廣西甘蔗田馬唐對莠去津抗性水平檢測及機理初探
2017-09-15 07:32:05王彥輝楊彩英馬永林黃輝曄郭成林覃建林馬躍峰
西南農業學報 2017年8期
王彥輝,楊彩英,馬永林,黃輝曄,郭成林,覃建林,馬躍峰
廣西甘蔗田馬唐對莠去津抗性水平檢測及機理初探
王彥輝1,2,楊彩英1,馬永林1,2,黃輝曄1,2,郭成林1,2,覃建林1,2,馬躍峰1,2
(1.廣西壯族自治區農業科學院植物保護研究所,廣西南寧530007;2.廣西作物病蟲害生物學重點實驗室,廣西南寧 530007)
【目的】開展廣西甘蔗田間雜草馬唐(Digitaria sanguinalis)對莠去津的抗藥性及抗性機理的研究,為其科學防除和抗性治理提供參考。【方法】采用盆栽試驗,利用整株法測定采集于廣西蔗區的25個馬唐種群對莠去津的抗性水平;采用生理生化和分子生物學等方法分析研究馬唐抗莠去津的機理。【結果】供試馬唐種群中76%對莠去津產生抗性,其中貴港1號馬唐種群相對抗性水平最高,抗性指數為37.2。崇左4號為敏感品系,百色1號和柳州2號種群3 d后GSTs相對活力明顯高于敏感種群;對葉綠素上的D-1蛋白的編碼基因psbA基因進行測序,發現抗性種群psbA基因均沒有發現突變,GSTs的活性增強與馬唐對莠去津的抗性相關。【結論】廣西甘蔗種植區馬唐對莠去津抗性水平為低到中抗水平,GSTs酶活性的增強可能是產生抗性的主要原因之一。
馬唐;莠去津;抗性;GSTs;psbA
【研究意義】莠去津(Arazine)為三氮苯類除草劑,其作用機理是通過改變膜蛋白的空間構型,干擾雜草的光合作用,導致雜草死亡,在我國主要用于甜高粱、甘蔗、高粱、玉米等作物田間除草。目前該類除草劑已成為產生抗性最為嚴重的除草劑之一[1]。馬唐(Digitaria sanguinalis)是我國分布廣泛的一種的農田雜草,主要危害玉米、豆類、棉花、花生、甘蔗、蔬菜和果樹等作物。在廣西地區甘蔗田中常年(使用10年以上)使用莠去津進行防除,單一使用該藥劑已經造成田間防效下降,因此,研究馬唐對莠去津的抗藥性機制,對合理防控雜草,延緩具有除草劑的使用年限具有重要意義。【前人研究進展】廣西蔗區常用的除草劑使用較為單一,其中芽前除草劑主要為莠去津和乙草胺。除草劑莠去津在甘蔗地使用量大和年限長,對抗藥性研究較少。1970年Ryan等首次報道了千里光對光系統Ⅱ(PSⅡ)抑制劑莠去津產生抗藥性,目前已經超過了80多種雜草生物型對PSⅡ抑制劑(三唑酮、尿嘧啶和其他三氮苯除草劑)產生抗藥性[1-2]。有關研究認為雜草抗PSⅡ抑制劑的機理主要是D-1蛋白psbA基因突變(靶標位點)和代謝能力增強(非靶標位點)[3-5]。大多數雜草對PSⅡ抑制劑的抗性是除草劑靶標位點或者靶標蛋白(D-1蛋白)改變、親和性下降引起的。Hirschberg和McIntosh首先報道了莧菜對莠去津的抗性機理,發現類囊體膜的D-1蛋白的228位上的一個絲氨酸被甘氨酸所取代,導致抗性的產生[3]。三氮苯類除草劑通過氫鍵與Ser264的羥基和Phe265的氨基于氨基酸在PQ結合相互作用,同時與Phe255的苯環產生疏水反應。氨基酸Ser264被Gly取代后三氮苯類除草劑在D-1蛋白上結合的氫鍵被移除,導致除草劑對D-1蛋白的親和力下降,從而降低除草劑的生物活性或產生抗性[6]。該突變也是造成其他PSⅡ抑制劑抗性原因,較多研究結果表明其為雜草對PSⅡ抑制劑三氮苯類除草劑產生抗藥性的分子機理[7-9]。【本研究切入點】目前對廣西甘蔗田馬唐對莠去津的抗性水平和抗性機理報道較少。【擬解決的關鍵問題】以廣西甘蔗區馬唐種群為研究對象,檢測其對莠去津的抗性水平,并對抗性機理進行初探。厘清廣西甘蔗田馬唐抗性水平,初步探明研究馬唐對莠去津的抗性機理,為指導科學合理用藥和抗PSⅡ抑制劑類雜草的治理提供實驗證據。
1 材料與方法
1.1 供試材料
于2014-2015年采集與廣西14個縣的甘蔗產區,直接采集成熟的馬唐種子;在室內自然風干后,放入250 mL的棕色玻璃瓶中,置于種子儲存柜中保存。供試儀器設備為:種子存儲柜(杭州托普儀器有限公司);光照培養箱(寧波賽福有限公司); 3WPSH-500D型生測噴霧塔(農業部南京農業機械化研究所制);UV-1240型紫外分光光度計(日本島津公司);Z326K高速冷凍離心機(德國hemmer公司);梯度PCR儀(德國耶拿分析儀器股份公司)。供試試劑包括:80%敵草隆水分散粒劑(以色列馬克西姆阿甘工業公司);谷胱甘肽S-轉移酶(Glutathione-S-transferase,GSTs)酶活性測定試劑盒(南京建成公司);植物DNA提取試劑盒(Omega Biotek公司);Taq PCR StarMix試劑(北京康潤誠業生物科技有限公司)等。
1.2 試驗方法
1.2.1 整株植物測定法[10]在溫室中將催芽后的30粒種子,均勻播種在直徑為12 cm(直徑)×10 cm(深)的塑料盆中,播種后馬唐種子覆蓋一層薄土,保證出苗率。試驗時期溫度為25~35℃。待植株長至2~3葉期間苗,保留長勢基本一致的植株10株,3~4葉期間噴藥處理。根據預實驗結果用配置一系列濃度,0、0.3125、0.625、1.250、2. 500、5.000、10.000和20.000 g/L。每個處理3次重復,14 d后剪取地上部分,稱量鮮物質質量,數據采用DPS統計分析軟件處理,計算抑制中量IC50。
1.2.2 GSTs活性測定植株前處理同 1.2.3的方法,選擇不同抗性種群進行GST酶活性研究,在3~4葉期噴濃度為1.25 g/L的莠去津,藥后0、0.25、1、3、5、7 d采集馬唐葉片,立即冷凍到-80℃冰箱中待用;每個處理3個重復。莠去津處理后GSTs酶液提取參照吳進才等的方法。稱取采集的莠去津樣品0.5 g,剪碎放入預冷后的研缽中,加入適量液氮快速研磨成細粉,再加入8 mL Tris-HCl緩沖液(0.1 mol/L,pH 8.0,含還原型谷胱甘肽25mmol/L,5%聚乙烯吡咯烷酮),研磨勻漿,取上清液于14 000× g再離心10 min,上清液即為粗酶液。GSTs活性測定采用南寧建成有限公司生產的試劑盒。蛋白質的含量測定采用Bradford的考馬斯亮藍G-250法,以牛血清蛋白作標準曲線。
1.2.3 靶標基因突變檢測 在NCBI上下載馬唐psbA基因序列(KM453691.1)以及其他同源性高的psbA基因序列,設計上游引物為F1(5′-ATGACTGCAATTTTAGAGAGACGCGAAAG-3′),下游引物R3 (5′-AATAGGTAATCCTCTACCTTGAAGTTCTCGTCG-3′);葉片在4~5葉期收集,對psbA全長進行PCR擴增。馬唐DNA提取采用Omega的植物提取試劑盒,按照說明書操作。總反應體系為20μl;正向引物與反向引物各1μl,模板DNA 1μl,Taq PCR Star-Max 10μl,ddH2O補足至20μl,PCR反應條件為:預變性95℃,5 min;35循環:變性95℃,30 s;退火55℃,1 min;延伸72℃,1min;最終延伸72℃,10 min;4℃保存。PCR反應結束后,取5μl PCR產物,1.5%瓊脂糖凝膠電泳檢測是否有目的條帶,然后進行膠回收測序。
2 結果與分析
2.1 馬唐對莠去津的抗性水平
從測定結果(表1)可以看出,不同馬唐生物型對莠去津的抗性水平差異較大,所測25個馬唐生物型對莠去津的抗性水平差異較大,以崇左4號為最敏感種群,其中5個生物型(河池1號、來賓2號、崇左1號、崇左3號、百色2號等)對莠去津較敏感;5個生物型(南寧1號、崇左2號、貴港3號、百色1號、崇左5號)抗藥性水平較低,抗藥性指數為2~10;其余15個生物型對其抗性為中等水平,其中崇左1號抗藥性指數最高,為37.2,南寧2號和南寧3號對莠去津抗藥性指數也在30及上。

表1 馬唐對莠去津抗性水平測定Table 1 The resistance level of different Digitaria sanguinalis populations to atrazine
2.2 莠去津對馬唐GSTs活性的影響
以崇左4號敏感種群,分別選取抗性種群百色1號、柳州2號和貴港1號測定GSTs活性,研究GSTs活性與抗性的關系。因不同的種群GSTs酶活性本底不同,本研究采用相對活性研究GSTs的變化。經莠去津1.25 g/L處理后,所有種群GSTs相對活力開始上升,敏感種群在第1天達到最大值,是對照的1.64倍,然后開始下降,第5天之后趨于平緩。柳州2號和百色1號種群經藥劑處理后,在第3 d達到最高值,為對照的1.73和1.55倍,之后下降,在第3天后的相對活性均高于敏感種群。然而貴港種群在藥劑處理后GSTs相對活性一直低于敏感種群和另外兩個種群,這說明除GSTs酶參與抗性,可能還有其他抗性機制的存在(圖1)。
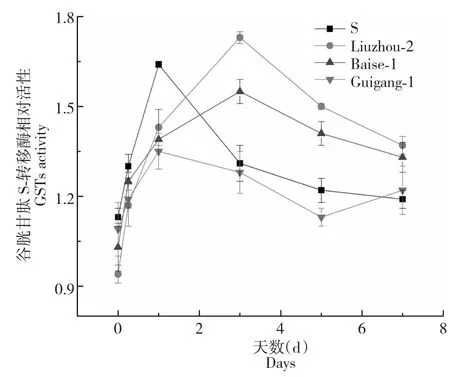
圖1 抗性及敏感馬唐種群莖葉組織GSTs活力變化Fig.1 The activity variation of GSTs in resistant and susceptible Digitaria sanguinalis
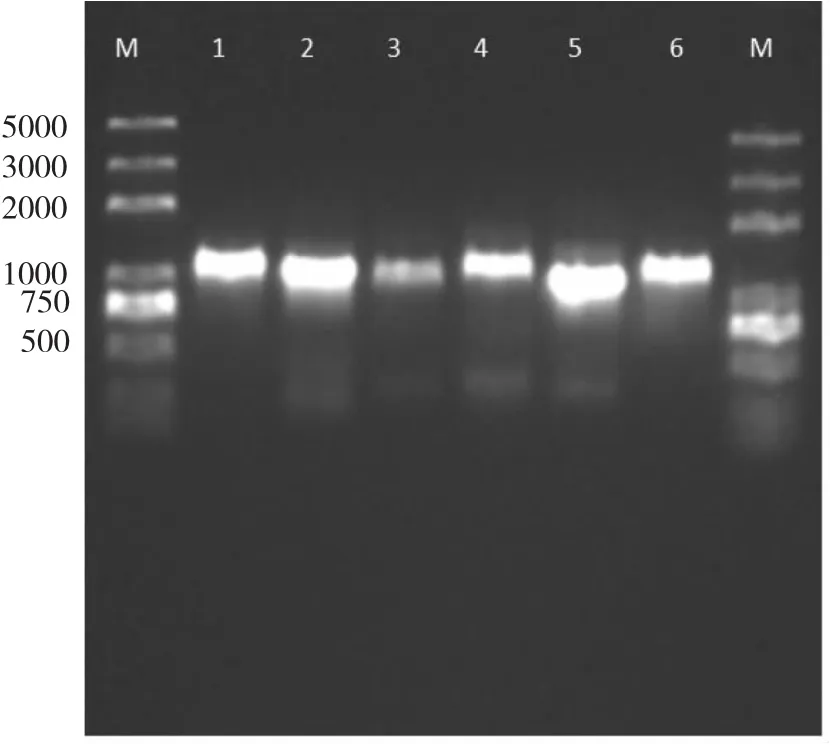
圖2 馬唐psbA基因DNA凝膠電泳檢測Fig.2 Identification of psbA gene from Digitaria sanguinalis on 1.0 %agarose gel
2.3 靶標抗性檢測
根據設計的引物進行PCR擴增,能很好的擴增出1000 bp左右的基因,見圖2。通過對測序結果進行比對分析與NCBI提交的psbA基因序列相似度高于90%,證明該引物能很好的擴增出psbA基因。與敏感性種群對比,抗性種群psbA基因并沒有發生突變,說明馬唐對莠去津的抗性機制很可能是非靶標抗性。
3 討 論
廣西是我國主要的甘蔗生產地之一,莠去津因除草效果好、性價比高,受到農民的歡迎。然而使用年限較長,田間藥效試驗發現對馬唐的防除效果明顯下降的現象。周青等[11]通過田間和室內生物測定,發現河南安陽地區玉米田的馬唐對阿特拉津、乙阿合劑產生抗性,田間用藥劑量為推薦劑量的1倍以上。王金信[1]也報道了山東地區的馬唐對莠去津產生了抗性。本研究結果證實了廣西甘蔗田馬唐對莠去津已經產生不同程度的抗藥性,相對抗藥性倍數最高達37.2。造成廣西不同地點甘蔗田馬唐抗性程度差異較大的原因,也與當地的用藥歷史、地理特征和作物種植方式有關。造成雜草抗性水平高低差異的原因還與其抗藥機制不同有關。
一些學者已經證明雜草對莠去津的抗性主要機制是GSTs酶活性的增強和psbA靶標基因發生突變。高等植物體內谷胱甘肽-S-轉移酶活性和解毒能力增強是許多雜草對三氮苯類、脲類及酰胺類除草劑的一種重要解毒機制[12-13]。谷胱甘肽-S-轉移酶是一個多功能酶家族,它催化谷胱甘肽與多種親電子和疏水底物相結合,使與之相結合的除草劑或者外源物排除體外從而達到解毒目的。玉米耐受三氮苯類除草劑是一個典型的例子,它因GSTs有較高的催化活性使三氮苯與谷胱甘肽軛合,因此玉米田能廣泛使用三氮苯除草劑。GSTs在玉米體內的解毒機理已經被很好的闡述,該類酶均為二聚體,通過不同的亞基組成不同的同工酶,通過酶解除莠去津毒害。事實也證明,抗性和敏感性苘麻生物型體內類囊體均被莠去津相同程度地抑制,證明其抗藥性不是由于D-1蛋白敏感性降低,三氮苯類除草劑莠去津與谷胱甘肽軛合作用的增強,提高了苘麻(Abutilon theophrasti)對除草劑的解毒能力,才是產生抗藥性的主要原因。
靶標位點突變是導致雜草對PSII抑制劑抗性的另一個重要機制。目前已經報道的突變位點主要包括Val219Ile[14-16],Phe255Ile[17],Ser264Thr[18],Asn266Thr[19],Ala251Val[20],Leu218Val和Ser264Gly[21]。不同的位點突變會導致不同的交互抗性。Val219Ile突變引起雜草對敵草隆、嗪草酮、利谷隆和丁噻隆等4種PSII抑制劑均能產生抗性。然而Ser264Gly突變僅能對三嗪類產生較高的抗性。為了預防雜草抗藥性突變以及控制抗藥性雜草的擴散、蔓延,應避免單一或同一作用機制除草劑;同時,還應及時診斷、鑒別,提供適應于當地抗藥性雜草的防除技術措施,延緩雜草抗藥性的發生,從而延長除草劑的使用壽命,保障雜草的有效治理和農業的可持續發展。
4 結 論
本研究結果表明,廣西大部分甘蔗種植區馬唐對莠去津產生抗性,抗性水平在低到中抗水平。選取不同抗性水平的3個種群對抗性機理進行初探,并沒有發現靶標基因突變,GSTs酶活性的增強可能是抗性主要原因之一。
[1]陳月芳,張超蘭,黃 河,等.有機無機營養物料對莠去津污染土壤微生物量磷和可溶性無機磷的影響[J].安徽農業科學,2015 (11):81-84.
[2]Ryan G F.Resistance of common groundsel to simazine and atrazine [J].Weed Science,1970,18:614-616.
[3]Hirschberg J,McIntosh L.Molecular basis of herbicide resistance in Amaranthus kybridus[J].Science,1983,222(4630):1346-1349.
[4]Anderson M P,Gronwald JW.Atrazine resistance in a velvetleaf(A-butilon theophrasti)biotype due to enhanced glutathione-Stransferase activity[J].Plant Physiology,1991,96:104-109.
[5]Burnet M W M,Loveys B R,Holtum JA M,et al.Increased detoxication is amechanism of simazine resistance in Lolium rigidum[J].Pesticide Biochemistry and Physiology,1993,46:207-218.
[6]Powle S B,Yu Q.Evolution in action:plants resistant to herbicides [J].Annual Review of Plant Biology,2010,61:317-47.
[7]Devine M D,Eberlein C V.Physiological,biochemical and molecular aspects of herbicide resistance based on altered target sites[A].In: Roe RM,Burton JD,Kuhr R J,eds.Herbicide Activity:Toxicology,Biochemistry and Molecular Biology[C].Amsterdam,The Netherlands:IOS,1997:159-185.
[8]Gronwald JW.Resistance to photosystem II inhibiting herbicides [A].In:Powles SB and Holtum JAM,eds.Herbicide Resistance in Plants:Biology and Biochemistry[C].Boca Raton,F L:Lewis,1994:27-60.
[9]Oettmeier W.Herbicide resistance and supersensitivity in photosystem II[J].Cellular and Molecular Life Sciences,1999,55:1255-1277.
[10]王慶亞,董立堯,婁遠來,等.農田雜草抗藥性及其檢測鑒定方法[J].雜草科學,2002(2):1-5.
[11]周 青,高金芬,劉金榮,等.玉米田雜草馬唐對阿特拉津、乙阿合劑的抗藥性試驗簡報[J].雜糧作物,2005,25:274.
[12]Cummins I,Dixon D P,Freitag-Pohl S,etal.Multiple roles for plant glutathione transferases in xenobiotic detoxification[J].Drug Metabolism Reviews,2011,43:266-280.
[13]Dixon D P,Skipsey M,Edwards R.Roles for glutathione transferases in plant secondarymetabolism[J].Phytochemistry,2010,71:338 -350
[14]Mengistu LW,Mueller-WarrantGW,Liston A,etal.psbA mutation (valine219 to isoleucine)in Poa annua resistant to metribuzin and diuron[J].Pest Management Science,2000,56:209-217.
[15]Mengistu L W,Christoffers M J,Lym R G.A psbA mutation in Kochia scoparia(L.)Schrad from railroad rights-of-way with resistance to diuron,tebuthiuron and metribuzin[J].Pest Management Science,2005,61:1035-1042.
[16]Dumont M,Letarte J,Tardif F J.Identification of a psbA mutation (valine219 to isoleucine)in Amaranthus powellii(S.Wats)conferring resistance to linuron[J].Weed Science,2016,64:6-11.
[17]Perez-Jones A,Intanon S and Mallory-Smith C.Molecular analysis of hexazinone-resistant shepherd's-purse(Capsella bursa-pastoris) reveals a novel psbA mutation[J].Weed Science,2009,57:574-578.
[18]Masabni JG,Zandstra BH.A serine-to-threoninemutation in linuron-resistant Portulaca oleracea[J].Weed Science,1999,47:393-400.
[19]Park KW,Mallory-Smith C A.psbA mutation(Asn266 to Thr)in Senecio vulgaris L.confers resistance to several PSII-inhibiting herbicides[J].Pest Management Science,2006,62:880-885.
[20]Mechant E,De Marez T,Hermann O,et al.Target site resistance to metarnitron in Chenopodium album L.[J].Journal of Plant Diseases and Protection XXI(Special Issue),2008:37-40.
[21]Thiel H,Varrelmann M.Identification of a new PSII target-site psbA mutation leading to D1 amino acid Leu218Val exchange in the Chenopodium album D1 protein and comparison to cross-resistance profiles of knownmodifications at positions251 and 264[J].Pest Management Science,2014,70:278-285.
(責任編輯 汪羽寧)
Resistance Level and M echanism s of Digitaria Sanguinalis to Atrazine in Sugarcane Field of Guangxi
WANG Yan-hui1,2,YANG Cai-ying1,MA Yong-lin1,2,HUANG Hui-ye1,2,GUO Cheng-lin1,2,QIN Jian-lin1,2,MA Yue-feng1,2
(1.Plant Protection Research Institute,Guangxi Academy of Agricultural Sciences,Guangxi Nanning 530007,China;2.Guangxi Key Laboratory of Biology for Crop Diseases and Insect Pests,Guangxi Nanning 530007,China)
【Objective】The resistance and resistancemechanism ofweed crabgrass Digitaria sanguinalis to atrazine in Guangxisugarcane field were studied to provide references for scientific control and resistancemanagement.【Method】The resistance level of25 Digitaria sanguinalis population collected in Guangxi sugarcane area was determined by pot experiment and the whole plantmethod.The resistancemechanisms were studied by using physiologicalbiochemical andmolecular biologymethods.【Result】As to the resistance of76%to atrazine in the population,Guigang-1 was the highest,which resistance index(RI)was37.2.Chongzuo-4 wasa sensitive strain,the relative vigor of the GSTs of Baise-1 and Liuzhou 2 population were significantly higher than thatof the sensitive population after3 days,and the psbA gene of D-1 protein on chlorophyll was sequenced,and the psbA gene of the resistant population were found to have nomutation,and the activity of GSTs was increased with the resistance of Digitaria sanguinalis to Atrazine.【Conclusion】The resistance of Digitaria sanguinalis to atrazine in Guangxi sugarcane field is low tomedium level.The resistance of the resistant population to atrazinemay be related to the enhancement of GSTs activity.
Digitaria sanguinalis;Atrazine;Resistance;GSTs;PsbA
S566.1
A
1001-4829(2017)8-1790-05
10.16213/j.cnki.scjas.2017.8.016
2017-03-29
國家自然科學基金項目(31460479);公益性行業(農業)科研專項(201303031);廣西作物病蟲害生物學重點實驗室基金項目(14-045-50-ST-08);廣西農業科學院基本科研業務專項(2015YT41)
王彥輝(1984-),男,河南開封人,博士,副研究員,主要從事雜草抗藥性研究,E-mail:wangyanhui@gxaas.net。
猜你喜歡
科教新報(2022年22期)2022-07-02 12:34:28
今日農業(2021年5期)2021-11-27 17:22:19
高中生學習·閱讀與寫作(2019年2期)2019-09-10 07:22:44
今日農業(2019年15期)2019-01-03 12:11:33
現代園藝(2017年19期)2018-01-19 02:50:21
現代農業(2016年5期)2016-02-28 18:42:36
營銷界(2015年23期)2015-02-28 22:06:18
現代農業(2015年5期)2015-02-28 18:40:49
新疆農墾科技(2014年3期)2014-02-28 19:19:34
中國質量與標準導報(2013年8期)2013-03-11 19:54:07
