乳源酪蛋白糖巨肽對NF-κB信號通路中關鍵蛋白的調控作用
2017-09-09 05:37:26龔建苗龐廣昌閻亞麗陳慶森
食品科學 2017年17期
王 泳,龔建苗,賈 彥,趙 培,龐廣昌,閻亞麗*,陳慶森*
(天津市食品生物技術重點實驗室,天津商業大學生物技術與食品科學學院,天津 300134)
乳源酪蛋白糖巨肽對NF-κB信號通路中關鍵蛋白的調控作用
王 泳,龔建苗,賈 彥,趙 培,龐廣昌,閻亞麗*,陳慶森*
(天津市食品生物技術重點實驗室,天津商業大學生物技術與食品科學學院,天津 300134)
以結腸癌細胞HT-29為細胞系,在確定乳源性酪蛋白糖巨肽(casein glycomacropeptide,CGMP)對脂多糖(lipopolysaccharides,LPS)誘導的HT-29細胞核因子-κB(nuclear factor-κB,NF-κB)亞單位p65蛋白影響的基礎上,在最適作用時間條件下,利用Western blotting技術進一步檢測乳源CGMP對NF-κB信號通路上關鍵蛋白IκBα、p-IκBα、E3RSIκB、UBC5表達水平的影響,以闡述乳源CGMP調控NF-κB信號通路中關鍵蛋白的作用機制。結果表明:乳源CGMP組的3 種質量濃度(0.001、0.010、0.100 μg/mL)均可在一定程度上抑制LPS誘導的HT-29細胞NF-κB信號通路上IκBα蛋白的降解,0.100 μg/mL作用較為明顯,與空白對照組比較有顯著性差異(P<0.01)。研究明確地證實了乳源CGMP可通過抑制p-IκBα、E3RSIκB、UBC5蛋白的表達來抑制IκBα蛋白的降解,進而抑制NF-κB信號通路的激 活。結論:乳源CGMP可顯著降低NF-κB信號通路關鍵蛋白IκBα的降解,其機制是抑制了IκBα的磷酸化和泛素化,使p-IκBα和泛素化關鍵蛋白E3RSIκB和UBC5的表達均有所下降,進而減少了IκBα的降解,增加了IκBα-p65-p 50蛋白三聚體的數量,使p65蛋白核移位效應降低,進而減少下游基因的表達。因此,研究結果科學地闡釋了乳源CGMP是通過調控NF-κB信號通路發揮抗炎的作用。
酪蛋白糖巨肽;結腸癌細胞;核因子-κB;磷酸化核因子-κB抑制因子α;泛素-蛋白連接酶亞單位
王泳, 龔建苗, 賈彥, 等. 乳源酪蛋白糖巨肽對NF-κB信號通路中關鍵蛋白的調控作用[J]. 食品科學, 2017, 38(17): 26-31. DOI:10.7506/spkx1002-6630-201717005. http://www.spkx.net.cn
WANG Yong, GONG Jianmiao, JIA Yan, et al. Regulatory effect of milk-derived casein glycomacropeptide on key enzymes involved in NF-κB signaling pathway[J]. Food Science, 2017, 38(17): 26-31. (in Chinese with English abstract) DOI:10.7506/spkx1002-6630-201717005. http://www.spkx.net.cn
Janus激酶-信號轉導轉錄激活因子(janus kinasesignal transduction and transcription activator,JAK-STAT)、絲裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)及核因子-κB(nuclear factor κB,NF-κB)是細胞內3 條重要信號通路,對于維持細胞的正常生理功能具有重要意義[1-2]。其中NF-κB廣泛存在于真核生物細胞中,是與炎癥反應、免疫、腫瘤的發生與發展、細胞凋亡的調節以及胚胎發育等相關的信號傳導途徑的樞紐,是一種密切聯系這些信號途徑的重要核轉錄因子[3-4]。
NF-κB/Rel蛋白屬于Rel蛋白家族,以一定的方式兩兩結合成同源二聚體或異源二聚體[3,5-6]。據研究證明,所有的NF-κB復合物均與抑制因子IκB結合,在激活靶基因的轉錄[7]和在終止NF-κB信號中起著非常重要的作用[8]。在經典的NF-κB信號通路中,IKK是由IKKα、IKKβ和IKKγ組成的三亞基蛋白復合物,其中IKKγ是IKK復合物最后一個被發現的調節亞基 (也被稱作NEMO,NF-κB essential modulator);有研究指出,NEMO(48 kD)的結構含有一定的螺旋結構,C-末端也具有LZ,但NEMO序列的前50 個氨基酸殘基和后50 個氨基酸殘基并無二級結構。IKKα和IKKβ首先借助于分子中的亮氨酸拉鏈結構域形成二聚體,再分別通過C-末端一段短的結構域與NEMO結合形成穩定的三聚體[9-10]。研究顯示,NF-κB通過促進下游基因的轉錄而廣泛參與調控細胞凋亡[11-12]、細胞黏附[13]、細胞增殖、天然以及適應性免疫應答、炎癥[3,14-15]、胞內脅迫應答和組織重塑等過程。在諸如炎癥、腫瘤[16]、眼部疾病、神經損傷[17-18]、糖尿病[19]等疾病的發生發展過程中起著重要作用。
乳源性酪蛋白糖巨肽(casein glycomacropeptide,CGMP)源于牛乳中酪蛋白,是奶酪加工過程中釋放的一種由64 個氨基酸殘基組成的肽,該肽一半以上被糖基化修飾,故稱為“酪蛋白糖巨肽”[4,20-25]。本實驗室經過多年的研究,揭示了乳源CGMP是一種重要的乳源生物活性肽,并圍繞該肽開展了較為系統的研究工作取得了一些成果[26-31]。但乳源CGMP明確的調控NF-κB信號通路發揮其免疫調節作用的機制還有待進一步研究。本研究采用結腸癌細胞HT-29為細胞系,脂多糖(lipopolysaccharides,LPS)刺激干預,利用Western blotting測定乳源CGMP對腸上皮細胞NF-κB信號通路關鍵蛋白的影響,確定相應的作用靶點,以探討并闡述乳源CGMP調控NF-κB的分子機制,為確定乳源CGMP的抗炎分子機制以及維護相關腸黏膜內環境生物學活性功能提供科學依據,為乳源CGMP作為功能性食品用于炎癥性腸病的生物免疫治療提供參考。
1 材料與方法
1.1 材料與試劑
CGMP成品 新西蘭Tatua公司;人類結腸癌HT-29細胞 江蘇齊氏生物科技有限公司。
1.2 儀器與設備
倒置顯微鏡 日本 Nikon 公司;CO2培養箱 美國Thermo公司;多功能讀板機 美國Molecular Devices公司;迷你雙垂直電泳槽、迷你轉印電泳槽、電泳儀 北京六一儀器廠;高速冷凍離心機 美國Sigma公司。
1.3 方法
1.3.1 細胞分組及總蛋白質提取
人結腸癌細胞株HT-29細胞培養參考文獻[4]方法。
用無血清培養基同步化培養6 h后,將細胞分為5 組:空白對照組(N組)、LPS組(1 μg/mL,L組)、LPS(1 μg/mL)和乳源CGMP組(設計低(0.001 μg/mL,C1)、中(0.010 μg/mL,C2)、高(0.100 μg/mL,C3)3 個劑量)。各組處理參考文獻[3]方法及CGMP、LPS作用細胞的方法。按照107個細胞加入1 mL細胞總蛋白抽提試劑(含有蛋白酶、磷酸酶抑制劑),置于4 ℃搖床中速振蕩10 min,4 ℃條件下14 000 r/min離心15 min,收集上清液。并利用BCA方法定量測定總蛋白質含量。
1.3.2 Western blotting分析
乳源CGMP對LPS誘導HT-29細胞NF-κB信號通路中關鍵蛋白IκBα、p-IκBα、E3RSIκB和UBC5的Western blotting分析方法為:1)制備凝膠UBC5用質量分數15%聚丙烯酰胺分離膠;2)濕法轉膜:UBC5用迷你濕式轉印電泳槽200 mA恒流轉移1 h,將蛋白轉移至聚偏二氟乙烯膜;3)封閉:取下聚偏二氟乙烯膜,切掉右下角作標記后p-IκBα置于2%牛血清白蛋白中,室溫條件下搖床封閉1 h,主要封閉膜上的非特異結合部位。其他蛋白操作參考文獻[3]方法。
1.4 統計學分析
所有數據均做平行實驗,使用SPSS 17.0統計軟件進行分析處理,處理結果以±s表示,定義P<0.05為差異有統計學意義。
2 結果與分析
2.1 乳源CGMP對LPS誘導HT-29細胞IκBα蛋白及其磷酸化的影響
IκBα分子中均含有6 個錨蛋白(ankyrin)的重復序列,而這些重疊螺旋結構與p65分子中的RHR(rel homology region,RHR)區域結合,與p65的核定位序列(nuclear localization sequence,NLS)結合,使p65鈍化而滯留在胞質中。Arenzana-Seisdedos等[8]的研究證實了IκBα在終止p65活性的過程,確定了該活化的關鍵步驟是分子IκBα的磷酸化及降解。因此本研究選擇IκBα作為其中靶點之一,對其磷酸化前后蛋白含量變化進行觀測。
采用LPS激活打通結腸癌細胞HT-29的NF-κB信號通路后,接著給予乳源CGMP對LPS激活的HT-29細胞進行研究,探討對IκBα、p-IκBα蛋白的影響。較適劑量乳源CGMP作用48 h后,IκBα、p-IκBα蛋白的表達量見圖1、2。
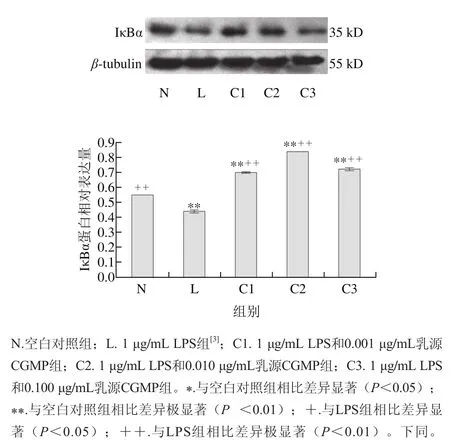
圖 1 乳源CGMP作用下IκBα蛋白的表達量變化Fig. 1 Effect of CGMP on the expression of IκBα
由圖1可知,與空白對照組相比,LPS組IκBα蛋白含量極顯著降低(P<0.01),降解極為明顯,LPS和乳源CGMP組IκBα蛋白表達先增加后降低,其中1 μg/mL LPS+ 0.010 μg/mL乳源CGMP抑制HT-29細胞IκBα蛋白的降解極為顯著(P<0.01),0.001、0.100 μg/mL乳源CGMP較0.010 μg/mL乳源CGMP抑制效果降低,但也極顯著抑制了IκBα蛋白的降解(P<0.01);與LPS組相比,1 μg/mL LPS分別與0.001、0.010、0.100 μg/mL乳源CGMP結合均極顯著增加了LPS刺激下IκBα蛋白的表達(P<0.01),其表達量極顯著高于空白對照組。
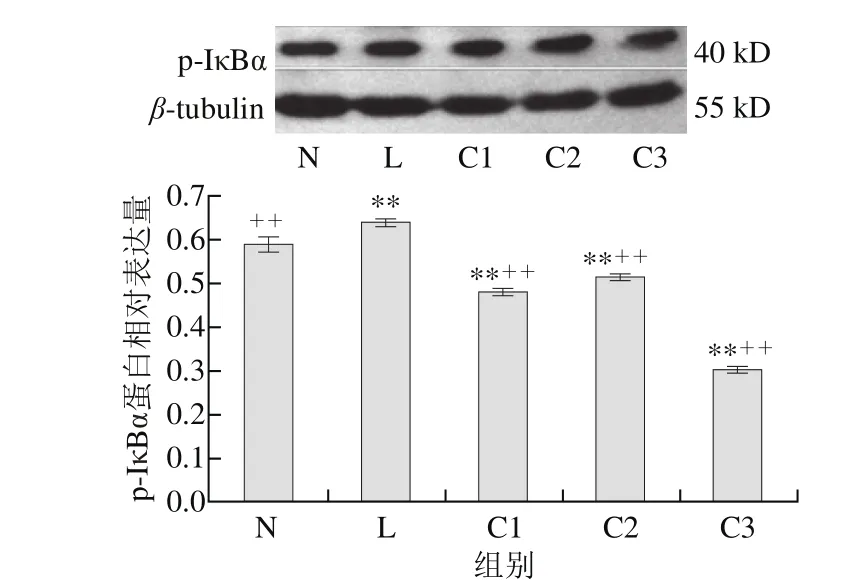
圖 2 乳源CGMP作用下p-IκBα蛋白的表達量變化Fig. 2 Effect of CGMP on the expression of p-IκBα
由圖2可知,與空白對照組相比,LPS組p-IκBα蛋白含量極顯著增加(P<0.01),1 μg/mL LPS+0.100 μg/mL乳源CGMP抑制HT-29細胞p-IκBα蛋白的表達極為顯著(P<0.01),較0.001、0.010 μg/mL乳源CGMP抑制效果好,但0.001、0.010 μg/mL乳源CGMP也極顯著抑制了p-IκBα蛋白的表達(P<0.01);與LPS組相比,0.001、0.010、0.100 μg/mL乳源CGMP均極顯著抑制了LPS刺激下p-IκBα蛋白的表達(P<0.01),其表達量極顯著低于空白對照組。
2.2 乳源CGMP對LPS誘導HT-29細胞IκBα降解泛素蛋白E3RSIκB和UBC5的影響
目前,研究揭示了泛素-蛋白酶體系統(ubiquitinproteasome pathway,UPP)[32-33]主要作用于細胞內一些半衰期短、結構異常、錯構或受損傷的蛋白,所以在調節信號通路中蛋白質降解與功能的作用越來越受到關注。因此,研究該通路途徑中泛素蛋白與乳源CGMP相互作用及其含量變化具有重要的意義,即泛素蛋白體系中相關蛋白表達水平成為人們研究NF-κB途徑的另一重要靶點。
采用LPS激活打通結腸癌細胞HT-29的NF-κB信號通路后,接著給予乳源CGMP對LPS激活的HT-29細胞進行研究,探討對E3RSIκB、UBC5蛋白的影響。較適劑量乳源CGMP作用48 h條件下,E3RSIκB和UBC5蛋白的表達量見圖3、4。
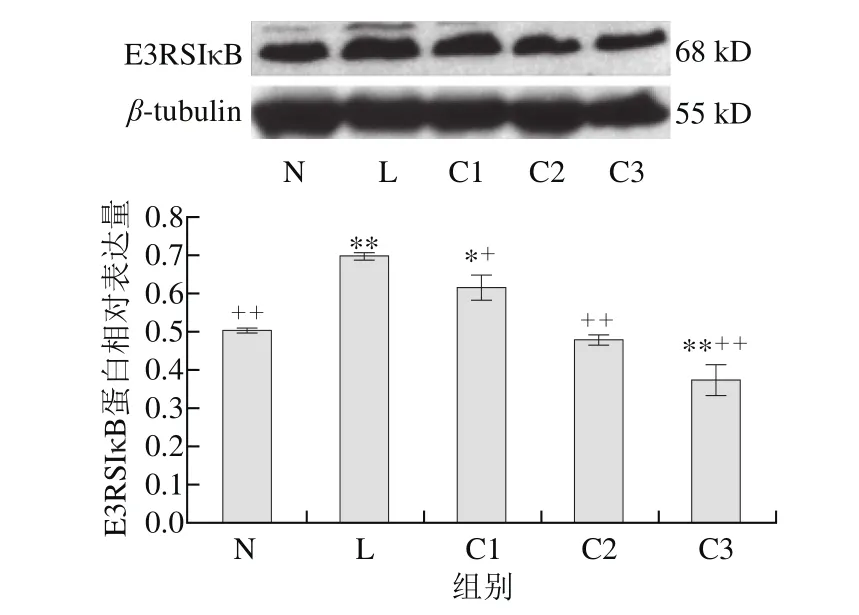
圖 3 乳源CGMP作用下E3RSIκB蛋白的表達量變化Fig. 3 Effect of CGMP on the expression of E3RSIκB
由圖3可知,與空白對照組相比,LPS組E3RSIκB蛋白表達量極顯著增加(P<0.01),LPS和乳源CGMP組E3RSIκB蛋白的表達量逐漸降低,且呈劑量依賴性,以1 μg/mL LPS+0.100 μg/mL乳源CGMP抑制E3RSIκB蛋白的表達最為顯著(P<0.01),C1抑制效果較低,仍顯著高于對照組(P<0.05),C2抑制效果也不明顯(P>0.05);與LPS組相比,C1、C2和C3顯著抑制了LPS刺激下E3RSIκB蛋白的表達(P<0.05,P<0.01),其中0.010、0.100 μg/mL乳源CGMP抑制效果較好(P<0.01)。
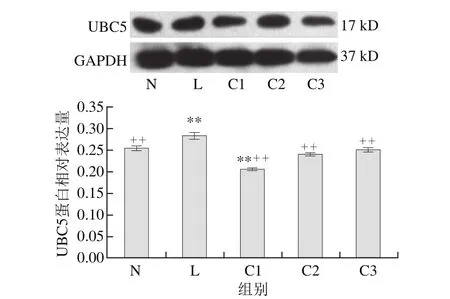
圖 4 乳源CGMP作用下UBC5蛋白的表達量變化Fig. 4 Effect of CGMP on the expression of UBC5
由圖4可知,與空白對照組相比,LPS組UBC5蛋白表達量極顯著增加(P<0.01),LPS和乳源CGMP組UBC5蛋白的表達量逐漸增加,且呈劑量依賴性,以0.001 μg/mL乳源CGMP抑制效果最為顯著(P<0.01),0.010、0.100 μg/mL乳源CGMP抑制效果不具顯著性(P>0.05);與LPS組相比,C1、C2和C3極顯著抑制了LPS刺激下UBC5蛋白的表達(P<0.01),其表達量極顯著低于LPS組。
3 討 論
當今,現代代謝綜合征的發展和蔓延,已成為嚴重的社會問題,越來越引起社會特別是科技界的重視,使人們談糖色變、談脂色變,而生物活性肽具有多種人體代謝和生理調節功能,易消化吸收;已有眾多的研究報道公布,由營養蛋白質釋放的生物活性肽可表現出多種生物學活性,如緩解和治療慢性炎癥、調節脂代謝、抵抗和排斥有害菌、抗病毒、降血壓、降血脂等作用;且食用安全性極高,重要的是在微量的狀態下就能發揮強大的生理活性,因此,是目前國際食品界最熱門的研究課題和極具發展前景的功能因子,從食源性的生物活性物質研究的角度,解決在相關疾病的預防及治療取得功效更是一條可行的途徑,乳源CGMP就是其中一個[34]。近期的研究已經揭示,生物活性肽發揮生理作用主要是通過和腸道內復雜的受體系統、微生物系統、代謝系統相互作用,通過改變自己的信號系統對機體發揮各種生理功能[35]。廣泛存在于機體細胞的NF-κB信號通路途徑能迅速對外界刺激產生反應,參與細胞凋亡、炎癥等生理條件下的基因調控而受到了越來越多的重視。此外,無論是非特異性免疫還是特異性免疫的應答均是通過各種各樣的信號傳遞實現的,因此,要研究乳源CGMP調控免疫發揮生理作用的機制,其干預免疫信號通路的研究是必不可少的。
科學研究表明NF-κB已成為疾病治療中的一個重要靶點,其激活依靠通路上的關鍵蛋白,倘若抑制某些必不可少的蛋白的活性,也會間接地抑制NF-κB的激活。因此,越來越多的研究選取NF-κB經典通路中必不可少的蛋白IκBα,對其磷酸化、泛素化相關蛋白進行測定。Rahman[36]發現具生物活性的谷胱甘肽可通過抑制IκB磷酸化而減少NF-κB活化,進而抑制相關細胞因子如TNF-α、細胞間黏附分子等的合成,而達到減少對細胞蛋白作用,緩解細胞炎癥狀態,或減少細胞凋亡。Fraternal等[37]的研究表明GSH-C4(巰基進行修飾)略微減少了p-IκBα和p65的表達。Gao等[38]用免疫共沉淀技術研究證實抗菌肽PR39(antibacterial peptide PR39)能可逆地結合到26S蛋白酶體的α7亞基上而抑制IκBα的降解,并沒有抑制IκBα的磷酸化和泛素化,且通過Western bloting技術證實這一結論。研究結果發現,以結腸癌細胞HT-29為細胞系,30 min的LPS刺激可以使IκBα的含量急劇下降,而乳源CGMP各組細胞IκBα含量均增加,且隨著乳源CGMP質量濃度的升高先升高后降低,但均極顯著高于空白對照組(P<0.01),以0.010 μg/mL作用效果最好,那么乳源CGMP阻止細胞IκBα的含量下降是通過刺激IκBα蛋白表達增加來實現的,亦或是通過抑制其磷酸化泛素化實現。p-IκBα蛋白表達的結果表明,LPS極顯著增加了p-IκBα蛋白的表達(P<0.01),而乳源CGMP的3 個質量濃度組均極顯著地抑制LPS刺激后的HT-29細胞p-IκBα蛋白的表達(P<0.01),作用效果明顯,不僅極顯著低于LPS組(P<0.01),更使p-IκBα蛋白的表達極顯著低于空白對照組(P<0.01),且以0.100 μg/mL的CGMP作用效果最好。因此,在某種程度上講,乳源CGMP減少了IκBα磷酸化也就是減少IκBα的降解,而是否促進IκBα的表達,還有待從基因表達水平上進行研究。
大量研究已證實,UPP在維持細胞內環境穩定方面具有關鍵調節作用,并且UPP對NF-κB通路的調控是在多個不同水平上進行的[33-34,39],也對各種疾病(如炎癥)的發生發展調控表現出該途徑的重要地位。因此,本研究中,為了證實乳源CGMP減少IκBα磷酸化而抑制IκBα的降解,這種抑制IκBα降解的作用是單純IκBα磷酸化減少的結果,還是也抑制了后續的泛素化?本研究測定了IκBα泛素化關鍵蛋白E3RSIκB和UBC5的表達。E3RSIκB分子中的WD40重復區域,是其與IκBα的結合位點,而E3RSIκB分子中的F-box負責誘導IκBα的泛素化[40]。本研究結果顯示,LPS刺激的HT-29細胞中E3RSIκB蛋白與空白對照組相比表達量極顯著增加(P<0.01),乳源CGMP降低了LPS刺激后E3RSIκB蛋白的表達,且呈劑量依賴關系,隨乳源CGMP劑量增加,抑制強度逐漸增大,以0.100 μg/mL的乳源CGMP作用效果最好(P<0.01),0.001 μg/mL的乳源CGMP雖然顯著降低了LPS刺激后E3RSIκB蛋白的表達,但仍顯著高于空白對照組,0.010 μg/mL乳源CGMP極顯著降低了LPS刺激后E3RSIκB蛋白的表達,但與空白對照組相比,不具有顯著性。在UBC5蛋白的測定中發現,LPS組UBC5蛋白與空白對照組相比表達量增加非常明顯,有極顯著差異(P<0.01),乳源CGMP同樣降低了LPS刺激后UBC5蛋白的表達,且呈劑量依賴關系,隨乳源CGMP劑量增加,抑制強度逐漸降低,以0.001 μg/mL的乳源CGMP作用效果最好,0.100、0.010 μg/mL的乳源CGMP雖然極顯著降低了LPS刺激后UBC5蛋白的表達,但與空白對照組相比,不具有顯著性。這些結果說明乳源CGMP可以通過UPP中關鍵酶的作用,抑制IκBα降解調控NF-κB信號通路,以發揮抗炎活性。
4 結 論
本研究指出,乳源CGMP具有顯著調節NF-κB信號通路,改善炎癥性腸病引發的結直腸癌的作用,其機制是通過顯著降低NF-κB信號通路關鍵蛋白IκBα的降解,抑制了IκBα的磷酸化和泛素化,使p-IκBα和泛素化關鍵蛋白E3RSIκB和UBC5的表達均有所下降,進而減少了IκBα的降解,增加了IκBα-p65-p50蛋白三聚體的數量,使p65蛋白核移位效應降低,進而減少下游基因的表達。研究結果較明確地闡釋了乳源CGMP是通過調控NF-κB信號通路以發揮抗炎活性的。
[1] 劉輝, 姚詠明. 細胞內炎癥信號通路交匯作用研究進展[J]. 感染、炎癥、修復, 2005, 4(8): 1607-1613. DOI:10.3969/j.issn.1672-8 521.2003.04.014.
[2] MA C F, WANG Y, DONG L, et al. Anti-inflammatory effect of resveratrol through the suppression of NF-κB and JAK/STAT signaling pathways[J]. 生物化學與生物物理學報(英文版), 2015(3): 207-213. DOI:10.1093/abbs/gmu135.
[3] GONG J M, CHEN Q S. Effect of casein glycomacropeptide on subunit p65 of nuclear transcription factor-κB in lipopolysaccharidestimulated human colorectal tumor HT-29 cells[J]. Food Science and Human Wellness, 2014, 3(2): 51-55. DOI:10.1016/j.fshw.2014.04.001.
[4] 龔建苗, 陳慶森, 閻亞麗, 等. 生物活性肽調節NF-κB信號通路的研究進展[J]. 食品科學, 2013, 34(21): 379-388. DOI:10.7506/s pkx1002-6630-201321075.
[5] GHOSH S, MAY M J, KOPP E B. NF-κB and Rel proteins: evolutionarily conserved mediators of immune responses[J]. Annual Review of Immunology, 1997, 16(1/2): 225-260. DOI:10.1146/ annurev.immunol.16.1.225.
[6] GILMORE T D. The Rel/NF-κB/IκB signal transduction pathway and cancer[J]. Spring US, 2004, 45(32): 221-228. DOI:10.1007/0-306-48158-8-10.
[7] KARIN M, BENNERIAH Y. Phosphorylation meets ubiquitination: the control of NF-κB activity[J]. Annual Review of Immunology, 2003, 18(1): 621-663. DOI:10.1146/annurev.immunol.18.1.621.
[8] ARENZANA-SEISDEDOS F, TURPIN P, RODRIGUEZ M, et al. Nuclear localization of IκBα promotes active transport of NF-κB from the nucleus to the cytoplasm[J]. Journal of Cell Science, 1997, 110 (Pt 3)(1): 369-378.
[9] LEE J, BURCKART G. Nuclear factor κB: important transcription factor and therapeutic target[J]. Journal of Clinical Pharmacology, 1998, 38(11): 981-993. DOI:10.1016/S0001-4575(01)00019-7.
[10] ZANDI E, ROTHWARF D M, DELHASE M, et al. The IκB kinase complex (IKK) contains two kinase subunits, IκKα and IκKβ, necessary for IκB phosphorylation and NF-κB activation[J]. Cell, 1997, 91(2): 243-252. DOI:10.1016/S0092-8674(00)80406-7.
[11] KUCHARCZAK J, SIMMONS M J, FAN Y, et al. To be, or not to be: NF-kappaB is the answer-role of Rel/NF-κB in the regulation of apoptosis[J]. Oncogene, 2003, 22(22): 8961-8982. DOI:10.1038/ sj.onc.1207230.
[12] DJAVAHERIMERGNY M, AMELOTTI M, MATHIEU J, et al. NF-κB activation represses tumor necrosis factor-alpha-induced autophagy[J]. Journal of Biological Chemistry, 2006, 281(41): 30373-30382. DOI:10.1074/jbc.M602097200.
[13] 崔嵩, 劉學鋒, 吳斌. NF-κB在腫瘤中的研究進展[J]. 現代腫瘤醫學, 2009, 17(1): 134-137.
[14] 鄭力新. NF-κB及其與淋巴細胞、細胞因子、病毒的基因調控[J].免疫學雜志, 1990(3): 147-152.
[15] CLEVERS H. At the crossroads of infi ammation and cancer[J]. Cell, 2004, 118(6): 671-674. DOI:10.1016/j.cell.2004.09.005.
[16] HUNOT S, BRUGG B, RICARD D, et al. Nuclear translocation of NF-κB is increased in dopaminergic neurons of patients with parkinson disease[J]. Proceedings of the National Academy of Sciences of the United States of America, 1997, 94(14): 7531-7536. DOI:10.1073/ pnas.94.14.7531.
[17] O’NEILL L, KALTSCHMIDT C. NF-κB: a crucial transcription factor for glial and neuronal cell function[J]. Trends in Neurosciences, 1997, 20(6): 252-258. DOI:10.1016/S0166-2236(96)01035-1.
[18] FLODSTR?MA M, WELSH N, EIZIRIK D L. Cytokines activate the nuclear factor κB (NF-κB) and induce nitric oxide production in human pancreatic islets[J]. Febs Letters, 1996, 385(1/2): 4-6. DOI:10.1016/0014-5793(96)00337-7.
[19] KWON G, CORBETT J A, HAUSER S, et al. Evidence for involvement of the proteasome complex (26S) and NF-κB in IL-1beta-induced nitric oxide and prostaglandin production by rat islets and RINm5F cells[J]. Diabetes, 1998, 47(4): 583-591. DOI:10.2337/ diabetes.47.4.583.
[20] DELFOUR A, JOLLèS J, ALAIS C, et al. Caseino-glycopeptides: characterization of a methionine residue and of the N-terminal sequence[J]. Biochemical & Biophysical Research Communications, 1965, 19(4): 452-455. DOI:10.1016/0006-291X(65)90145-2.
[21] BRODY E P. Biological activities of bovine glycomacropeptide[J]. Food Science and Technology, 2000, 84(5): 39-46.
[22] JIMENEZ H F J R. Nomenclature of the proteins of cows’ milk: sixth revision[J]. Journal of Dairy Science, 2004, 87(6): 1641-1674. DOI:10.3168/jds.S0022-0302(04)73319-6.
[23] FIAT A M, JOLLèS P. Caseins of various origins and biologically active casein peptides and oligosaccharides: structural and physiological aspects[J]. Molecular & Cellular Biochemistry, 1989, 87(1): 5-30. DOI:10.1007/BF00421079.
[24] PISANO A, PACKER N H, REDMOND J W, et al. Characterization of O-linked glycosylation motifs in the glycopeptide domain of bovine kappa-casein[J]. Glycobiology, 1994, 4(6): 837-844. DOI:10.1093/ glycob/4.6.837.
[25] ZEVACO C, RIBADEAU D B. A study on the carbohydrate binding sites of bovine beta-casein using high performance liquid chromatography[J]. Milchwissenschaft-milk Science International, 1984, 39(4): 206-210.
[26] 李博智, 陳慶森. 利用TGase結合微濾技術從乳清粉中分離純化GMP的研究[J]. 食品科學, 2009, 30(20): 97-103.
[27] 曹晉宜, 陳慶森. 瑞士乳桿菌對小鼠腸道粘膜和腸組織中細胞因子影響的研究[J]. 食品科學, 2009, 30(21): 343-347. DOI:10.3321/ j.issn:1002-6630.2009.21.079.
[28] 李偉, 陳慶森. 酪蛋白糖巨肽對小鼠腸道免疫系統的影響[J]. 食品科學, 2010, 31(15): 240-243.
[29] CHEN Q S, WANG H, ZHU C C, et al. Anti-apoptotic effects of milkderived casein glycomacropeptide on mice with ulcerative colitis[J]. Food and Agricultural Immunology, 2014, 25(4): 453-466. DOI:10.10 80/09540105.2013.823912.
[30] 龐廣昌, 陳慶森, 胡志和. 食品是如何通過細胞因子網絡控制人類健康的(Ⅰ)[J]. 食品科學, 2006, 27(6): 258-264. DOI:10.3321/ j.issn:1002-6630.2006.05.058.
[31] MING Z, JIA Y, YAN Y L, et al. Amelioration effect of bovine casein glycomacropeptide on ulcerative colitis in mice[J]. Food & Agricultural Immunology, 2015, 26(5): 1-12. DOI:10.1080/09540105. 2015.1018874.
[32] ESTEBAN V, RRPEREZ M, SANCHEZLOPEZ E, et al. Angiotensin IV activates the nuclear transcription factor-kappaB and related proinflammatory genes in vaseular smooth musele cells[J]. Circulation Researeh, 2005, 96(9): 965-973. DOI:10.1161/01.RES. 0000166326.91395.74.
[33] BRUINSMA C. The role of UBE2A and UBE3A in motor- and cognitive function[D]. Rotterdam: Erasmus University Rotterdam, 2016: 12.
[34] GIESSEN T W, MARAHIEL M A. Ribosome-independent biosynthesis of biologically active peptides: application of synthetic biology to generate structural diversity[M]. Harvard: Harvard Business School Press, 1990: 2065-2075. DOI:10.1016/j.febslet.2012.01.017.
[35] 龐廣昌, 陳慶森, 胡志和, 等. 蛋白質的消化吸收及其功能評述[J].食品科學, 2013, 34(9): 375-391. DOI:10.7506/spkx1002-6630-201309074.
[36] RAHMAN I. Regulation of nuclear factor-κB, activator protein-1, and glutathione levels by tumor necrosis factor-alpha and dexamethasone in alveolar epithelial cells [J]. Biochemical Pharmacology, 2000, 60(8): 1041-1049. DOI:10.1016/S0006-2952(00) 00392-0.
[37] FRATERNALE A, CRINELLI R, CASABIANCA A, et al. Molecules altering the intracellular thiol content modulate NF-κB and STAT-1/ IRF-1 signalling pathways and IL-12 p40 and IL-27 p28 production in murine macrophages[J]. PLoS ONE, 2013, 8(3): e57866.DOI:10.1371/ journal.pone.0057866.
[38] GAO Y, LECKER S, POST M J, et al. Inhibition of ubiquitin-proteasome pathway-mediated IκBα degradation by a naturally occurring antibacterial peptide[J]. Journal of Clinical Investigation, 2000, 106(106): 439-448. DOI:10.1172/JCI9826.
[39] KRAPPMANN D, SCHEIDEREIT C. A pervasive role of ubiquitin conjugation in activation and termination of IκB kinase pathways[J]. Embo Reports, 2005, 6(4): 321-326. DOI:10.1038/sj.embor.7400380.
[40] MARGOTTIN F, BOUR S P, DURAND H, et al. A novel human WD protein, h-βTrCP, that interacts with HIV-1 Vpu connects CD4 to the ER degradation pathway through an F-Box motif[J]. Molecular Cell, 1998, 1(4): 565-574. DOI:10.1016/S1097-2765(00)800 56-8.
Regulatory Effect of Milk-Derived Casein Glycomacropeptide on Key Enzymes Involved in NF-κB Signaling Pathway
WANG Yong, GONG Jianmiao, JIA Yan, ZHAO Pei, PANG Guangchang, YAN Yali*, CHEN Qingsen*
(Tianjin Key Laboratory of Food Biotechnology, College of Biotechnology and Food Science, Tianjin Un iversity of Commerce, Tianjin 300134, China)
This study was undertaken to determine the effect of casein glycomacropeptide (CGMP) on the p65 subunit of nuclear factor-κB (NF-κB) of HT-29 human colon cancer cells challenged with lipopolysaccharide (LPS). Furthermore, Western blotting technology was used to detect and compared the expression levels of the key proteins involved in the NF-κB signaling pathway such as IκBα, p-IκBα, E3RSIκB and UBC5 in the control group, milk-derived CGMP group and LPS group to uncover the molecular mechanism of CGMP in regulating the NF-κB signaling pathway. The results showed that all three doses of CGMP (0.001, 0.010 and 0.100 μg/mL) could suppress the degradation of IκBα in some degree, which is involved in the NF-κB signaling pathway of HT-29 cells challenged with LPS and this effect was more significant at the dose of 0.100 μg/mL, which was significantly different when compared with the control group (P < 0.01). Therefore, it was confi rmed that CGMP could suppress the degradation of IκBα by repressing the expression of p-IκBα, E3RSIκB and UBC5, thereby inhibiting the activation of the NF-κB signaling pathway. As a result, CGMP can signifi cantly reduce the degradation of IκBα in the NF-κB signal pathway, increase the amount of protein trimer-IκBα-p65-p50 and decrease nuclear translocation of p65 and downstream gene expression by inhibiting the phosphorylation and ubiquitination of IκBα and reducing the expression of p-IκBα and key ubiquitin proteins (E3RSIκB and UBC5). This study illustrates that milk-derived CGMP plays an anti-infi ammatory role by regulating the NF-κB signal pathway.
casein glycomacropeptide; colon cancer cells (HT-29); nuclear factor-κB; IκBα; ubiquitin-protein ligase subunit
10.7506/spkx1002-6630-201717005
R151
A
1002-6630(2017)17-0026-06引文格式:
2016-09-28
國家自然科學基金面上項目(31071522)
王泳(1993—),女,碩士研究生,研究方向為發酵生物技術、功能成分與腸道健康的關系。E-mail:15620616749@163.com *通信作者:閻亞麗(1962—),女,副教授,碩士,研究方向為發酵生物技術、微生物學。E-mail:yyali@tjcu.edu.cn陳慶森(1957—),男,教授,博士,研究方向為發酵生物技術、功能成分與腸道健康的關系。E-mail:chqsen@tjcu.edu.cn
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
中國生殖健康(2019年3期)2019-02-01 06:12:26
汽車工程學報(2017年2期)2017-07-05 08:13:02
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
