甜櫻桃斑果病研究進展及其防治
2017-09-16 04:45:16劉慶忠
落葉果樹 2017年5期
關鍵詞:檢測
劉慶忠
(山東省果樹研究所,山東泰安 271000)
甜櫻桃斑果病研究進展及其防治
劉慶忠
(山東省果樹研究所,山東泰安 271000)
類病毒是一類單鏈、閉合環(huán)狀低分子量的致病性RNA,大小為246~463bp,無蛋白質外殼包裹,是目前發(fā)現(xiàn)的最小病原物。類病毒侵染性強,可導致被侵染的細胞或組織發(fā)生異常,侵染后造成類似病毒感染的矮化、斑駁、葉變形、裂皮、斑果和壞死等癥狀。從類病毒的危害癥狀、檢測及鑒定、致病機理、傳播及防治等方面介紹了引起甜櫻桃斑果病的啤酒花矮化類病毒。
啤酒花矮化類病毒;甜櫻桃;斑果病;鑒定;癥狀;防治

隨著甜櫻桃新品種、新砧木和高效栽培技術在生產上的廣泛應用,甜櫻桃種植在中國取得了快速發(fā)展。2016年中國甜櫻桃栽培面積達18萬hm2,產量70萬t,種植面積在近幾年翻了兩翻。在生產快速發(fā)展的同時,新的病害不斷出現(xiàn),如葉片退綠黃化、花而不實、果實畸形、皺縮、果實出現(xiàn)斑點、果實變小、叢枝、枯枝死樹等,嚴重影響了甜櫻桃的產量和品質。
1971年美國科學家Diener在研究馬鈴薯紡錘塊莖病的過程中,提出了“類病毒”的概念[1]。類病毒(Viroid)是一類單鏈、閉合環(huán)狀低分子量的致病性RNA,大小為246~463bp,無蛋白質外殼包裹,是目前發(fā)現(xiàn)的最小病原物。類病毒侵染性強,通常可導致被侵染的細胞或組織發(fā)生異常。侵染后造成類似病毒感染的矮化、斑駁、葉變形、裂皮、斑果和壞死等癥狀。筆者主要介紹由啤酒花矮化類病毒(Hop stunt viroid,HSVd)引起的甜櫻桃斑果病(Dapple fruit)。
1 HSVd在櫻桃上的危害癥狀
2008年土耳其研究者首次報道了啤酒花矮化類病毒侵染甜櫻桃和酸櫻桃,研究者在26個果園收集了127株樹上的樣品,采集樣品均無癥狀表現(xiàn),經RT-PCR檢測甜櫻桃HSVd感病率為13.5%,酸櫻桃感病率為45.5%[2]。中國于2017年首次報道了啤酒花類病毒能引起“紅燈”甜櫻桃果實出現(xiàn)深紅色斑點,即“Dapple fruit”的癥狀[3]。進一步田間調查發(fā)現(xiàn),該病害在山東、遼寧、山西、甘肅、北京、陜西等甜櫻桃產區(qū)普遍存在。甜櫻桃品種紅燈、早大果、紅蜜、雷尼、美早、薩米特、桑提娜、先鋒、芝罘紅和布魯克斯等均有發(fā)生。發(fā)病嚴重的樹體不僅出現(xiàn)斑果,而且果實變小、皺縮,成熟期變晚,失去商品價值。有的還表現(xiàn)出枯枝、樹勢衰退甚至整枝死亡等癥狀。嫁接樹結果后2~3年,即表現(xiàn)典型的斑果癥狀(圖1)。目前,中國櫻桃還未見染病現(xiàn)象。
2 甜櫻桃HSVd的檢測及其鑒定
類病毒分為馬鈴薯紡錘塊莖類病毒科(Pospiviroidae)和鱷梨日斑類病毒科(Avsunviroidae)2個科。至今發(fā)現(xiàn)的30多種類病毒,其中大部分屬于馬鈴薯紡錘塊莖類病毒科,其科下分為5個屬28個種,引起甜櫻桃斑果病的HSVd是該科啤酒花矮化類病毒屬(Hostuviroid)的唯一成員,無蛋白質外殼包裹,又無編碼蛋白的功能,不能采用酶聯(lián)免疫吸附反應(ELISA)等血清學方法進行檢測。目前,常用的檢測方法主要有生物學檢測及分子生物學技術檢測兩種。

圖1 甜櫻桃斑果病田間癥狀
2.1 指示植物法
HSVd檢測采用四葉黃瓜(CucumissativusL. cv Suyo)作為指示植物。具體的做法是:將營養(yǎng)土裝入塑料營養(yǎng)缽內,用水均勻灌溉,無積水時將黃瓜種整齊的播種于營養(yǎng)缽內,置于溫室條件下生長。待黃瓜長至二葉期時,將待檢測的各樣品低分子RNA溶液與接種緩沖液(100mM Tris-base,10mM EDTA,pH7.5)以1∶100的比例混合后,用棉棒蘸取摩擦接種于黃瓜葉片上,以不接種的四葉黃瓜作為陰性對照,每個樣品接種10株。待黃瓜長出第四片真葉時,將每株分別移栽于營養(yǎng)缽內,溫室保持在28~32℃。觀察記錄黃瓜生長情況。接種3~4周后接種植株發(fā)生植株明顯矮化,節(jié)間縮短,葉片翻卷等現(xiàn)象(圖2),健康植株未出現(xiàn)任何癥狀。生物學鑒定能夠提供類病毒繁殖和致病的證據(jù),是一種重要的檢測方法。
2.2 RT-PCR擴增
逆轉錄-聚合酶鏈式反應(RT-PCR)是目前非常成熟、應用廣泛,靈敏度較高的檢測方法,已成功用于檢測多種類病毒。HSVd檢測具體操作為:選取健康植株和具有斑果癥狀的甜櫻桃葉片作為試材,提取總RNA,利用引物R1 (5′-GCTGGATTCTGAGAAGAGTT-3′)進行反轉錄合成cDNA,然后用R2 (5′-AAACCCGGGGCTCCTTTCTCA-3′) 和 F1 (5′-AAACCCGGGGCAACTCTTCTC-3′) 進行 PCR擴增[4]。對獲得的產物進行連接轉化、測序。測序結果與Genebank數(shù)據(jù)庫中已有序列比對,構建系統(tǒng)進化樹。結果表明導致甜櫻桃(Prunusavium)斑果病的HSVd與李屬(Prunussalicina)HSVd親緣關系最近(圖3)。

圖2 接種四葉黃瓜發(fā)病植株與健康植株
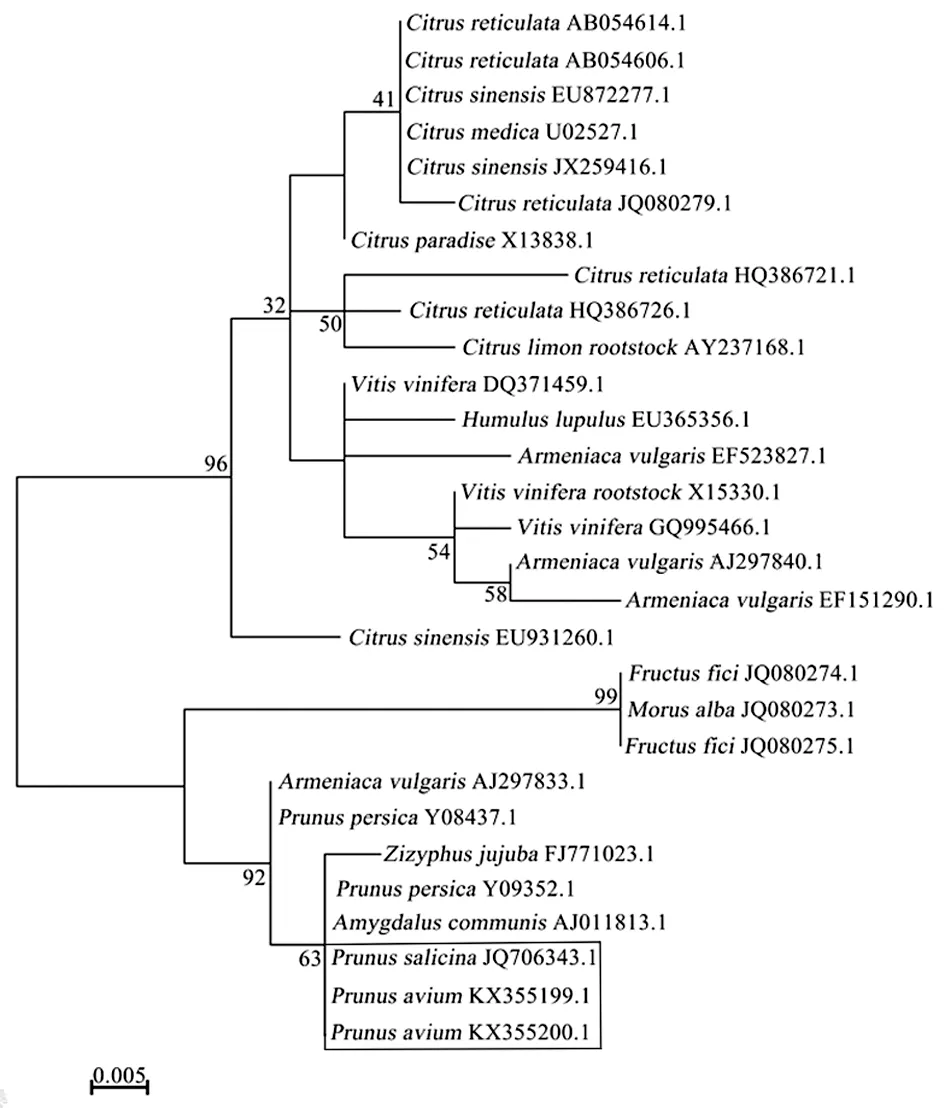
圖3 甜櫻桃HSVd序列與下載HSVd序列系統(tǒng)進化關系
2.3 高通量深測序技術(NGS)
高通量深測序技術具有非序列依賴性,能同時檢測植物樣品中可培養(yǎng)和不可培養(yǎng)的、含量高及含量低的所有DNA病毒、RNA病毒以及類病毒。通過高通量測序可對一個物種的轉錄組和基因組進行全面的分析,然后利用生物信息學軟件進行基因組組裝,完成病毒或類病毒的全基因組測序。相對于其他的植物病毒類病毒檢測方法,NGS具有其優(yōu)勢,主要表現(xiàn)在5個方面:①NGS能快速檢測病毒、類病毒,傳統(tǒng)方法檢測周期長,而NGS一般一個測序反應需要2~3天即可完成,后期的數(shù)據(jù)分析和病毒或類病毒的確認一般也可在短時間內完成,因此,NGS能夠快速診斷生產實踐上的疑難雜癥和一些爆發(fā)性病害的病原,可為病害防治提供依據(jù)。②NGS可廣譜的檢測病毒或類病毒的寄主,無論是DNA病毒、RNA病毒還是類病毒,都能在一個測序反應中被檢測到,不受病毒基因組類型的限制。③NGS檢測病毒或類病毒靈敏度高。NGS測定的是在RNA沉默這個寄主防衛(wèi)反應過程中抵抗病毒或類病毒侵染產生的高度冗余的小RNA或病毒的轉錄本,所以即使極低效價的病毒或類病毒也能被鑒定出來。④NGS能夠對多個不同來源的樣品同時進行病毒或類病毒檢測。由于每個植物樣品的cDNA都帶有特異的標簽,因此序列來源可以追溯到原始地理帶的不同植物間,能快速比較病毒的分布和變異進化等特征。⑤NGS能夠通過非序列同源性的方式發(fā)現(xiàn)病毒或類病毒。傳統(tǒng)方法對于那些與已知的病毒或類病毒序列完全缺乏同源性的新序列在認定上存在困難,目前已經通過NGS測定siRNA和新的逐漸過濾重疊siRNA的數(shù)據(jù)分析方法,在葡萄等植物上發(fā)現(xiàn)了與已知的類病毒序列完全缺乏同源性的新類病毒。
3 HSVd的致病機理
HSVd是由294~303個核苷酸序列組成的RNA片段,二級結構如圖所示(圖4)。病毒侵入寄主敏感細胞后,通過細胞質到達相應的細胞核,在細胞核內寄主DNA依賴的RNA聚合酶(DNA dependent RNA Polymerase)作用下復制,新合成的類病毒個體又反向細胞質,通過胞間連絲進入相鄰的細胞,然后進入韌皮部維管系統(tǒng),長距離運輸?shù)街仓甑母鱾€部位,侵入植株其他的健康細胞。類病毒本身的RNA序列,RNA二級結構和高級結構與寄主細胞內的RNA分子互作,通過RNA沉默,激素調節(jié),特定基因的表達,調控基因細胞的蛋白質合成,干擾寄主正常的新陳代謝,導致植株表現(xiàn)出明顯的發(fā)育障礙,出現(xiàn)矮化、皺縮、花葉、病斑等癥狀。據(jù)報道,HSVd能夠影響植株體內MYB,bHLH和WRKY轉錄因子的表達[5]。
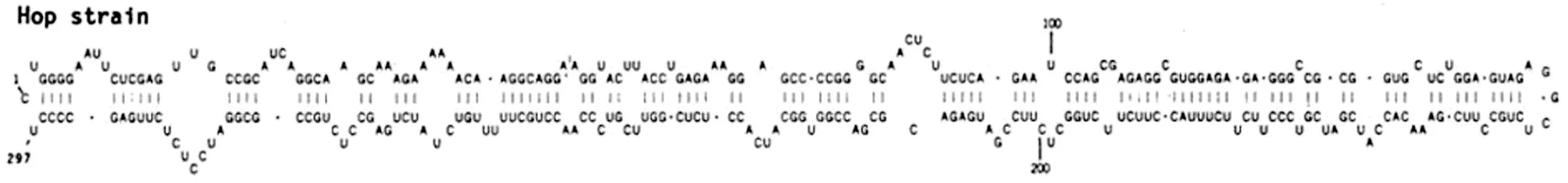
圖4 HSVd類病毒二級結構圖(Ohno et al., 1983)
4 HSVd的傳播與防治
HSVd首次在日本啤酒花上分離得到,自然寄主范圍十分廣泛,包括啤酒花、黃瓜、石榴、柑桔、梨、蘋果、桃、李、扁桃、櫻桃、棗和無花果等草本和木本植物,世界各國均有發(fā)生。類病毒可以通過帶毒材料的無性繁殖、機械摩擦、帶毒的種子或者花粉以及昆蟲介體等方式進行傳播[6]。主要傳播途徑是采用帶毒植物材料的無性繁殖。一旦母體材料被感染,通過快速擴繁,短時間內類病毒就能大規(guī)模擴散。20世紀六、七十年代,啤酒花矮化病害在日本突然大規(guī)模流行,給日本的啤酒花生產和啤酒釀造業(yè)造成了嚴重的沖擊[7]。田間農事操作過程中,類病毒能夠通過被污染的操作工具進行傳播。尤其是對果樹來說,嫁接和修剪也是類病毒傳播的重要途徑之一。類病毒還能夠通過被感染的種子或者花粉進行傳播[8]。此外,類病毒還可以通過昆蟲及相鄰植株間的汁液進行傳播。如野生寄主中的類病毒可經蚜蟲、桃蚜傳播到實驗寄主中。然而大量實驗證明,昆蟲雖可作為類病毒傳播的介體,但其傳播效率較低且持久性比較弱。
HSVd在杏、棗、梨和蘋果等植物中呈潛伏侵染,在某些寄主上可造成嚴重的病害,如啤酒花矮化病、柑桔矮化裂皺病、李和桃斑果病等。HSVd侵染寄主植物后是否有癥狀表現(xiàn)與地區(qū)、品種有關,如HSVd在土耳其侵染甜櫻桃,感染率達13.8%,但未見有癥狀報道。而在中國HSVd侵染甜櫻桃引起嚴重斑果病、縮果病、小果病、枝枯病、植株流膠、整株死亡等現(xiàn)象,個別果園100%植株感染發(fā)病,危害十分嚴重。應當加強綜合防治。首先應制定法律法規(guī),建立嚴格的檢疫制度,防止病害擴散。其次,生產上要使用無毒苗木。第三,建立健全果園巡視制度,及時淘汰果園病株。第四,甜櫻桃生產園、苗圃必須與生產櫻花及其他核果類、棗、葡萄、柑桔類和仁果類等果樹的果園隔離。第五,及時清除果園雜草,防止交叉感染。第六,加強熱處理、組織培養(yǎng)、超低溫冷凍脫毒技術研究、培育無病毒苗,加強弱株系交叉防護、開展轉基因抗類病毒病研究、培育抗病品種。
值得注意的是,類病毒相對于病毒病來說,雖然類病毒的寄主范圍相對較窄,但近來研究發(fā)現(xiàn)類病毒的寄主范圍有逐漸擴大的趨勢[9]。如長期以來認為馬鈴薯紡錘塊莖類病毒(PSTVd)只侵染馬鈴薯和番茄,但有報道表明鱷梨也是其自然寄主;開始HSVd只在啤酒花中被發(fā)現(xiàn),后續(xù)研究表明其還能侵染黃瓜、蘋果、桃、李、棗、無花果和梨等草本和木本植物。此外,有研究表明與馬鈴薯和啤酒花生長在同一農田的野草,雖不是PSTVd和HSVd的自然寄主,接種類病毒cDNA后,卻能夠支持類病毒在其體內進行復制。因此,如果條件合適,類病毒可能會侵染一些新的植物,所以應該嚴格防控類病毒傳播。
[1] Diener, T.O. Potato spindle tuber "virus":IV. A replicating, low molecular weight RNA[J]. Virology, 1971, 45:411-428.
[2] Gazel M, Ulubas C, CAg Layan K, et al. Detection of Hop stunt viroid in sweet and sour cherry trees in Turkey by RT-PCR[J].Acta Horticulturae, 2008, 795(795):955-958.
[3] Xu L.javascript:popRefFull('aff1'), Wangjavascript:popRefFull('aff1') J.W., Zhu D Z.javascript:popRefFull('aff1'), et al. First Report of Hop stunt viroid From Sweet Cherry With Dapple Fruit Symptoms in China[J]. Plant disease, 2017, 101(2):394.
[4] Hassan M., Zouhar M., Rysanek P. Development of a PCR method of each latent mosaic viroid and hop stunt viroid detection for certification of planting material. Acta Horti., 2004, 675: 391-5.
[5] Zoltán Füssya, Josef Patzakc, Jan Stehlíkb, et al. Imbalance in expression of hop (Humulus lupulus) chalcone synthase H1 and itsregulators during hop stunt viroid pathogenesis[J]. Journal of Plant Physiology, 2013, 170:688-695.
[6] 張志想. 啤酒花矮化類病毒寄主適應性和致病性研究 [D]. 北京:中國農業(yè)科學院, 2012.
[7] Yamamoto, H., Kagami, Y., Kurokawa, M., et al. Studies on hop stunt disease in Japan[J]. Pharmaceutical Research Laboratory, Kirin Brewery Co., Ltd 1973, 16: 49.
[8] Mink, G. Pollen- and seed-transmitted viruses and viroids. Ann. Rev. Phytopath., 1993, 31, 375-402.
[9] Ding, B. The biology of viroid-host interactions[J]. Annual Review of Phytopathology 2009, 47: 105-131.
2017-02-13
“十二五”國家科技支撐計劃(2013BAD02B03-3-2); 山東省現(xiàn)代農業(yè)產業(yè)技術體系果品創(chuàng)新團隊(SDAIT-06-04)。
劉慶忠(1963-),男,山東莒南人, 研究員,主要從事果樹種質資源與生物技術育種工作。E-mail: qzliu001@126.com
S662.5
A
1002-2910(2017)05-0001-04
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2020年12期)2021-01-18 06:57:46
中學生數(shù)理化·七年級數(shù)學人教版(2020年12期)2021-01-18 06:57:46
中學生數(shù)理化·七年級數(shù)學人教版(2019年9期)2019-11-25 07:34:36
中學生數(shù)理化·七年級數(shù)學人教版(2019年9期)2019-11-25 07:34:34
中學生數(shù)理化·七年級數(shù)學人教版(2019年12期)2019-05-21 02:53:50
中學生數(shù)理化·七年級數(shù)學人教版(2019年12期)2019-05-21 02:53:48
