子葉損傷對(duì)苦豆子幼苗早期生長(zhǎng)的影響
2017-09-03 08:59:11劉影趙玉崔東冷家明董芳慧
草業(yè)學(xué)報(bào) 2017年8期
劉影,趙玉*,崔東,冷家明,董芳慧
(1.伊犁師范學(xué)院生物與地理科學(xué)學(xué)院,新疆 伊寧 835000;2.新疆自治區(qū)普通高校天然產(chǎn)物化學(xué)與應(yīng)用重點(diǎn)實(shí)驗(yàn)室,新疆 伊寧 835000;3.伊犁師范學(xué)院化學(xué)與環(huán)境科學(xué)學(xué)院,新疆 伊寧 835000;4.江蘇衡譜分析檢測(cè)技術(shù)有限公司,江蘇 南京 210007)
子葉損傷對(duì)苦豆子幼苗早期生長(zhǎng)的影響
劉影1,2,趙玉1,2*,崔東1,冷家明3,董芳慧4
(1.伊犁師范學(xué)院生物與地理科學(xué)學(xué)院,新疆 伊寧 835000;2.新疆自治區(qū)普通高校天然產(chǎn)物化學(xué)與應(yīng)用重點(diǎn)實(shí)驗(yàn)室,新疆 伊寧 835000;3.伊犁師范學(xué)院化學(xué)與環(huán)境科學(xué)學(xué)院,新疆 伊寧 835000;4.江蘇衡譜分析檢測(cè)技術(shù)有限公司,江蘇 南京 210007)
為探討子葉損傷對(duì)新疆伊犁河谷常見的本土惡性入侵植物苦豆子幼苗早期生長(zhǎng)發(fā)育的影響,在實(shí)驗(yàn)室人工氣候箱盆播條件下,研究了不同程度子葉切除處理(切除0.5、2×0.5、1、1.5和2片子葉,以不切除子葉為對(duì)照)對(duì)苦豆子幼苗早期生長(zhǎng)的影響。結(jié)果表明,子葉切除的程度對(duì)幼苗的死亡率、株高和葉面積均影響顯著;高強(qiáng)度處理下,幼苗的主根長(zhǎng)與對(duì)照及低強(qiáng)度處理組間無(wú)顯著差異,而側(cè)根數(shù)和側(cè)根長(zhǎng)則均顯著低于對(duì)照組及低強(qiáng)度處理組。即使是低強(qiáng)度的子葉切除,苦豆子幼苗的莖、葉生物量以及幼苗的總生物量均會(huì)受到不同程度的抑制,而根生物量和根冠比受影響較小;子葉切除對(duì)幼苗各構(gòu)件的生物量分配的影響表現(xiàn)為:莖的生物量分配不受影響,根的顯著提高,而葉則受到不同程度的抑制。結(jié)果表明,子葉損傷是苦豆子幼苗建成的瓶頸。
苦豆子;幼苗;子葉損傷;生長(zhǎng)
幼苗階段是種子植物生長(zhǎng)最脆弱、易受外界環(huán)境影響的敏感階段,這主要緣于幼苗建成階段幼苗所積累的資源受限, 缺乏相應(yīng)的物理防御結(jié)構(gòu)或措施, 極易遭受動(dòng)物取食, 常見的如幼苗被去頭和子葉被取食等[1-2],極易導(dǎo)致種群更新受限甚至失敗。作為種子植物胚的組成部分之一的子葉,是植物發(fā)育時(shí)的第一片葉或第一對(duì)或第一輪葉中的一個(gè),在種子萌發(fā)時(shí)以及幼苗在長(zhǎng)出真葉能夠自養(yǎng)后的一段時(shí)間內(nèi),子葉在生理上仍起提供能量和營(yíng)養(yǎng)的作用,甚至充當(dāng)苗期的光合作用器官。無(wú)論幼苗萌發(fā)和定居的環(huán)境是否優(yōu)越,動(dòng)物取食或人為去除植物的子葉都將對(duì)幼苗的存活和生長(zhǎng)產(chǎn)生不利影響[3-8]。同時(shí),高強(qiáng)度的子葉去除對(duì)幼苗莖稈與根長(zhǎng)的生長(zhǎng)、各生長(zhǎng)構(gòu)件的干物質(zhì)積累以及內(nèi)含物的含量等均有較大影響[9-11],甚至降低植物幼苗在第2個(gè)生長(zhǎng)季節(jié)的繁殖能力和干物質(zhì)積累[12-13]。因此,開展子葉損傷或丟失的相關(guān)研究,對(duì)于植物的種群更新、群落演替以及農(nóng)業(yè)生產(chǎn)等均具有重要的理論及現(xiàn)實(shí)意義。
苦豆子(Sophoraalopecuroides)屬豆科槐屬中旱生、輕度耐鹽堿的多年生草本植物,主要分布于我國(guó)西北沙漠地區(qū)、華北地區(qū)以及中亞一帶[14-15]。由于苦豆子是干旱半干旱地區(qū)一種典型的無(wú)性系植物,通過無(wú)性繁殖可以有效地鞏固局部傳播和持久性,種子繁殖可以使物種傳播到較遠(yuǎn)的距離[16]。在新疆的伊犁河谷地區(qū),苦豆子在旱、濕和鹽堿生境均廣泛分布,河谷荒漠、農(nóng)田、濕地和草原生態(tài)系統(tǒng)均可形成以苦豆子為優(yōu)勢(shì)種或主要伴生種的草本植物群落。近10年來,由于氣候變化、人為活動(dòng)等原因,苦豆子在伊犁河谷山地草原的生態(tài)入侵步伐不斷加快,在多個(gè)縣境的草原上出現(xiàn)了連片分布景象,單個(gè)單優(yōu)群落面積可高達(dá)數(shù)十公頃[14],且種群向該地區(qū)更高海拔生境入侵的趨勢(shì)不斷加劇,給當(dāng)?shù)氐男竽翗I(yè)生產(chǎn)、生物多樣性和草原生態(tài)系統(tǒng)安全造成了嚴(yán)重威脅。
目前,對(duì)苦豆子生物生態(tài)學(xué)特性的研究大都集中在種子萌發(fā)生態(tài)學(xué)[16-19]、種間關(guān)系[20]等方面。有關(guān)苦豆子萌發(fā)直到幼苗初期生長(zhǎng)時(shí)影響因素的研究主要集中在鹽脅迫、干旱脅迫、土壤含水量、溫度等環(huán)境因素上[21-22],而有關(guān)苦豆子幼苗建成及其影響因素的研究還鮮有報(bào)道。在苦豆子的群落演替和種群擴(kuò)張過程中,種子具有重要作用,由于具有較高程度的硬實(shí)現(xiàn)象,在土壤微生物和土壤水分等的聯(lián)合作用下,部分種子可以在翌年春末至夏末萌發(fā),子葉中豐富的營(yíng)養(yǎng)物和光合產(chǎn)物可持續(xù)不斷地供給幼苗的生長(zhǎng),甚至與第一片復(fù)葉共存一段時(shí)間,確保了子葉部分丟失后幼苗的存活和生長(zhǎng)。由于放牧活動(dòng)和昆蟲采食是苦豆子苗期的主要擾動(dòng)者,采食或其他方式導(dǎo)致的子葉丟失,其損失程度對(duì)幼苗建成究竟產(chǎn)生什么樣的影響?因此,為了探明子葉損傷程度對(duì)苦豆子幼苗早期建成的影響,采用人工模擬手段切除子葉,研究幼苗死亡率、株高、葉面積、根長(zhǎng)、側(cè)根數(shù)、根冠比、各構(gòu)件生物量及生物量分配的影響,以期為苦豆子這一無(wú)性系植物補(bǔ)充更多的資料,也為新疆草場(chǎng)和牧區(qū)苦豆子的合理防控等提供科學(xué)理論依據(jù)。
1 材料與方法
1.1 材料
供試種采集自伊犁哈薩克自治州察布查爾錫伯自治縣境內(nèi)。察布查爾縣地處新疆西天山支脈烏孫山北麓,伊犁河以南(N 43°17′-43°57′,E 80°31′-81°43′)。縣域?qū)俅箨懶员睖貛睾透珊禋夂颍募痉置鳎韭L(zhǎng)、冬春長(zhǎng)、夏秋短,冬季寒冷,夏季炎熱,年均氣溫7.9 ℃;全年有效光照時(shí)數(shù)達(dá)2846 h,無(wú)霜期177 d,積溫3800 ℃,年平均降水量222 mm。縣域內(nèi)的山地草原、干旱荒漠、濕地和農(nóng)田生態(tài)系統(tǒng)均有大面積的苦豆子分布。
1.2 方法
2015年4月上旬,選取圓潤(rùn)、飽滿的苦豆子種子,采用機(jī)械破除的方法[19],避開種阜部位在種皮上切割一小缺刻,室溫下浸種24 h。盆播深度為1 cm,置于人工氣候箱內(nèi)培養(yǎng)(12 h光照,12 h黑暗,恒溫25 ℃)。待子葉出土張開60°后進(jìn)行子葉切除處理,分別為對(duì)照組(CK)和去除0.5片子葉(處理Ⅰ)、2×0.5片子葉(處理Ⅱ)、1片子葉(處理Ⅲ)、1.5片子葉(處理Ⅳ)和2片子葉(處理Ⅴ)的處理組,每個(gè)處理14個(gè)重復(fù),共90盆,每盆10棵苗。以6 d為一個(gè)觀察周期,每個(gè)周期統(tǒng)計(jì)一次幼苗的死亡情況和株高。經(jīng)過兩個(gè)月的培養(yǎng),于2015年5月下旬收獲各處理組的幼苗。用分析天平稱量各處理組苦豆子幼苗的根、莖和葉的鮮重,并用直尺測(cè)量幼苗的主根長(zhǎng)、株高、側(cè)根長(zhǎng)并計(jì)數(shù)側(cè)根數(shù),幼苗的葉面積用掃描儀(Canon LiDE 210)掃描后,用葉面積處理軟件(植物葉片分析軟件V 1.0)處理各處理組幼苗的葉面積。
1.3 統(tǒng)計(jì)分析
采用Excel 2003進(jìn)行數(shù)據(jù)統(tǒng)計(jì)分析,不同處理間的差異顯著性分析使用DPS (v7.05)中的單因子方差分析方法進(jìn)行。
2 結(jié)果與分析
2.1 子葉切除對(duì)幼苗存活的影響
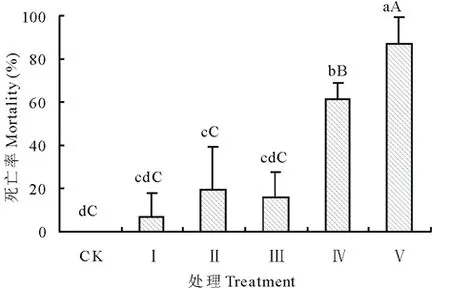
圖1 子葉切除對(duì)苦豆子幼苗死亡率的影響 Fig.1 Mortality of S. alopecuroides seedlings under different treatment of cotyledon excision 不同小寫字母表示不同處理間差異顯著(P<0.05),大寫字母表示差異極顯著(P<0.01)。下同。Different small letters indicate significantly different at the level of 0.05, and different capital letters indicate significantly different at the level of 0.01 between different treatment, the same below.
不同程度的子葉切除處理后,苦豆子幼苗的死亡率受到不同程度的影響,幼苗的死亡率與子葉切除的程度呈正相關(guān),結(jié)果如圖1所示。其中,CK、Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ處理組幼苗的死亡率分別為0、7.00%、19.50%、16.00%、61.34%和86.95%。同時(shí),處理Ⅰ與處理Ⅲ組間差異不顯著(P>0.05),兩處理組與CK間也均無(wú)顯著性差異(P>0.05);Ⅱ、Ⅳ和Ⅴ處理組與CK間均存在顯著性差異(P<0.05),處理Ⅰ、Ⅱ和Ⅲ間差異不顯著(P>0.05);CK、Ⅰ、Ⅱ、Ⅲ處理間均不存在極顯著差異(P>0.01),Ⅳ與Ⅴ間存在極顯著差異(P<0.01),且兩處理與對(duì)照及其他處理間也均存在極顯著差異(P<0.01)。結(jié)果顯示,幼苗的死亡率隨子葉丟失程度的增加而顯著增加。
2.2 子葉切除對(duì)幼苗生長(zhǎng)的影響
切除苦豆子出土子葉對(duì)幼苗的株高即莖的伸長(zhǎng)生長(zhǎng)具有一定的抑制效應(yīng),且抑制效應(yīng)隨子葉去除程度呈正相關(guān),各去除處理與對(duì)照間均存在極顯著差異(P<0.01),結(jié)果如表1所示。與對(duì)照相比,地下構(gòu)件主根的伸長(zhǎng)生長(zhǎng)受子葉去除程度的影響并不顯著,且各去除處理間也均不存在顯著性差異(P>0.05)。隨著子葉去除程度的增加,幼苗的側(cè)根數(shù)基本呈下降趨勢(shì),且高強(qiáng)度去除(處理Ⅳ和Ⅴ)與對(duì)照及其他處理間均存在極顯著(P<0.01)或顯著差異(P<0.05)。同時(shí),子葉損傷處理的Ⅰ、Ⅱ、Ⅲ、Ⅳ和Ⅴ幼苗側(cè)根的累積長(zhǎng)度均出現(xiàn)了不同程度的降低,且影響極顯著(P<0.01)。作為光合作用主器官的葉片,各子葉損傷處理組的幼苗葉面積與對(duì)照間也均存在極顯著的差異(P<0.01)。結(jié)果表明,除主根長(zhǎng)外,即使是低強(qiáng)度的子葉切除也會(huì)抑制苦豆子幼苗的生長(zhǎng),且抑制效應(yīng)與子葉的去除程度呈正相關(guān)關(guān)系。
2.3 子葉切除對(duì)幼苗生物量生長(zhǎng)的影響
由表2可以看出,子葉切除對(duì)苦豆子幼苗鮮生物量的影響具有構(gòu)件差異性。其中,根生物量及根冠比受子葉去除的影響較小,各去除處理與對(duì)照及各處理間均無(wú)顯著差異(P>0.05)。幼苗莖的生物量受子葉去除的影響較為顯著,Ⅱ、Ⅲ、Ⅳ和Ⅴ處理與CK間均存在極顯著差異(P<0.01),而Ⅱ、Ⅲ、Ⅳ、Ⅴ各處理間則差異不顯著(P>0.05)。受不同程度子葉去除的影響,葉生物量表現(xiàn)為CK>Ⅰ>Ⅱ>Ⅲ>Ⅳ>Ⅴ,處理Ⅳ與Ⅴ僅分別為CK的48.61%和26.44%,各去除處理與CK間均存在極顯著差異(P<0.01)。作為苦豆子復(fù)葉葉柄,其生物量受子葉去除的影響較小,各處理組與CK間均無(wú)顯著性差異(P>0.05)。幼苗的總生物量受子葉去除的影響也較為顯著,各處理與CK間均存在極顯著差異(P<0.01)。結(jié)果表明,子葉去除對(duì)苦豆子幼苗生物量的生長(zhǎng)具有不同程度的抑制效應(yīng),且存在構(gòu)件差異性。
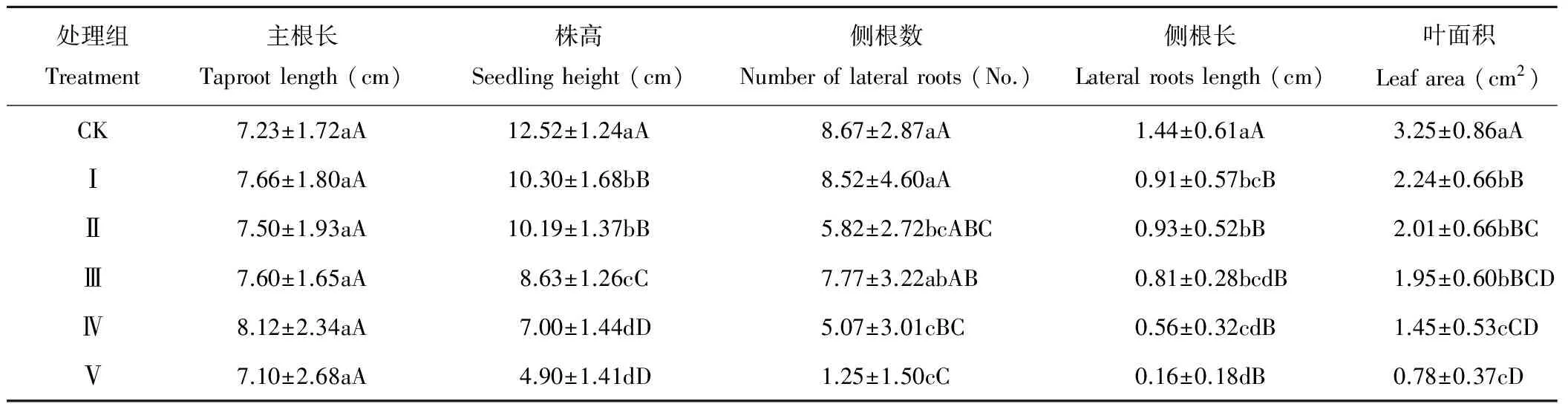
表1 不同子葉切除處理下苦豆子幼苗構(gòu)件生長(zhǎng)特征
注:同列不同小寫字母表示不同處理間差異顯著(P<0.05),不同大寫字母表示差異極顯著(P<0.01)。下同。
Note: Different small letters within the same column indicate significantly different at the level of 0.05, and different capital letters indicate significantly different at the level of 0.01, the same below.
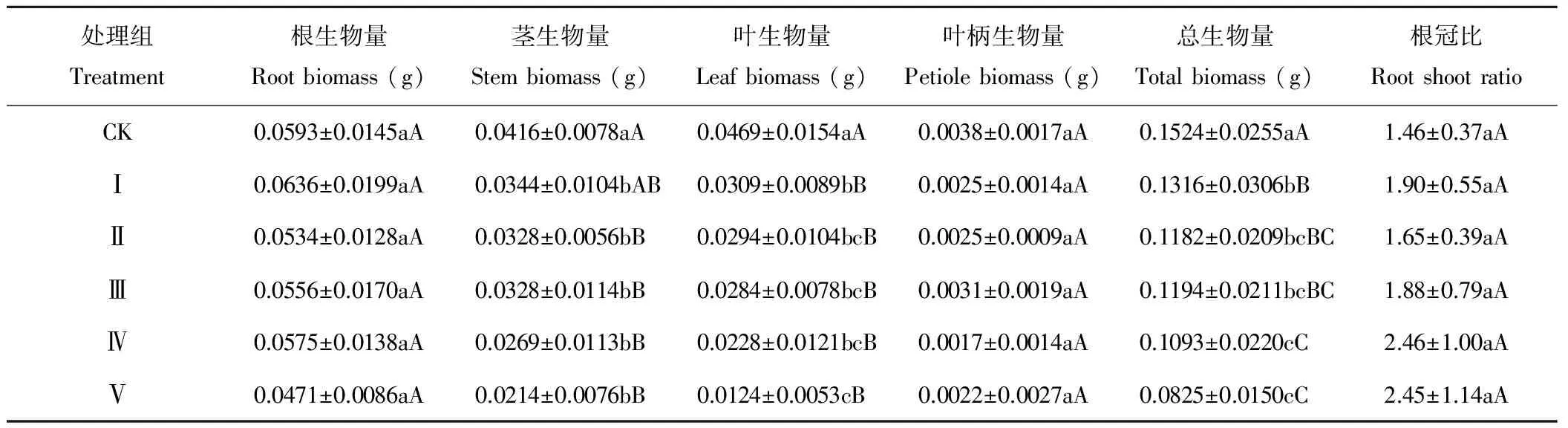
表2 子葉切除對(duì)苦豆子幼苗各構(gòu)件鮮生物量積累特征
2.4 子葉切除對(duì)苦豆子幼苗各構(gòu)件生物量分配的影響
在苦豆子幼苗各構(gòu)件的生物量結(jié)構(gòu)中,以地下根生物量分配為最高,其次為莖,葉構(gòu)件的最低(表3)。子葉去除可顯著增加幼苗根的生物量分配比例,處理Ⅱ、Ⅲ與Ⅳ和Ⅴ間存在顯著(P<0.05)或極顯著(P<0.01)差異,但與對(duì)照差異不顯著(P>0.05)。相較于地下的根,莖與葉構(gòu)成了幼苗的地上部,地上部生物量分配基本隨子葉去除程度的增加而降低,除處理Ⅱ外,各處理與對(duì)照間均存在顯著(P<0.05)或極顯著(P<0.01)差異,但各處理間差異不顯著(P>0.05)。對(duì)于地上部的莖,其生物量分配受子葉去除的影響程度較低, 各處理組與CK間差異不顯著(P>0.05)。葉生物量分配受子葉去除的影響則較為顯著,各處理組與對(duì)照間均存在極顯著差異(P<0.01),但處理間差異不顯著(P>0.05)。結(jié)果表明,即使是低強(qiáng)度的子葉切除,苦豆子幼苗的根構(gòu)件生物量分配顯著提高,而葉構(gòu)件生物量分配則受到不同程度的抑制。結(jié)果表明,子葉去除對(duì)苦豆子幼苗各構(gòu)件的生物量分配格局也具有一定的抑制效應(yīng),且存在構(gòu)件的差異性。
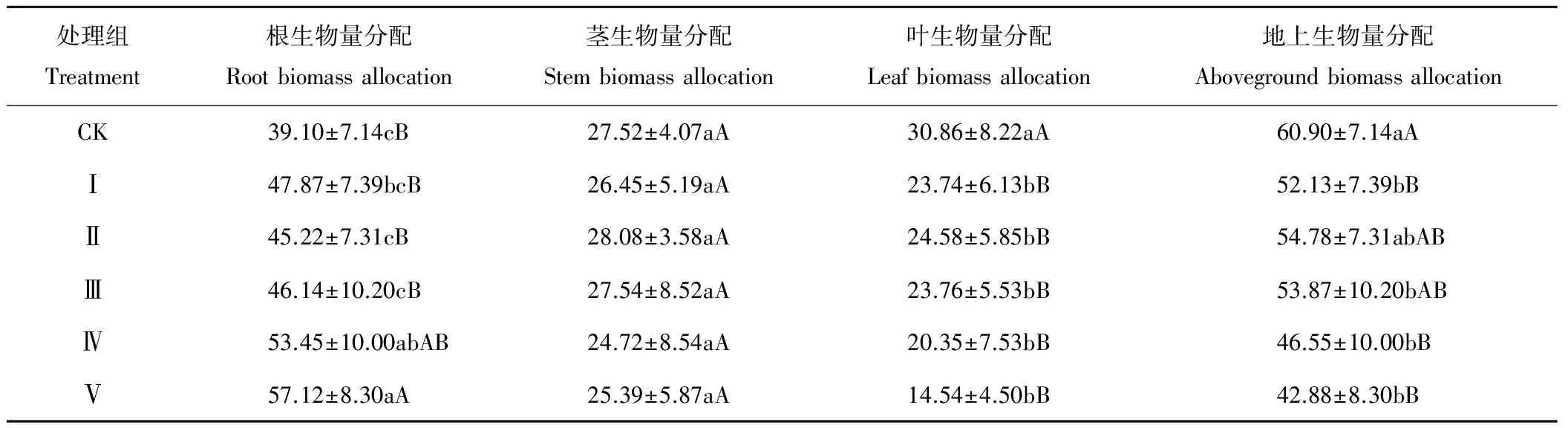
表3 子葉切除對(duì)苦豆子幼苗各構(gòu)件生物量分配特征
3 討論
子葉是豆科植物幼苗發(fā)育的主要營(yíng)養(yǎng)物質(zhì)來源,對(duì)種子萌發(fā)和幼苗建成均具有十分重要的作用。苦豆子種子在發(fā)育過程中及成熟后被豆莢螟取食危害的現(xiàn)象非常普遍[23],但苦豆子種子可以通過堅(jiān)硬種皮的機(jī)械防御及較高的種子硬實(shí)率而降低食種子動(dòng)物的取食壓力,留存的活力種子在種群的更新和新生境的拓展過程中發(fā)揮關(guān)鍵作用。作為一種雙子葉植物,苦豆子的子葉中貯藏著種子萌發(fā)和幼苗早期生長(zhǎng)所需的營(yíng)養(yǎng),并為幼苗抵御干旱脅迫并確保受損后生長(zhǎng)的恢復(fù)提供了物質(zhì)保障。同時(shí),子葉在種子萌發(fā)出土后變綠,成為幼苗生長(zhǎng)的第一個(gè)同化器官。已有的研究證明,子葉出土型種子萌發(fā)后,人為踐踏、動(dòng)物啃食或者病蟲害等均可使苦豆子幼苗的子葉受到損壞或全部丟失,幼苗生長(zhǎng)、物質(zhì)的同化與積累等均會(huì)受到不同程度的影響。因此,子葉的損傷可以影響干物質(zhì)的積累以及地上地下部分干物質(zhì)的分配[24],進(jìn)而影響幼苗的有效建成。本研究結(jié)果顯示,即使在種子萌發(fā)以后對(duì)苦豆子子葉進(jìn)行切除處理也會(huì)導(dǎo)致幼苗死亡,尤其在去除1.5和2片子葉的高強(qiáng)度處理下,幼苗的死亡率分別高達(dá)61.34%和86.95%,說明去除子葉阻斷了植株?duì)I養(yǎng)物質(zhì)的來源[25],進(jìn)而導(dǎo)致較高的幼苗死亡率,并導(dǎo)致苦豆子幼苗建成和種群更新的失敗。苦豆子種子萌發(fā)以后,幼苗生長(zhǎng)所需要的營(yíng)養(yǎng)物質(zhì)和能量主要依靠子葉的提供,如果子葉的丟失不高于1.5片,幼苗依靠?jī)H存的0.5片子葉依然可以保持50%以上的存活率并完成形態(tài)建成。
雙子葉植物間種子的大小存在一定的種屬差異,子葉儲(chǔ)存營(yíng)養(yǎng)物的量各不相同,導(dǎo)致各類子葉在動(dòng)物取食或其他原因?qū)е碌淖尤~部分或全部丟失對(duì)種子萌發(fā)、幼苗建成的影響也各不相同。對(duì)于體積較大的雙子葉植物如櫟屬植物,種子遠(yuǎn)大于多數(shù)草本植物的種子,子葉的部分損傷或丟失對(duì)種子萌發(fā)和幼苗建成并不會(huì)產(chǎn)生顯著的不良影響,這可能與大種子貯藏的營(yíng)養(yǎng)物多有關(guān)[10,16,25]。對(duì)于豆科的苦豆子,種子大小只有4.33 mm×3.21 mm×2.21 mm[26],屬于較小的種子。本研究結(jié)果發(fā)現(xiàn),無(wú)論中等強(qiáng)度或高強(qiáng)度子葉丟失對(duì)幼苗的根生物量及根冠比無(wú)影響,但對(duì)莖、葉生物量以及幼苗總生物量的抑制效應(yīng)顯著。黃瓜子葉比苦豆子的略大,子葉損傷對(duì)葉生物量的影響大于其他器官[27]。而對(duì)于種子更小的蕹菜,即使中等強(qiáng)度的子葉丟失,單株的根、莖、葉及整株的鮮重與干重等構(gòu)件數(shù)量和生物量指標(biāo)均顯著受到抑制[28]。由此說明,由于植物種屬造成的種子大小差異,子葉損傷對(duì)幼苗建成產(chǎn)生的影響存在一定的構(gòu)件差異性。
由于苦豆子種子的散布格局是聚集型散布,種子大多數(shù)散落于母株周圍,且多以莢果形式散布,豆莢及種皮較厚,種子不易破壞,被家畜啃食的種子經(jīng)過動(dòng)物消化道不易消化,還可以包裹于糞便中散往各處,在適宜的環(huán)境落地生根[16],所以苦豆子的子葉在萌發(fā)以前不易受到破壞。但苦豆子種子在春季萌發(fā)以后,人為踐踏、動(dòng)物啃食將會(huì)是子葉受損的最大因素,在干旱脅迫以及其他伴生種的競(jìng)爭(zhēng)下,殘留的子葉如何進(jìn)行營(yíng)養(yǎng)和能量分配也許會(huì)是在子葉受損初期幼苗能否存活并正常生長(zhǎng)的重要因素。研究結(jié)果發(fā)現(xiàn),去除2×0.5片子葉和去除1片子葉兩個(gè)處理組在幼苗株高生長(zhǎng)方面存在一定的差異性,生長(zhǎng)前期兩個(gè)處理組的株高數(shù)值差異不顯著,生長(zhǎng)后期則表現(xiàn)為去除2×0.5片子葉的幼苗株高顯著高于去除1片子葉,且差異達(dá)到了顯著水平,這種現(xiàn)象可能與子葉受損的部位也有關(guān),切除子葉的部位距胚根越近,可能對(duì)株高的伸長(zhǎng)生長(zhǎng)影響也越大[7]。在本研究中發(fā)現(xiàn),子葉受損對(duì)主根伸長(zhǎng)生長(zhǎng)的影響并不大,但對(duì)根生物量分配的影響很明顯,對(duì)側(cè)根數(shù)及其伸長(zhǎng)生長(zhǎng)有顯著抑制效應(yīng),說明子葉的貯存物在優(yōu)先供應(yīng)主根的伸長(zhǎng)生長(zhǎng),并在同時(shí)期萌生數(shù)量龐大的側(cè)根以汲取更多的營(yíng)養(yǎng)物質(zhì)和水分,保證子葉出土以后植株的正常生長(zhǎng)。
References:
[1] Hoshizaki K, Suzuki W, Sasaki S. Impacts of secondary seed dispersal and herbivory on seedling survival inAesculustrubinata. Journal of Vegetation Science, 1997, 8: 735-742.
[2] Hoshizaki K, Miguchi H, Forget P M,etal. Influence of forest composition on tree seed predation and rodent responses: a comparison of monodominant and mixed temperate forests in Japan[M]//Seed Fate: Predation, Dispersal and Seedling Establishment. Oxfordshire: CABI Publishing, 2005: 253-267.
[3] Schafer D E, Chilcote D O. Factors influencing persistence and depletion in buried seed populations. I. A model for analysis of parameters of buried seed persistence and depletion. Cropence, 1969, 9(4): 417-419.
[4] Vander Wall S B. A model of caching depth: Implications for scatter hoarders and plant dispersal. American Naturalist, 1993, 141(2): 217-232.
[5] Pizo M A, Oliveira P S. Size and lipid content of nonmyrmecochorous diaspores: effects on the interaction with litter-foraging ants in the Atlantic rain forest of Brazil. Plant Ecology, 2001, 157(1): 37-52.
[6] Lombardo J A, Mccarthy B C. Seed germination and seedling vigor of weevil-damaged acorns of red oak. Canadian Journal of Forest Research, 2009, 39(8): 1600-1605.
[7] Hou X, Yi X, Yang Y,etal. Acorn germination and seedling survival ofQ.variabilis: effects of cotyledon excision. Annals of Forest Science, 2010, 67(7): 711-717.
[8] Liang C L, Dai S H, Wu Y F,etal. Effect of injury to cotyledon on the later period growth and development ofVignaunguiculata. Crop Research, 2013, 27(2): 135-138. 梁成亮, 戴思慧, 吳藝飛, 等. 子葉損傷對(duì)豇豆生長(zhǎng)發(fā)育的影響. 作物研究, 2013, 27(2): 135-138.
[10] Fukumoto H, Kajimura H. Effects of insect predation on hypocotyl survival and germination success of matureQuercusvariabilisacorns. Journal of Forest Research, 2000, 5(5): 31-34.
[11] Liu Y, Li Q M, Liu Y,etal.Effects of mechanical treatments on seed germination ofQuercusfabriHance. Chinese Journal of Eco-Agriculture, 2013, 21(7): 867-871. 劉艷, 李慶梅, 劉勇, 等. 不同機(jī)械處理對(duì)白櫟種子萌發(fā)的影響. 中國(guó)生態(tài)農(nóng)業(yè)學(xué)報(bào), 2013, 21(7): 867-871.
[12] Andersson C, Frost I. Growth ofQuercusroburseedlings after experimental grazing and cotyledon removal. Acta Botanica Neerlandica, 1996, 45: 85-94.
[13] Giertych M J, Suszka J. Consequences of cutting off distal ends of cotyledons ofQuercusroburacorns before sowing. Annals of Forest Science, 2011, 68(2): 433-442.
[14] Lv D K, Ba Y S, Liu Y,etal.The effect of aqueous extract fromSophoraalopecuroidesseed on germination and seedling growth ofFestucaarundinacea. Xinjiang Agricultural Sciences, 2012, 49(8): 1477-1482. 呂篤康, 巴音山, 劉影, 等. 苦豆子浸出液對(duì)高羊茅種子萌發(fā)及幼苗生長(zhǎng)的影響. 新疆農(nóng)業(yè)科學(xué), 2012, 49(8): 1477-1482.
[15] Yang Y, Liu B R. Research progress on plant characteristics and resource utilization ofSophoraalopecuroides. Guizhou Agricultural Sciences, 2013, (12): 4-9. 楊陽(yáng), 劉秉儒. 苦豆子植物特性及資源化利用研究進(jìn)展. 貴州農(nóng)業(yè)科學(xué), 2013, (12): 4-9.
[16] Yang H, Hua P, Huang P Y. Study on the relationship between seed characteristics and population spreading ofSophoraalopecuroides. Journal of Arid Land Resources and Environment, 2006, 20(1): 198-201. 楊輝, 華鵬, 黃培祐. 苦豆子種子特性與種群擴(kuò)展關(guān)系的研究. 干旱區(qū)資源與環(huán)境, 2006, 20(1): 198-201.
[17] Zhang S R, Ji Y, Lin H M. Effects of nitrogen nutrition on dynamics of biological character and biomass accumulation ofSophoraalopecuraides. Pratacultural Science, 2008, 25(3): 37-42. 張守潤(rùn), 紀(jì)瑛, 藺海明. 施氮對(duì)苦豆子生物性狀和生物量積累動(dòng)態(tài)的響應(yīng). 草業(yè)科學(xué), 2008, 25(3): 37-42.
[18] Lv D K, Ouyang Y, Deng Y X. Terrestrial biomass and reproductive allocation characteristics ofSophoraalopecuroidesL. in Ili river valley. Xinjiang Agricultural Sciences, 2011, 48(7): 1333-1338. 呂篤康, 歐陽(yáng)艷, 鄧燕霞, 等. 伊犁苦豆子地上生物量及生殖分配特性研究. 新疆農(nóng)業(yè)科學(xué), 2011, 48(7): 1333-1338.
[19] Lv D K, Ba Y S, Zhao Y,etal. Seed germination characteristics ofSophoraalopecuroidesL. Acta Agrectir Sinica, 2011, 19(5): 889-892. 呂篤康, 巴音山, 趙玉, 等. 苦豆子種子萌發(fā)特性研究. 草地學(xué)報(bào), 2011, 19(5): 889-892.
[20] Chen L M G, Liu M S, Huang Z,etal. Distribution pattern of root biomass and inter-specific relationship inAchnatherumsplendens-Sophoraalopecuroidescommunity in northwest China.Chinese Journal of Plant Ecology, 2009, 33(4): 748-754. 晨樂木格, 劉茂松, 黃崢, 等. 我國(guó)西北地區(qū)芨芨草-苦豆子群落根系分布與種間關(guān)系. 植物生態(tài)學(xué)報(bào), 2009, 33(4): 748-754.
[21] Wang J, Han D H, Chen Y,etal. Influence of environmental factors on the seed germination and seedling growth ofSophoraalopecuroides. Acta Agrestia Sinica, 2007, 15(3): 259-262. 王進(jìn), 韓多紅, 陳葉, 等. 環(huán)境因子對(duì)苦豆子種子萌發(fā)和幼苗生長(zhǎng)的影響. 草地學(xué)報(bào), 2007, 15(3): 259-262.
[22] Si M Y·Ba L T, Yi L H M·Ai H M T, Ma L F. Effect of NaCl press on seed germination ofGlycyrrhizaglabraandSophoraalopecuroides. Seed, 2013, 32(1): 39-43. 司馬義·巴拉提, 伊力哈木·艾合買提, 馬劉峰. NaCl脅迫對(duì)光果甘草和苦豆子種子萌發(fā)的影響. 種子, 2013, 32(1): 39-43.
[23] Guo J J, Zhang D Z, Yang T T,etal. Spatial distribution pattern of seed pets ofSophoraalopecuroidesin desert area. Jiangsu Agricultural Sciences, 2012, 40(10): 111-113. 郭晶靜, 張大治, 楊婷婷, 等. 荒漠風(fēng)沙區(qū)苦豆子種子害蟲的空間分布型. 江蘇農(nóng)業(yè)科學(xué), 2012, 40(10): 111-113.
[24] Bonfil C. The effect of seed, cotyledon reserves, and herbivory on seedling survival and growth inQuercusrugosaandQ.laurina(Ragaceae). American Journal of Botany, 1998, 85(1): 79-87.
[25] Kitajima K. Impact of cotyledon and leaf removal on seedling survival in three tree species with contrasting cotyledon functions. Biotropica, 2009, 35: 429-434.
[26] Yi X F, Zhang Z B. Influence of insect-infested cotyledons on early seedling growth of Mongolian oak,Quercusmongolica. Photosynthetica, 2008, 46(1): 139-142.
[27] Jiang X F, Liu Q, Qiao F. Effect of cotyledon damage on the growth rate of cucumber seedlings. Chinese Journal of Tropical Crops, 2009, 30(3): 269-273. 江雪飛, 劉瓊, 喬飛. 子葉損傷對(duì)黃瓜幼苗生長(zhǎng)速率的影響. 熱帶作物學(xué)報(bào), 2009, 30(3): 269-273.
[28] Liu Y M, Peng J, Huang X F,etal. Effect of cutting the cotyledon on growth and development of water splnach seedling. Plant Physiology Communications, 1998, 34(6): 432-434. 劉義滿, 彭靜, 黃新芳, 等. 切除子葉對(duì)蕹菜幼苗生長(zhǎng)發(fā)育的影響. 植物生理學(xué)通訊, 1998, 34(6): 432-434.
Effect of cotyledon damage on the early growth ofSophoraalopecuroidesseedlings
LIU Ying1,2, ZHAO Yu1,2*, CUI Dong1, LENG Jia-Ming3, DONG Fang-Hui4
1.BiologyandGeographySchool,YiliNormalUniversity,Yining835000,China; 2.UniversityandCollegeKeyLabofNaturalProductChemistryandApplicationinXinjiang,Yining835000,China; 3.ChemistryandEnvironmentScienceSchool,YiliNormalUniversity,Yining835000,China; 4.HOPEAnalytechInc,Nanjing210007,China
The aim of these experiments was to determine the effects of cotyledon damage on the growth ofSophoraalopecuroides, a native invasive species in the Ili River Valley of Xinjiang. To simulate cotyledon predation, cotyledons were cut with different degrees of severity (excision of half a cotyledon, half of two cotyledons, one cotyledon, 1.5 cotyledons, and two cotyledons) or left uncut (control), and the growth of the seedlings was evaluated. The mortality rate, plant height, and leaf area strongly depended on the degree of cotyledon damage. There were no significant differences in seedlings’ taproot length among the damaged and non-damaged treatments. Compared with the control, the seedlings with 1.5 and two damaged cotyledons formed significantly fewer and shorter lateral roots, while those with half or one damaged cotyledon showed no significant difference in lateral root formation. Cotyledon damage significantly decreased the stem biomass, leaf biomass, and total biomass of seedlings, but had only small effects on the root biomass and root∶shoot ratio. As the severity of cotyledon damage increased, biomass allocation to the stem was unaffected, biomass allocation to the root significantly increased, and biomass allocation to the leaf significantly decreased. These results indicate that cotyledon damage is a bottleneck in seedling establishment and population regeneration.
Sophoraalopecuroides; seedling; cotyledon damage; growth
10.11686/cyxb2017052
http://cyxb.lzu.edu.cn
2017-02-17;改回日期:2017-04-19
國(guó)家自然科學(xué)基金項(xiàng)目(31360101)和新疆青年科技創(chuàng)新人才培養(yǎng)工程(qn2015yx021)資助。
劉影(1978-),女,安徽淮北人,高級(jí)實(shí)驗(yàn)師。E-mail:zylyzhlily@126.com*通信作者Corresponding author. E-mail: 2001zhaoyu@sohu.com
劉影, 趙玉, 崔東, 冷家明, 董芳慧. 子葉損傷對(duì)苦豆子幼苗早期生長(zhǎng)的影響. 草業(yè)學(xué)報(bào), 2017, 26(8): 139-145.
LIU Ying, ZHAO Yu, CUI Dong, LENG Jia-Ming, DONG Fang-Hui. Effect of cotyledon damage on the early growth ofSophoraalopecuroidesseedlings. Acta Prataculturae Sinica, 2017, 26(8): 139-145.
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
中學(xué)生數(shù)理化·八年級(jí)物理人教版(2022年3期)2022-03-16 05:55:08
當(dāng)代陜西(2021年2期)2021-03-29 07:41:24
小讀者(2021年2期)2021-03-29 05:03:48
華人時(shí)刊(2019年13期)2019-11-17 14:59:54
小天使·一年級(jí)語(yǔ)數(shù)英綜合(2019年8期)2019-08-27 02:23:00
文苑(2018年22期)2018-11-19 02:54:14
小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04
紅領(lǐng)巾·萌芽(2016年1期)2016-09-10 07:22:44
中國(guó)塑料(2016年3期)2016-06-15 20:30:00
