MCM2-7復合體參與DNA復制機制及調控特點
2017-08-29 10:45:24王明剛毛德文張榮臻龍富立王秀峰
胃腸病學和肝病學雜志 2017年7期
王明剛,毛德文,張榮臻,龍富立,王秀峰
廣西中醫藥大學第一附屬醫院肝病一區,廣西 南寧 530023
綜述
MCM2-7復合體參與DNA復制機制及調控特點
王明剛,毛德文,張榮臻,龍富立,王秀峰
廣西中醫藥大學第一附屬醫院肝病一區,廣西 南寧 530023
脫氧核糖核酸(deoxyribonucheic acid, DNA)無差錯復制是基因組信息成功遺傳到子代的主要保障,對于生命的遺傳信息傳遞和延續意義重大。微小染色體維持蛋白(minichromosome maintenance proteins, MCM)家族是廣泛存在的高保守蛋白,MCM2-7六聚體是DNA復制所必須的。MCMs基因轉錄與啟動子區E2F結構域密切相關,在基因表達層面可受到miRNA-34s和miRNA-1269的影響。在MCM2-7生物學功能過程中可受到Cdt1、cyclinE、p27Kip1、Rb、ATM、ATR蛋白的調控和影響。
DNA復制;微小染色體維持蛋白;調控;微小核糖核酸
脫氧核糖核酸(deoxyribonucleic acid,DNA)是由兩條脫氧多核苷酸鏈反向平行盤繞所形成的雙螺旋結構,內部由許多脫氧核苷酸按一定堿基順序以3′,5′-磷酸二酯鍵相連構成[1]。DNA是生命遺傳信息的主要存儲形式。DNA復制是單鏈模板DNA復制成兩個互補DNA鏈的生物學過程。對于細胞分裂,DNA復制是必須的,且多發生在細胞周期的S期,一個細胞周期DNA復制僅發生一次。DNA無差錯復制是基因組信息成功遺傳到子代的主要保障,對于生命的遺傳信息傳遞和延續意義重大[2]。
微小染色體維持蛋白(minichromosome maintenance proteins, MCM)家族是高度保守的DNA解旋蛋白復合體,其保守性可追溯到上古時代單細胞形成[3]。在細胞內MCM2、MCM3、MCM4、MCM5、MCM6、MCM7形成六聚體才具有解螺旋酶活性,MCMs蛋白功能正常是DNA復制所必須的[4]。新近研究發現,MCMs蛋白家族除了經典的MCM2-7外,MCM1、MCM8、MCM9、MCM10相繼被發現和證實[5-7]。本文總結了近年來經典MCM2-7蛋白復合體參與DNA復制機制及該蛋白復合體被調控的特點。
1 真核細胞DNA復制進程
DNA復制是真核細胞有絲分裂S期的重要生物學事件,其主要分為DNA復制起始和延伸兩個部分[8]。DNA 復制源識別復合物 (origin recognition complex, ORC)首先識別并結合到復制起始位點上,作為預復制復合物組裝平臺(pre-replication complex, pre-RC)[9]。隨后招募Cdc6、Cdt1和MCM2-7蛋白,完成pre-RC組裝[10]。真核細胞有絲分裂進入S期后,CDK和DDK蛋白將依次磷酸化激活pre-RC蛋白,活化的pre-RC進一步招募Cdc45 和 GINS形成CMG復合物(Cdc45-MCM-GINS complex),CMG復合物具有解螺旋活性可激活起始復制源[11]。后續RPA、RFC、PCNA及 DNA 聚合酶依次結合到被激活的復制源上開始DNA復制進程[12]。
2 MCMs蛋白結構的特點及功能學基礎
MCMs蛋白首先在釀酒酵母中被發現[13],結構研究表明MCM蛋白是AAA+ATPase家族的一個亞群,中央結構域是由約200個氨基酸組成的高保守結構區,MCMs蛋白形成六聚體是功能活性的基礎[14]。此外,MCM4、MCM6和MCM7在蛋白N端鑲嵌了一個鋅指結構域,該結構域對MCM形成六聚體起關鍵作用;MCM2和MCM3中具有核定位序列;MCM2和MCM4的N端具有額外的細胞周期蛋白依賴激酶(cyclin dependent kinase, CDK)結合序列[15]。在MCM2-7六聚體中,MCM4、MCM6和MCM7形成核心,其他MCMs蛋白依次結合,依賴復合體核定位序列進入細胞核和CDK結合序列招募CDK蛋白[16]。
3 MCM2-7參與的DNA復制過程
MCM2-7(MCM2、MCM3、MCM4、MCM5、MCM6、MCM7)蛋白六聚體是較早被發現的MCM蛋白功能復合體,復合體中MCM蛋白的特殊結構是實現核-膜穿梭和磷酸化激活的基礎。晶體研究顯示MCM2-7復合體呈指環狀,中間有足夠容納dsDNA通過的孔道[17]。DNA復制過程中dsDNA從中間孔道通過,解螺旋后的ssDNA從側邊孔道延出[18]。在真核細胞有絲分裂G期MCM2-7復合體與染色體結合,完成預復制復合物組裝,進入S期后,MCM2-7復合體被激酶活化,誘發下游聚合酶-引物合成酶裝配,同時和DNA聚合酶結合,形成功能性復制叉,開始DNA解螺旋與復制[19]。MCM2-7必須在G期結合到起始點上,缺少MCM2-7的結合,DNA復制將不能進行[20]。
4 MCM2-7的轉錄調控
MCM2-7基因的轉錄有明顯的周期性規律,具體表現在基因轉錄mRNA表達的高峰在細胞周期的G1晚期[21],進入S期后mRNA表達將受到抑制[22]。在哺乳動物中,MCMs基因轉錄與啟動子區E2F結構域密切相關,細胞轉錄因子E2F結合到啟動子特定區域是MCMs基因轉錄所必須的[23]。在正常情況下E2F與Rb蛋白結合,抑制E2F與啟動子結合。當細胞進入分裂周期以后,cyclin D結合CDK4與CDK6磷酸化Rb蛋白,Rb蛋白磷酸化后,E2F從復合體中解離,下游轉錄大量蛋白,包括MCMs蛋白家族,推進細胞周期變化[24]。在細胞周期的G1晚期,Rb蛋白被磷酸化,E2F從復合體中大量解離,結合到啟動子區E2F結構域上,轉錄靶基因(見圖1)。
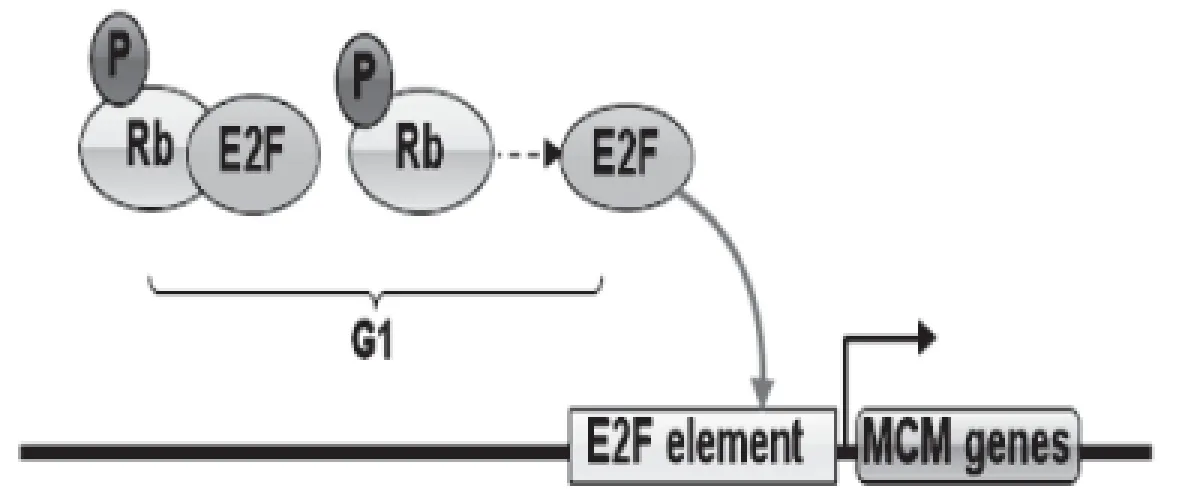
圖1 E2F參與的MCM基因轉錄 Fig 1 E2F transcription involved in MCM gene
5 MCM2-7功能過程的調控
MCM2-7生物學功能的調控可分為間接調控和直接調控。在預復制復合物組裝過程中,Geminin蛋白可與Cdt1緊密結合,阻斷Cdt1與Cdc6結合及下游招募MCM2-7[25]。CDKs可磷酸化Cdc6,促進Cdc6降解,MCM2-7復合體將不能與染色體結合[26]。在哺乳動物中,cyclin E蛋白是MCM2-7復合體與染色體結合所必須的[27]。另外,MCM2-7復合體可受到一些因子的直接調控,p27Kip1可與MCM7相互作用,誘導細胞周期阻滯[28]。負性調控基因產物Rb蛋白可與MCM7相互作用,影響細胞周期[29]。細胞周期點檢驗蛋白ATM和ATR可介導MCM2和MCM3蛋白磷酸化,影響MCM2-7復合體的ATP酶活性和解鏈酶活性[30]。
在正常細胞周期下Cdc6 結合Cdt1招募MCM2-7,完成pre-RC組裝。Geminin蛋白可與Cdt1緊密結合,阻斷Cdt1與Cdc6結合及下游招募MCM2-7。CDKs可磷酸化Cdc6,致使Cdc6降解,阻斷MCM2-7與染色體結合(見圖2)。

圖2 MCM2-7在結合到染色體前受到的調控Fig 2 Regulation of MCM2-7 binding to chromosomes
6 miRNA參與的MCM2-7表達調控
miRNA是21-25 nt的單鏈小分子RNA,是非編碼RNA的重要組成部分,miRNA具有高度的保守性、時序性和組織特異性[31]。成熟的單鏈miRNA與蛋白質復合物miRNP結合,引導復合物通過部分互補結合靶基因mRNA 3′UTR,從而抑制靶基因翻譯,或通過直接切割互補mRNA,使目標mRNA失效[32]。目前關于miRNA與MCMs表達調控及交互影響的相關研究報道尚少。已證實miRNA-1269可靶向調控MCM2基因表達,影響前列腺癌細胞增殖[33]。基因組關聯分析發現p53表達依賴的miRNA-34家族與MCM基因3′UTR有互補序列[34],后在細胞模型中證實miRNA-34家族可靶向調控MCM2-7基因表達,影響DNA復制進程[35]。
MCMs蛋白家族在細胞中廣泛存在,其高度的保守性預示著其功能的重要性。MCM2-7復合體是維持基因組穩定和參與DNA復制所必須的。MCMs表達水平與細胞增殖狀態及再生能力呈正相關。MCMs生物學過程了MCMs基因轉錄、MCM2-7復合體形成、核膜穿梭、與起始位點結合、DNA解螺旋及最后與染色體解離等眾多環節。目前對各環節的激活原因及調控過程了解并不十分透徹,MCMs的功能和機制研究仍有進一步深入研究的必要,探索非編碼RNA與MCMs的交互調控機制將打開更廣闊的研究視野。
[1]Bartlett RS, Jette ME, King SN, et al.Fundamental approaches in molecular biology for communication sciences and disorders [J]. J Speech Lang Hear Res, 2012, 55(4): 1220-1231.
[2]Jones MJ, Colnaghi L, Huang TT. Dysregulation of DNA polymerase kappa recruitment to replication forks results in genomic instability [J]. EMBO J, 2012, 31(4): 908-918.
[3]Tye BK.MCM proteins in DNA replication [J]. Annu Rev Biochem, 1999, 68: 649-686.
[4]Tuteja N, Tran NQ, Dang HQ, et al.Plant MCM proteins: role in DNA replication and beyond [J]. Plant Mol Biol, 2011, 77(6): 537-545.
[5]Park J, Long DT, Lee KY, et al.The MCM8-MCM9 complex promotes RAD51 recruitment at DNA damage sites to facilitate homologous recombination [J]. Mol Cell Biol, 2013, 33(8): 1632-1644.
[6]Chang VK, Donato JJ, Chan CS, et al. Mcm1 promotes replication initiation by binding specific elements at replication origins [J]. Mol Cell Biol, 2004, 24(14): 6514-6524.
[7]Quan Y, Xia Y, Liu L, et al. Cell-Cycle-regulated interaction between Mcm10 and double hexameric Mcm2-7 is required for helicase splitting and activation during S phase [J]. Cell Rep, 2015, 13(11):2576-2586.
[8]Warner DF, Evans JC, Mizrahi V.Nucleotide metabolism and DNA replication [J]. Microbiol Spectr, 2014, 2(5): 10.
[9]Bell SP, Stillman B.ATP-dependent recognition of eukaryotic origins of DNA replication by a multiprotein complex [J]. Nature, 1992, 357(6374): 128-134.
[10]Diffley JF.Eukaryotic DNA replication [J]. Curr Opin Cell Biol, 1994, 6(3): 368-372.
[11]Dowell SJ, Romanowski P, Diffley JF. Interaction of Dbf4, the Cdc7 protein kinase regulatory subunit, with yeast replication origins in vivo [J]. Science, 1994, 265(5176): 1243-1246.
[12]Bell SP, Dutta A. DNA replication in eukaryotic cells [J]. Annu Rev Biochem, 2002, 71: 333-374.
[13]Maine GT, Sinha P, Tye BK.Mutants of S. cerevisiae defective in the maintenance of minichromosomes [J]. Genetics, 1984, 106(3):365-385.
[14]Gomez EB, Catlett MG, Forsburg SL.Different phenotypes in vivo are associated with ATPase motif mutations in Schizosaccharomyces pombe minichromosome maintenance proteins [J]. Genetics, 2002, 160(4): 1305-1318.
[15]Forsburg SL. Eukaryotic MCM proteins: beyond replication initiation [J]. Microbiol Mol Biol Rev, 2004, 68(1): 109-131.
[16]Remus D, Blanchette M, Rio DC, et al.CDK phosphorylation inhibits the DNA-binding and ATP-hydrolysis activities of the Drosophila origin recognition complex [J]. J Biol Chem, 2005, 280(48):39740-39751.
[17]Takahashi TS, Wigley DB, Walter JC. Pumps, paradoxes and ploughshares: mechanism of the MCM2-7 DNA helicase [J]. Trends Biochem Sci, 2005, 30(8): 437-444.
[18]Li D, Zhao R, Lilyestrom W, et al.Structure of the replicative helicase of the oncoprotein SV40 large tumour antigen [J]. Nature, 2003, 423(6939): 512-518.
[19]Baltin J, Leist S, Odronitz F, et al.DNA replication in protein extracts from human cells requires ORC and Mcm proteins [J]. J Biol Chem, 2006, 281(18): 12428-12435.
[20]Costa A, Pape T, van Heel M, et al.Structural studies of the archaeal MCM complex in different functional states [J]. J Struct Biol, 2006, 156(1): 210-219.
[21]Kimura H, Kuriyama T.Airway occlusion and airway narrowing [J]. Ryoikibetsu Shokogun Shirizu, 1994,(3): 287-290.
[22]Kimura H, Takizawa N, Nozaki N, et al.Molecular cloning of cDNA encoding mouse Cdc21 and CDC46 homologs and characterization of the products: physical interaction between P1(MCM3) and CDC46 proteins [J]. Nucleic Acids Res, 1995, 23(12): 2097-2104.
[23]Tsuruga H, Yabuta N, Hosoya S, et al.HsMCM6: a new member of the human MCM/P1 family encodes a protein homologous to fission yeast Mis5 [J]. Genes Cells, 1997, 2(6): 381-399.
[24]Andrusiak MG, McClellan KA, Dugal-Tessier D, et al.Rb/E2F regulates expression of neogenin during neuronal migration [J]. Mol Cell Biol, 2011, 31(2): 238-247.
[25]Ballabeni A, Melixetian M, Zamponi R, et al.Human geminin promotes pre-RC formation and DNA replication by stabilizing CDT1 in mitosis [J]. EMBO J, 2004, 23(15): 3122-3132.
[26]Ogino H, Ishino S, Haugland GT, et al. Activation of the MCM helicase from the thermophilic archaeon, thermoplasma acidophilum by interactions with GINS and Cdc6-2 [J]. Extremophiles, 2014, 18(5): 915-924.
[27]Hossain M, Stillman B. Opposing roles for DNA replication initiator proteins ORC1 and CDC6 in control of Cyclin E gene transcription [J]. Elife, 2016, 5: e12785.
[28]Sharma SS, Ma L, Pledger WJ. p27Kip1 inhibits the cell cycle through non-canonical G1/S phase-specific gatekeeper mechanism [J]. Cell Cycle, 2015, 14(24): 3954-3964.
[29]Numata Y, Ishihara S, Hasegawa N, et al.Interaction of human MCM2-7 proteins with TIM, TIPIN and Rb [J]. J Biochem, 2010, 147(6): 917-927.
[30]Martin L, Rainey M, Santocanale C, et al.Hypoxic activation of ATR and the suppression of the initiation of DNA replication through cdc6 degradation [J]. Oncogene, 2012, 31(36): 4076-4084.
[31]Khan HA, Zhao Y, Wang L, et al.Identification of miRNAs during mouse postnatal ovarian development and superovulation [J]. J Ovarian Res, 2015, 8: 44.
[32]Baran-Gale J, Kurtz CL, Erdos MR, et al.Addressing bias in small RNA library preparation for sequencing: a new protocol recovers microRNAs that evade capture by current methods [J]. Front Genet, 2015, 6: 352.
[33]Majid S, Dar AA, Saini S, et al.Regulation of minichromosome maintenance gene family by microRNA-1296 and genistein in prostate cancer [J]. Cancer Res, 2010, 70(7): 2809-2818.
[34]Kaller M, Liffers ST, Oeljeklaus S, et al.Genome-wide characterization of miR-34a induced changes in protein and mRNA expression by a combined pulsed SILAC and microarray analysis [J]. Mol Cell Proteomics, 2011, 10(8): M111.010462.
[35]Bai G, Smolka MB, Schimenti JC. Chronic DNA replication stress reduces replicative lifespan of cells by TRP53-dependent, microRNA-assisted MCM2-7 downregulation [J]. PLoS Genet, 2016, 12(1): e1005787.
(責任編輯:馬 軍)
MCM2-7 complex participates in the mechanism and control of DNA replication
WANG Minggang, MAO Dewen, ZHANG Rongzhen, LONG Fuli, WANG Xiufeng
Department of Hepatology, the First Affiliated Hospital of Guangxi Medical University, Nanning 530023, China
Deoxyribonucleic acid (DNA) replication error-freely is the main security of genomic information successfully inherited to offspring. There is great significance for the life of the genetic information transfer and continuation. The minichromosome maintenance proteins (MCM) family is highly conserved protein. The MCM2-7 hexamer is essential for DNA replication. MCMs gene transcription is closely related to the promoter region E2F domain, at the gene expression level, it can be affected by miRNA-34s and miRNA-1269. In biological function MCM2-7 can be subjected by Cdt1, cyclinE, p27Kip1, Rb, ATM, ATR proteins.
DNA replication; MCMs; Regulate; MicroRNA
廣西高等學校高水平創新團隊及卓越學者項目;國家中醫藥管理局“慢性重型肝炎解毒化瘀”重點研究室;國家自然科學基金課題(81460718);廣西中醫藥大學第一附屬醫院青年基金(QN14010)
王明剛,碩士,研究方向:肝衰竭肝細胞再生機制及中醫藥的調控研究。E-mail:wmgyx2012@163.com
毛德文,博士,研究方向:肝衰竭的中醫藥療效機制研究。E-mail:mdwboshi2005@163.com
10.3969/j.issn.1006-5709.2017.07.024
Q75
A
1006-5709(2017)07-0809-03
2016-08-03
