云南撒壩豬EPEC HPI irp2缺失株的攝鐵能力研究
2017-09-15 07:32:05劉超英吳程華嚴玉霖富國文苗淑淑
西南農業學報 2017年8期
劉超英,吳程華,高 洪,嚴玉霖,富國文,趙 汝,苗淑淑,劉 嫦
云南撒壩豬EPEC HPI irp2缺失株的攝鐵能力研究
劉超英,吳程華,高 洪*,嚴玉霖,富國文,趙 汝,苗淑淑,劉 嫦
(云南農業大學動物科學技術學院,云南昆明 650201)
【目的】本研究探討了云南撒壩豬EPEC HPI毒力島的攝鐵功能與致病性之間的關系。【方法】本試驗通過攝鐵實驗對5株腸致病性E.coli(A-E)和CVCC1565鐵載體的合成和在不同2,2’-聯吡啶含量下的最低生長抑制濃度及對數期生長速率進行了測定,同時利用已構建的E.coli irp2基因缺失株和親本株進行對比,檢測HMWP2蛋白的表達,并通過半數致死量實驗比較兩者之間的毒力。【結果】除E.coli(D)外,其余菌株的2,2’-聯吡啶最低生長抑制濃度為0.6 mM;親本株能夠表達HMWP2蛋白,irp2缺失株不表達該蛋白;CAS固體培養基中菌株能產生橘黃色的透明暈圈;當2,2’-聯吡啶濃度高于0.2 mM時,親本株的生長率高于irp2缺失株;親本株的LD50(8.73×107cfu/mL)低于缺失株(9.95×107cfu/mL)。【結論】云南撒壩豬EPEC HPI鐵攝取功能和致病性之間存在著一定關系。
EPEC;HPI;irp2基因;攝鐵功能
【研究意義】撒壩豬是云南省優良的地方豬種,具有繁殖能力和抗病力強,耐粗飼,肉質好等優點[1],但因飼養水平較低,使腸致病性大腸桿菌(Enteropathogenic Escherichia coli,EPEC)感染引起的原發性或繼發性豬大腸桿菌病成為嚴重威脅云南特色養豬業發展的重要因素。【前人研究進展】有研究表明,鐵元素是微生物生長的必須元素,鐵載體能夠特異性結合鐵離子并運輸至細胞內[2-3]。毒力島(High-pathogeniticity island,HPI)是研究細菌的毒力因子和致病性之間關系的基因簇,與細菌的毒力密切相關。耶爾森菌強毒力島主要含有與攝鐵有關的毒力基因簇[4],研究表明耶爾森桿菌HPI可以在耶爾森菌和E.coli之間水平傳播,與EPEC的毒力進化有著密切的關系[5]。其中irp1、irp2基因在鐵缺乏的條件下分別表達2個高分子量蛋白:HMWP1和HMWP2[6]。HMWPs(High molecular weight pro-teins,高分子量鐵調節蛋白)參與表達耶爾森桿菌素的陽性表型,被認為對鐵載體耶爾桿菌素的合成有著非常重要的作用。【本研究的切入點】HMWP2的表達及CAS平板上暈圈的大小能夠證明細菌攝鐵能力的高低,親本株和缺失株LD50的大小與細菌毒力直接相關。【擬解決的關鍵問題】因此通過試驗研究irp2基因與細菌攝鐵能力之間的關聯,探索其對細菌致病性的影響,不僅可對豬源EPEC HPI與攝鐵能力之間的關系進一步深入研究,還可闡明HPI的致病過程和致病機理,具有非常重要的現實意義。
1 材料與方法
1.1 試驗材料
5株E.coli分離株(A-E)、CVCC1565菌株和利用RED同源重組方法[7]構建的E.coli irp2基因缺失株均由云南農業大學獸醫研究實驗室保存提供。試驗小鼠購自昆明醫科大學。
1.2 試驗方法
1.2.1 2,2’-聯吡啶對實驗菌株的最低抑制濃度的測定 分別配置含有0.1、0.2、0.3、0.4、0.5、0.6、0.7、0.8、0.9、1.0 mM 2,2'-聯吡啶的LB濃度梯度培養基,細菌不能生長的某一個濃度即為2,2'-聯吡啶的最低抑制濃度。
1.2.2 實驗菌株鐵載體的檢測 采用通用鉻天青S(Chrom azurol S,CAS)檢測法對E.coli內存在的鐵載體進行初步檢測[8]。將E.coli(A-E)和CVCC1565菌株的少許單菌落點種在CAS平板上,37℃培養,每12 h觀察1次菌落周圍顏色的變化,測定菌落直徑直至不再變化。
1.2.3 irp2基因缺失株與親本株蛋白表達差異的比較 通過SDS-PAGE電泳分析比較缺失株E.coli (E)/Δirp2與親本株E.coli(E)蛋白表達圖譜的差異。
1.2.4 親本株E.coli(E)和缺失株E.coli(E)/ Δirp2生長速率的測定 分別于第0、1.5、3、4、6、8、10、12、14、16、18、20小時測定含有0.1、0.2、0.3、0. 4、0.5、0.6、0.7、0.8、0.9、1.0 mM 2,2'-聯吡啶的LB濃度梯度培養基中親本株和irp2缺失株菌液的OD600值,利用公式計算(式中t1和t2為所取對數期2點的時間,W1和W2分別為相應時間測的的OD600值,G為代時),并繪制生長率曲線。1.2.5 親本株E.coli(E)和缺失株E.coli(E)/ Δirp2 LD50的測定 用生理鹽水將E.coli(E)和E. coli(E)/Δirp2肉湯培養液分別稀釋至3×108、3× 107、3×106、3×105、3×104cfu/mL。將75只實驗小鼠平均分為1、2和3號組。1和2號組以0.1 mL/只劑量分別腹腔注射親本株和缺失株培養液,3號組為對照組。注射后每4 h觀察1次,連續觀察3 d。記錄其死亡數量,并按Reed-Muench法計算半數致死量。
2 結果與分析
2.1 2,2'-聯吡啶對6株E.coli的最低抑制濃度的測定
在不同濃度的2,2′-聯吡啶限鐵環境,E.coli(D)在0.4 mM濃度下停止生長,其余5株均在0.6 mM濃度下停止生長。
2.2 HPI陽性株鐵載體的檢測
除了E.coli(D),檢測的6株E.coli在CAS固體平板上生長良好,培養1 d后就有明顯的透明暈圈出現在菌落的周圍,8 d后暈圈直徑達到最大。可以斷定產生橘黃色透明暈圈的E.coli在生長過程中分泌的物質對鐵有較高的親和力,使培養基發生明顯的由藍到橙的顏色改變。表1為實驗菌株產生的暈圈大小情況。
2.3 親本株與irp2缺失株蛋白表達差異的比較
在2,2'-聯吡啶濃度為0.2 mM的情況下,親本株能夠表達HMWP2,但是irp2缺失株不能表達HMWP2(圖1)。
2.4 親本株與irp2缺失株在不同2,2'-吡啶濃度下生長速率的測定
以不同的鐵螯合劑濃度為橫坐標,生長率為縱
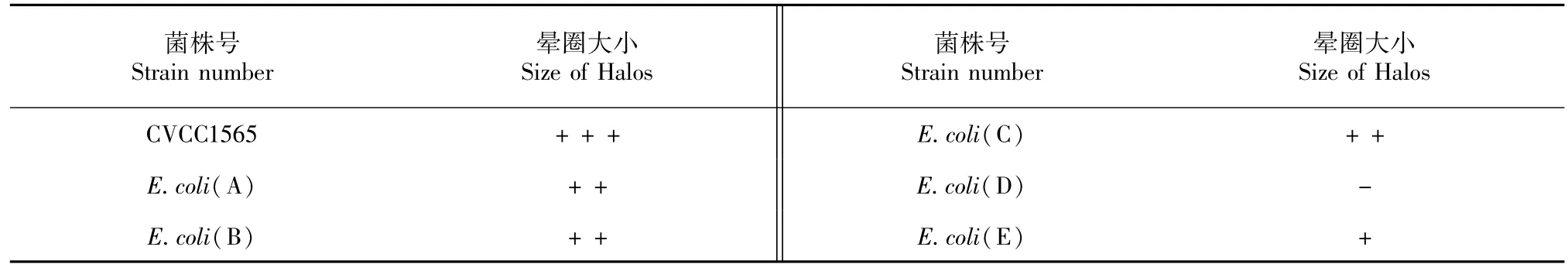
表1 鐵載體產生菌產生的暈圈大小情況Table 1 The size of halos of the siderophore producing bacteria
坐標,繪制出親本株和irp2基因缺失株的生長速率曲線(圖2)。親本株和缺失株均隨著鐵螯合劑濃度的升高而降低,親本株在0.6 mM濃度時停止生長;而缺失株在0.4mM時即停止生長,irp2基因缺失株的攝鐵能力顯著降低。
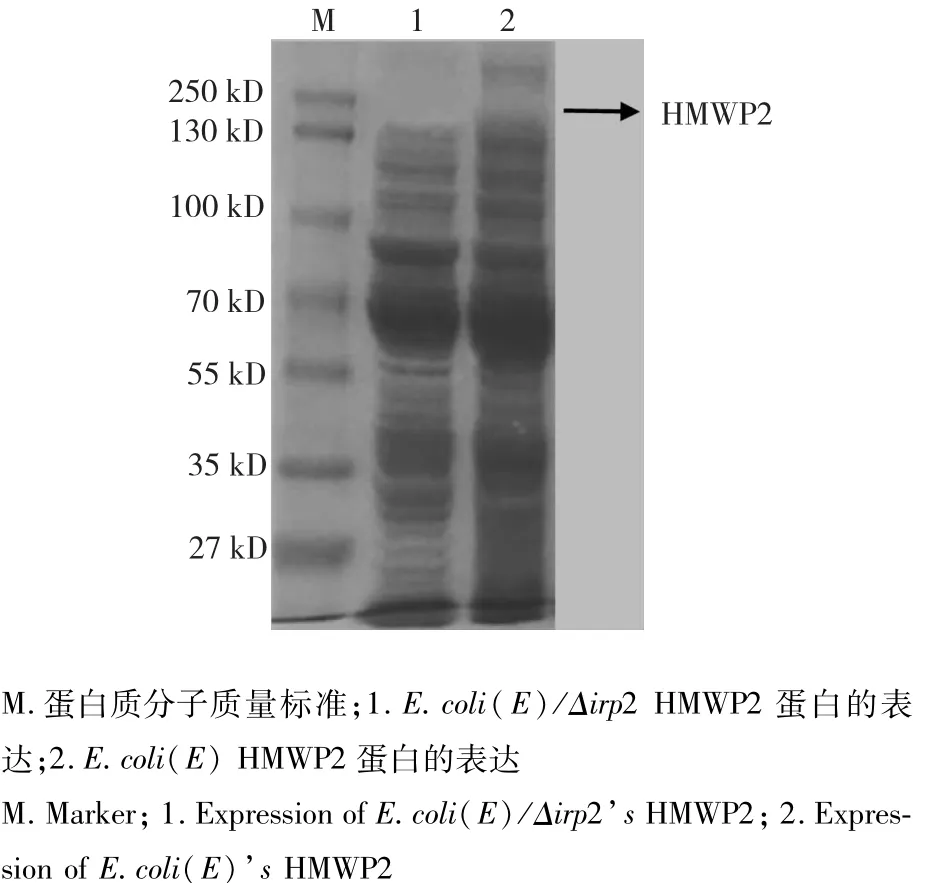
圖1 irp2親本株E.coli(E)與缺失株E.coli(E)/Δirp2蛋白表達比較Fig.1 Comparsion of proteins of E.coli(E)and E.coli(E)/Δirp2 expressed under iron starvation
2.5 親本株與irp2缺失株LD50的測定
感染3 d后對照組小鼠均健康存活;攻毒后6 h小白鼠開始陸續死亡,死亡高峰發生在接種后7~13 h。缺失株的LD50為9.95×107cfu/mL,親本株為8.73 ×107cfu/mL,因此可判斷親本株的毒力大于缺失株。
3 討 論
隨著養豬業的集約化發展,EPEC所導致的仔豬黃痢、仔豬白痢和水腫病成為養豬場常發的傳染病,對養豬業造成了較大的威脅和重大的經濟損失[9-10]。E.coli的致病性主要取決于鐵攝取系統、菌毛及毒素等特定的毒力因子,HPI毒力島中的irp1、irp2及fyuA是主要鐵調節基因[11-14],能夠通過Ybt的鐵載體奪取宿主中的鐵元素進而加重機體的感染,從而增強細菌的存活率,它們在攝鐵菌素的生物合成、調節及攝取中發揮著極其重要的作用[15-18]。HPI毒力島在豬源EPEC中有著很高的攜帶率[19],對毒力島的攝鐵功能及其與宿主致病機制之間的關系進行初步探索,對防治豬源致病性大腸桿菌有著重要的理論意義和實踐價值。

圖2 親本株與irp2缺失株在不同濃度2,2'-聯吡啶下的生長率Fig.2 The growth curves of parent strain and irp2 deletion strain
鐵元素是微生物生長的必須元素。當細菌處于缺鐵環境時,會產生鐵載體及其特異性受體[20]。2,2'-聯吡啶是一種鐵螯合劑,可以結合培養基中的鐵離子,使細菌生長環境處于鐵缺乏狀態,從而測定細菌的最低抑制濃度。試驗結果顯示,除E.coli(D)在0.4 mM濃度下停止生長外,其余5株均在0.6 mM濃度下停止生長。鐵載體是一類能特異性結合鐵離子的低分子量的物質,CAS結合鐵后形成一種藍色復合物,當鐵載體與復合物中的鐵結合后,其顏色可由藍色變為橘黃色。在CAS培養基中,可以出現鐵載體的分泌圈,從而判斷細菌是否產生了鐵載體。本試驗中,CVCC1565的暈圈最大,E.coli(E)暈圈最小,E.coli(D)無暈圈。
本試驗采用SDS-PAGE技術對irp2缺失株和親本株的HMWP2蛋白的表達進行了檢測,結果顯示,親本株可以表達該蛋白,而缺失株則不能表達。有研究表明,irp2基因在鐵缺乏的條件下可表達HMWP2蛋白。由生長曲線可知,當2,2'-聯吡啶濃度大于0.2 mM時,缺失株E.coli(E)/Δirp2對數期的生長速率低于親本株E.coli(E),當2,2'-聯吡啶濃度為0.4 mM時,缺失株E.coli(E)/Δirp2不再生長,而親本株E.coli(E)在0.6 mM時停止生長。
4 結 論
irp2基因不是E.coli生長繁殖的必須功能基因,該基因的缺失對其生長會產生影響,但不會導致死亡。這可能是菌株在缺失irp2基因后不能很好的吸收營養物質,相對于親本株生長較慢。半數致死量試驗結果表明,缺失株E.coli(E)/Δirp2的LD50高于親本株,由此推斷能表達HMWP2的菌株毒力強于不表達HMWP2蛋白的菌株,而E.coli(E)/Δirp2雖然缺失了HPI irp2鐵調節基因,由于存在著另外的鐵攝取系統,仍然能夠在缺鐵的環境下生長,且其攝鐵能力和致病性之間存在著一定的聯系。
[1]呂春榮,權國波,吳國權,等.撒壩豬成纖維細胞系的建立及其生物學特性分析[J].黑龍江動物繁殖,2016,24(2):9-13.
[2]Miethke M,Marahiel M A.Siderophore-based iron acquisition and pathogen control[J].Microbiology&Molecular Biology Reviews,2007,71(3):413-451.
[3]Beasley FC,Heinrichs D E.Siderophore-mediated iron acquisition in the staphylococci[J].J Inorq.Biochem.,2010,104(3):282-288.
[4]Uladzimir A,Christina N,Jürgen H,et al.Independent acquisition of site-specific recombination factorsby asn tRNA gene-targeting genomic islands[J].International Journal of Medical Microbiology,2006,296(6):341-352.
[5]Rakin A,Schubert,Guilvout I,et al.Local hopping of IS3 elements into the A+T-rich part of the high-pathogenicity island in Yersinia enterocolitica 1B,O:8[J].Fems Microbiology Letters,2000,182 (2):225-229.
[6]李葉芳,涂 健,邵 穎,等.利用Red重組系統敲除APEC毒力島irp2基因[J].安徽農業大學學報,2012,39(6):854-858.
[7]吳程華,嚴玉霖,高洪,等.RED同源重組技術在大腸埃希菌基因敲除中的應用[J].上海畜牧獸醫通訊,2015(1):9-11.
[8]李葉芳,白 灝,彭開松,等.禽致病性大腸桿菌強毒力島攝鐵功能與致病性關系的研究[J].中國獸醫科學,2012,42(9):906-910.
[9]陳 重,鄭金鑫,楊唯枝,等.腸致病性大腸埃希菌EPEC Deng致病島基因進化特點[J].中國感染控制雜志,2016(1):1-9.
[10]張 強.腸致病性大腸桿菌EPEC及其突變株UMD874感染對腸粘膜屏障功能及腸道菌群的影響[D].南京大學,2011.
[11]邵 穎,涂 健,汪雪雁,等.APEC強毒力島核心基因irp2、fyuA敲除對其致病性影響的研究[J].中國獸醫學報,2014(4): 564-570.
[12]盧 琴,高 洪,嚴玉霖,等.撒壩豬大腸桿菌血清型鑒定及藥敏試驗與HPI irp2基因的檢測[J].黑龍江畜牧獸醫,2014(3): 115-117.
[13]冀 輝,邵長勝,涂 健,等.禽源大腸桿菌耶爾森氏菌強毒力島核心基因的檢測及其irp2和int基因同源性[J].微生物學通報,2013(11):2022-2029.
[14]董炳敏.大腸桿菌HPI毒力島irp1與irp2基因的研究[D].揚州大學,2010.
[15]盧 琴,高 洪,嚴玉霖,等.撒壩豬大腸桿菌血清型鑒定及藥敏試驗與HPI irp2基因的檢測[J].黑龍江畜牧獸醫,2014(3): 115-117.
[16]李葉芳,白 灝,彭開松,等.禽致病性大腸桿菌強毒力島攝鐵功能與致病性關系的研究[J].中國獸醫科學,2012,42(9):906 -910.
[17]Fischbach M A,Lin H N,Liu D R,et al.How pathogenic bacteria evademammalian sabotage in the battle for iron[J].Nat Chem.Biol.,2006,2(3):132-138.
[18]Perry R D,Fetherston JD.Yersiniabactin iron uptake:mechanisms and role in Yersinia pestis pathogenesis[J].Microbes Infect.,2011,13(10):808-817.
[19]王 勇,王 紅,向 前,等.產毒性大腸桿菌(ETEC)中毒力島分布的研究[J].熱帶醫學雜志,2001(1):17-21.
[20]孫紅啟.鐵載體和鐵離子對細菌生長過程的影響[D].山東大學,2008.
(責任編輯 王家銀)
Iron-uptake Function of EPEC HPI irp2 Gene Deletion Strains Isolated from Yunnan Saba Pigs
LIU Chao-ying,WU Cheng-hua,GAO Hong*,YAN Yu-lin,FU Guo-wen,ZHAO Ru,MIAO Shu-shu,LIU Chang
(Faculty of Animal Science and Technology,Yunnan Agricultural University,Yunnan Kunming 650201,China)
【Objective】The present paper aims to explore the relationships between iron-uptake function of EPEC HPI and pathogenicity in Yunnan Saba pigs.【Method】This experiment tested the synthesis of siderophore,theminimum growth inhibition concentration under different2,2′-Dipyridyl contentand growth rate of5 strains E.coli and CVCC1565.The virulence of strain and the expression of HMWP2 were also detected respectively bymedian lethal dose experiment and SDS-PAGE.【Result】Except E.coli(D),theminimum growth inhibition concentration of other strainswere0.6 mM,the parent strain could expressed HMWP2 protein,but themutation strain could notand strains cultured by CAS solid culture medium produced orange transparent halo.When the content of 2,2′-Dipyridyl was higher than 0.2 mM,the growth rate of parent strain was also higher than mutation strain,parent strain and mutation strain's LD50was 8.73×107and 9.952× 107cfu/m L,respectively.【Conclusion】Above results showed that there was an certain relationship between iron-uptake function of EPEC HPIand pathogenicity in Yunnan Saba pigs.
EPEC;HPI;irp2 gene;Iron-uptake function
S828
A
1001-4829(2017)8-1914-04
10.16213/j.cnki.scjas.2017.8.038
2016-01-15
國家自然科學基金(31260594、31660704);云南生豬產業技術體系建設專項經費資助(云財農[2009]171號);云南農業大學科技創新創業行動基金(2017ZKX030)
劉超英(1992-),女,河南安陽人,在讀博士,從事特種經濟動物健康養殖方向研究,E-mail:1107946950@qq.com,*為通訊作者:E-mail:gaohongping@163.com。
猜你喜歡
中國糖料(2023年4期)2023-11-01 09:34:46
環境保護與循環經濟(2021年7期)2021-11-02 08:10:42
林業科技情報(2020年3期)2020-03-02 13:21:36
生物工程學報(2019年11期)2019-12-09 13:09:12
西南農業學報(2019年6期)2019-07-18 05:32:16
中國果業信息(2019年11期)2019-01-05 20:47:24
西南農業學報(2016年5期)2016-05-17 05:42:23
鄭州大學學報(理學版)(2014年3期)2014-03-01 04:21:06
新疆農墾科技(2014年10期)2014-02-28 19:21:08
無機化學學報(2014年12期)2014-02-28 17:33:51
