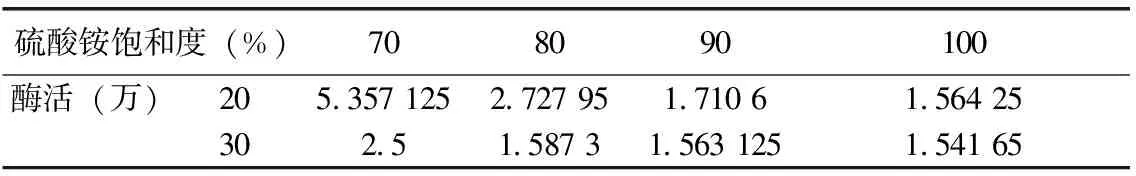
表1 不同硫酸銨飽和度下的酶活
2.2 酶的最適凝乳溫度
將已分離純化好的酶液分別在25~70 ℃范圍內(nèi)保溫10 min后測(cè)定凝乳酶活力。圖1結(jié)果表明,D3.11菌株所產(chǎn)凝乳酶在35~70 ℃有較高的凝乳活力,凝乳最適溫度為70 ℃,當(dāng)溫度低于30 ℃時(shí)凝乳速度緩慢。在溫度為35~70 ℃之間凝乳酶活力較高,從30 ℃開(kāi)始凝乳酶活力與溫度成正比,高于70 ℃時(shí),活力迅速下降,升至75 ℃時(shí)失活。在酶蛋白的活性范圍內(nèi),隨著溫度的升高會(huì)加快酶促反應(yīng)的進(jìn)行,但高于酶的最適反應(yīng)溫度后,高溫會(huì)使凝乳酶性質(zhì)發(fā)生改變,甚至使酶失去活力,所以導(dǎo)致相對(duì)酶活力迅速降低,當(dāng)凝乳酶失去活性后,其相對(duì)凝乳活力為零。
2.3 酶的熱穩(wěn)定性
將已分離純化好的酶液分別在40、50、60、70 ℃下保溫10、20、30、40、50、60 min,測(cè)定其剩余活力,表2結(jié)果表明,凝乳酶在40 ℃下處理60 min,凝乳活性比處理前有所增高;在50 ℃下處理10 min時(shí)酶活較處理前有所提高,但繼續(xù)處理20、30、40、50、60 min后酶活力逐漸降低;在60 ℃下處理10、20、30、40、50、60 min后酶活力大幅度降低,處理60 min時(shí)活力喪失73.1%;70 ℃下處理10 min后凝乳酶活力即完全喪失。酶的變性不僅取決與溫度的高低,還與該溫度下保持的時(shí)間長(zhǎng)短有關(guān),在高于變性溫度下,酶在很短時(shí)間內(nèi)出現(xiàn)高凝乳活性,但隨時(shí)間延長(zhǎng),活性逐漸損失,直至完全失活。

圖1 不同溫度下凝乳酶的相對(duì)酶活
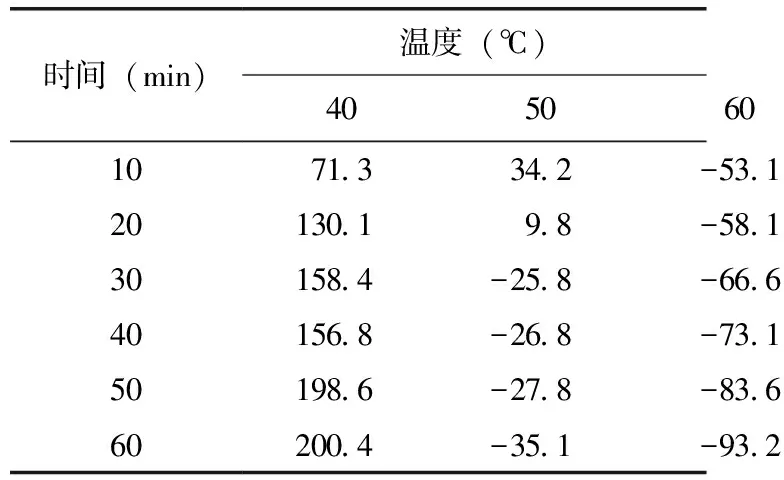
表2 一定溫度下保持一定時(shí)間后凝乳酶的相對(duì)酶活
2.4 底物pH值對(duì)凝乳酶活力的影響
將脫脂乳pH值分別調(diào)節(jié)為pH 5.5、5.8、6.1、6.4、6.7后測(cè)定凝乳酶活力。由圖2可知,隨著pH值的升高,凝乳活力整體呈下降趨勢(shì),在酸性條件下酶具有較高的凝乳活力,pH為5.5時(shí)凝乳活力最高。當(dāng)pH為6.7時(shí)凝乳活力則下降很快,不到對(duì)照試驗(yàn)的20 %。出現(xiàn)以上現(xiàn)象是因?yàn)閜H值的變化會(huì)影響到酶的空間結(jié)構(gòu),可能導(dǎo)致酶活性部位可基團(tuán)改變,酶分子活性中心催化基團(tuán)和結(jié)合基團(tuán)的離解,底物的離解及酶~底物復(fù)合物的離解,從而影響反應(yīng)動(dòng)力學(xué),影響酶反應(yīng)速度。
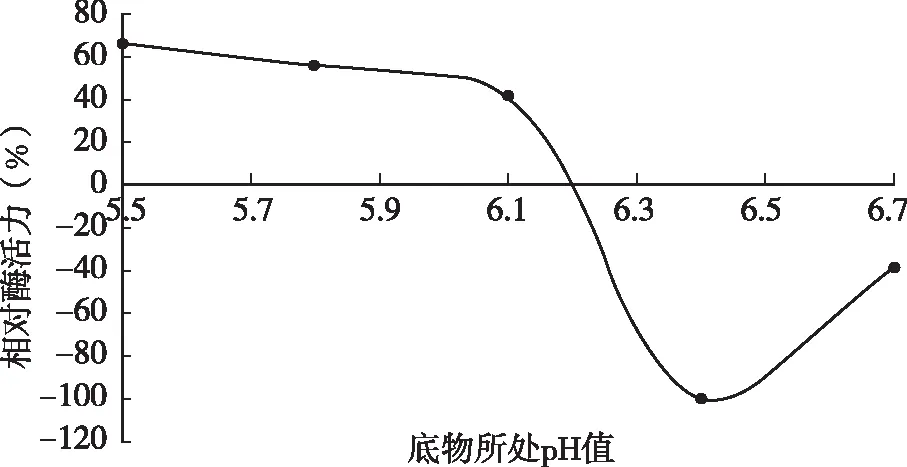
圖2 脫脂乳pH值對(duì)凝乳酶活性的影響
2.5 酶液在不同pH值時(shí)的穩(wěn)定性
用0.1mol/L的HCl和NaOH將不同酶液調(diào)至pH為2、3、4、5、6、7、8、9、10、11、12,室溫下放置過(guò)夜,然后將pH值調(diào)到6.0,測(cè)定凝乳酶活力。圖3結(jié)果表明,D3.11菌株所產(chǎn)凝乳酶在弱酸和中性偏堿的環(huán)境中都還保留有一定的凝乳活力,在pH為7時(shí)較穩(wěn)定,超過(guò)pH 7值,酶的穩(wěn)定性下降,活力受影響,甚至完全失活。當(dāng)pH高于7.0或低于2.0時(shí)酶活損失嚴(yán)重,說(shuō)明該凝乳酶在強(qiáng)酸,強(qiáng)堿條件下都不穩(wěn)定。過(guò)酸或過(guò)堿會(huì)影響酶蛋白的結(jié)構(gòu),甚至使酶變性而失活。
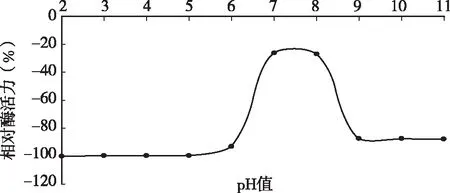
圖3 凝乳酶在不同pH值處理后酶活力
2.6 Ca2+對(duì)凝乳酶活性的影響
由圖4可知,CaCl2質(zhì)量濃度達(dá)到0.014 g/L時(shí)凝乳活力呈增加趨勢(shì),低濃度的Ca2+對(duì)酶的凝乳活力影響較小,到0.018 g/L趨于穩(wěn)定;從整體上看Ca2+加速了凝乳過(guò)程。當(dāng)CaCl2添加量不足時(shí),凝乳時(shí)間延長(zhǎng),而且凝塊組織松散;反之,添加過(guò)多時(shí),雖然凝乳速度加快,但凝塊堅(jiān)實(shí),所以CaCl2的添加量應(yīng)該在一個(gè)穩(wěn)定的范圍內(nèi),此凝乳酶在0.018 g/L趨于穩(wěn)定。
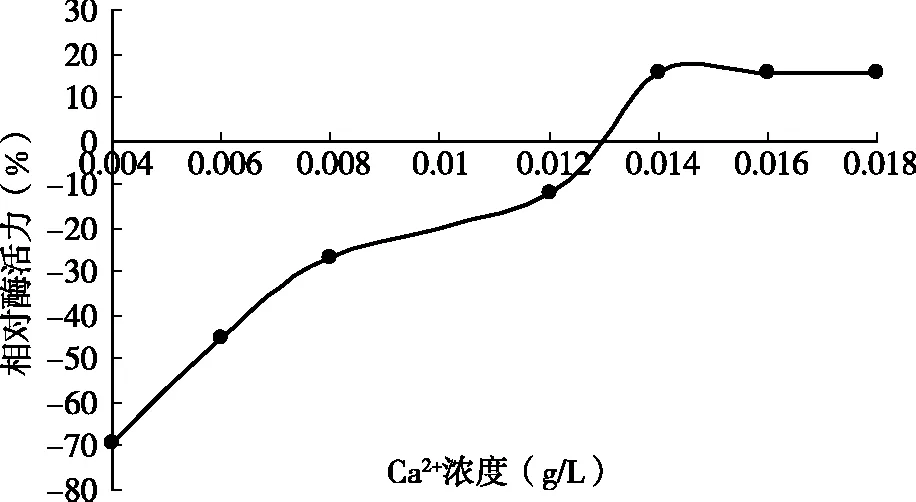
圖4 Ca2+對(duì)酶凝乳效果的影響
2.7 其他金屬離子對(duì)酶的凝乳效果的影響
當(dāng)脫脂乳中不同金屬離子的含量為0.05 mol/L時(shí),測(cè)定凝乳酶活力。由表3可知,F(xiàn)e2+、Fe3+、Mn2+、Mg2+、Al3+對(duì)凝乳活力有顯著的促進(jìn)作用,其中Fe2+最大可使凝乳活力提高46.3%,Mg2+最小也有1.6 %的促進(jìn)作用。Li2+、Na+、Cu2+、Zn2+對(duì)凝乳活力有抑制作用,Cu2+對(duì)凝乳活力抑制能力最大,達(dá)到60.8%,而Li2+、Na+抑制作用最小都達(dá)到13.1 %。k+對(duì)凝乳活力的影響不明顯。
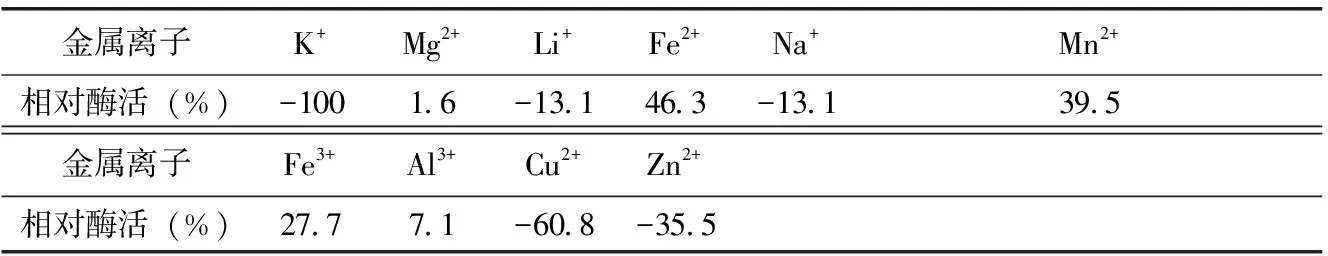
表3 不同金屬離子對(duì)凝乳酶活力影響
2.8 底物濃度對(duì)酶的凝乳效果的影響
由圖5可知,脫脂乳濃度分別為10、50、100、150、200、250 g/L時(shí)測(cè)定凝乳酶活力,結(jié)果表明脫脂乳的不同濃度對(duì)凝乳酶的酶活力有一定程度的影響,當(dāng)脫脂乳液的濃度高于一定程度時(shí),酶的活力有所減弱。底物濃度越大,凝乳酶活性越高,當(dāng)?shù)孜餄舛刃∮?00 g/L時(shí),隨著底物濃度的增大,酶凝乳活性增加較快,當(dāng)大于200g/L時(shí),酶的凝乳活性趨于穩(wěn)定,凝乳酶活力最大。
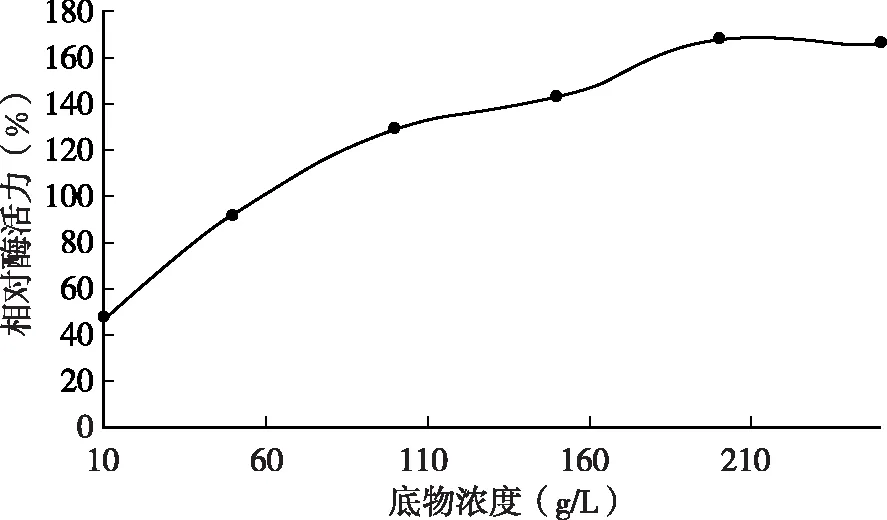
圖5 不同脫脂乳濃度時(shí)的凝乳酶相對(duì)酶活
3 結(jié)論與討論
(1)菌株D3.11產(chǎn)凝乳酶分離時(shí)采用硫酸銨為初始飽和度為20 %,加入硫酸銨到飽和度為70 %時(shí)效果最佳。(2)菌株D3.11產(chǎn)凝乳酶的最適凝乳溫度為70℃。在40 ℃下處理60 min,酶的活力較穩(wěn)定,凝乳活性比處理前有所增高;在50 ℃下處理10 min酶活較處理前有所提高,但繼續(xù)處理20、30、40、50、60 min酶活力逐漸降低;70℃下處理10 min后酶活力喪失。說(shuō)明該菌株產(chǎn)凝乳酶耐高溫,適合于工業(yè)生產(chǎn)。國(guó)內(nèi)薛璐等人對(duì)江米酒凝乳酶酶學(xué)特性的研究表明,其溫度在65℃后酶活力喪失[10]。高維東等[11]對(duì)微小毛霉凝乳酶酶學(xué)性質(zhì)的研究表明,其最適溫度為60、70℃后酶活力喪失。王博達(dá)等[12]將解淀粉芽孢桿菌所產(chǎn)凝乳酶進(jìn)行克隆、重組表達(dá)后發(fā)現(xiàn)重組后的凝乳酶在70℃時(shí)酶活力最高,說(shuō)明D3.11所產(chǎn)凝乳酶耐高溫性能較好。(3)pH為5.5時(shí)凝乳活力最高。菌株D3.11產(chǎn)凝乳酶,強(qiáng)堿條件下都不穩(wěn)。所以在該凝乳酶的應(yīng)用過(guò)程中,必須將酶的反應(yīng)環(huán)境pH值控制在弱酸性條件下。CARIAS J R等[13]對(duì)黃色粘球菌產(chǎn)凝乳酶研究表明,該菌在pH 6.0時(shí)酶活力最高。CAVALCANTI M T H等[14-15]對(duì)諾卡氏菌所產(chǎn)凝乳酶研究表明,其最適pH值為11.0和7.5,即在堿性條件下較穩(wěn)定,可見(jiàn)不同細(xì)菌所產(chǎn)凝乳酶最適pH值相差較大。(4)Fe2+、Fe3+、Mn2+、Mg2+、Al3+對(duì)菌株D3.11產(chǎn)凝乳酶凝乳活力有顯著的促進(jìn)作用。Li2+、Na+、Cu2+、Zn2+對(duì)菌株D3.11產(chǎn)凝乳酶凝乳活力有抑制作用。K+對(duì)菌株D3.11產(chǎn)凝乳酶的凝乳活力的影響不明顯。Ca2+最適濃度為0.014 g/L,Ca2+參與乳的凝固過(guò)程,當(dāng)酶作用于酪蛋白形成福κ-酪蛋白時(shí),Ca2+可加速副κ-酪蛋白的進(jìn)一步聚集而凝固。同時(shí)副κ-酪蛋白對(duì)凝乳酶本身也有一定的影響[3,16]。脫脂乳濃度為200g/L時(shí),酶活力最大。◇
[1]劉顯庭,張昊,郭慧媛,等.凝乳酶在干酪生產(chǎn)中的應(yīng)用[J].中國(guó)乳業(yè),2013(133):54-57.
[2]郭本恒.乳品化學(xué)[M].北京:中國(guó)輕工業(yè)出版社,2001:7.
[3]BRUNO M A,et al.Milk cloting and proteolytic activity of an enzyme preparation from Bromelia hieromymi fruits[J].LWT-Food Science and Technology,2010,43(4):697-701.
[4]高維東,甘伯中,丁福軍,等.微生物凝乳酶的研究進(jìn)展[J].中國(guó)乳品工業(yè),2009,37(5):34-36.
[5]張衛(wèi)兵,宋曦,賀曉玲,等.Bacillus licheniformis產(chǎn)凝乳酶培養(yǎng)基的優(yōu)化[J].中國(guó)釀造,2011,30(2):70-73.
[6]Prados,PINO A,F(xiàn)ernandez-salguero J.Effect of a powdered begetable coagulant from Cynara cardunculus in the accelerated ripening of Mznchego cheese[J].Int J Food Sci Technol,2007,42(5):556-561.
[7]杜琨,張亞寧.干酪的營(yíng)養(yǎng)價(jià)值及研究動(dòng)態(tài)[J].中國(guó)食物與營(yíng)養(yǎng),2005(10):45-46.
[8]杭鋒,洪青,王欽博,等.凝乳酶的研究進(jìn)展[J].食品科學(xué),2016,30(3):273-279.
[9]K Arima,I Shinjiro,T Gakuzo.Milk-clltting enzymes from microorganinism,Part InScreening test and identification of potent fungus[J].Agric Biol Chem,1967,31(5):540-545.
[10]薛璐,陳歷俊,姜鐵民,等.江米酒凝乳酶酶學(xué)特性的研究[J].食品科學(xué),2008,29(4):259-262.
[11]高維東,甘伯中,丁福軍,等.微小毛霉凝乳酶的酶學(xué)性質(zhì)研究[J].食品科學(xué),2010,31(3):185-188.
[12]王博達(dá),丁重陽(yáng),張梁,等.解淀粉芽孢桿菌凝乳酶的克隆、重組表達(dá)及酶學(xué)性質(zhì)[J].工業(yè)微生物,2016,45(6):9-15.
[13]CARIAS J R,RAUBGEAUD J,MAZAUD C,et al.A chymosin-like extracellular acidic endoprotease from Myxococcus xanthus[J].J Bacteriol,1992,174(15):5136-5140.
[14]CAVALCANTI M T H,et al.Partial purification of new milk-clotting enzyme produced by Nocardiopsis sp.[J].Bioresource Technol,2004,93(1):29-35.
[15]CAVALCANTI M T H,et al.Milk clotting protease production by Nocardiopsis sp.in an inexpensive medium[J].World J Microb Biot,2005,21(2):151-154.
[16]FOLTMANN B,et al.The primary sructure of calf chymosin[J].J Biol Chem,1979(254):8447-8456.
(責(zé)任編輯 李婷婷)
The Properties of Chymosin fromBacilluslicheniformis
SONG Xi1,WANG Hui1,SHANG Fei2,HAN Yong1, LIU Yuan-yuan1
(1College of Agriculture and Forestry;2College of life Science and Technology,Longdong University,Qingyang 745000,China)

Bacilluslicheniformis;chymosin
宋曦(1984— ),女,碩士研究生,講師,研究方向:食品營(yíng)養(yǎng)與衛(wèi)生。
