元江干熱河谷稀樹(shù)灌草叢植被碳儲(chǔ)量及凈初級(jí)生產(chǎn)力
2017-10-13 12:32:47金艷強(qiáng)張一平劉運(yùn)通費(fèi)學(xué)海李培廣
生態(tài)學(xué)報(bào) 2017年17期
金艷強(qiáng),李 敬,張一平,劉運(yùn)通,費(fèi)學(xué)海,李培廣
1 中國(guó)科學(xué)院西雙版納熱帶植物園,熱帶森林生態(tài)學(xué)重點(diǎn)實(shí)驗(yàn)室, 昆明 650223 2 中國(guó)科學(xué)院西雙版納熱帶植物園,元江干熱河谷生態(tài)站, 元江 653300 3 中國(guó)科學(xué)院大學(xué),北京 100049
元江干熱河谷稀樹(shù)灌草叢植被碳儲(chǔ)量及凈初級(jí)生產(chǎn)力
金艷強(qiáng)1,3,李 敬1,3,張一平1,*,劉運(yùn)通1,費(fèi)學(xué)海1,3,李培廣2
1 中國(guó)科學(xué)院西雙版納熱帶植物園,熱帶森林生態(tài)學(xué)重點(diǎn)實(shí)驗(yàn)室, 昆明 650223 2 中國(guó)科學(xué)院西雙版納熱帶植物園,元江干熱河谷生態(tài)站, 元江 653300 3 中國(guó)科學(xué)院大學(xué),北京 100049
稀樹(shù)灌草叢作為干熱河谷區(qū)特殊的植被類(lèi)型,其碳儲(chǔ)量等一直缺乏必要的研究。以元江干熱河谷稀樹(shù)灌草叢植被為對(duì)象,利用典型樣地法研究該區(qū)稀樹(shù)灌草叢植被的碳儲(chǔ)量與凈初級(jí)生產(chǎn)力。結(jié)果表明:元江稀樹(shù)灌草叢植被的碳儲(chǔ)量為32.13 t C/hm2,其中喬木、灌木和草本各層次的碳儲(chǔ)量為26.70、4.04、1.40 t C/hm2,分別占到總碳儲(chǔ)量的83.02%、12.57%、4.4%。喬木層中地上部分碳儲(chǔ)量占到66.70%。另外,元江稀樹(shù)灌草叢的凈初級(jí)生產(chǎn)力為3.88 t C hm-2a-1,其中林分的凈初級(jí)生產(chǎn)力為1.90 t C hm-2a-1,凋落物量為1.98 t C hm-2a-1;林下植被層對(duì)林分凈初級(jí)生產(chǎn)力的貢獻(xiàn)達(dá)到了46.92%。說(shuō)明元江稀樹(shù)灌草叢具有較高的碳儲(chǔ)量和碳匯能力。結(jié)果為稀樹(shù)灌草叢碳循環(huán)及碳匯功能研究提供了基礎(chǔ),同時(shí)也為干熱河谷區(qū)植被的保育與可持續(xù)經(jīng)營(yíng)提供了科學(xué)依據(jù)。
碳儲(chǔ)量;生產(chǎn)力;稀樹(shù)灌草叢;干熱河谷
Abstract: Savanna is a unique vegetation type that predominantly occurs in dry-hot valleys, yet its carbon storage capacity and net primary productivity (NPP) remain poorly understood. In the present study, we estimated carbon storage and NPP of savanna vegetation using permanent field plots in the dry-hot valley of Yuanjiang, Yunnan Province, southwestern China. We estimated tree carbon storage using allometric equations, and carbon storage in shrubs and herbs was estimated by the harvesting method. The NPP was calculated using two years of data from plot surveys. We found that the total carbon storage of the savanna in Yuanjiang was 32.13 t C/hm2. Carbon storage in the tree, shrub, and herb layers was 26.70 t C/hm2, 4.04 t C/hm2, and 1.40 t C/hm2, which contributed 83.02%, 12.57%, and 4.35% of the total carbon storage, respectively. Aboveground carbon storage accounts for 66.70% of tree layer carbon storage. Similarly, the NPP of the savanna was 3.88 t C hm-2a-1. NPP from the forest biomass and litter was 1.90 t C hm-2a-1and 1.98 t C hm-2a-1, respectively. The contribution to the NPP of forest biomass from the understory (shrubs and herbs) approached 46.92%. Thus, we conclude that, owing to its high carbon storage and NPP values, savanna vegetation is an important component of the carbon cycle; this finding provides a rationale for the conservation of vegetation and the sustainable management of savanna ecosystems in dry-hot valleys.
KeyWords: carbon storage; NPP; savanna; dry-hot valley
森林固碳能力或碳儲(chǔ)量是衡量森林生態(tài)功能的關(guān)鍵指標(biāo),也是研究碳循環(huán)的基礎(chǔ)。尤其是在全球變暖大背景下,森林在調(diào)節(jié)全球碳平衡的主導(dǎo)地位顯得尤為突出。然而,近年來(lái)針對(duì)森林碳儲(chǔ)量的研究主要集中于喬木林和灌木林,對(duì)不同林型、不同起源林分的碳儲(chǔ)量及生產(chǎn)力進(jìn)行了大量研究[1- 3]。但是,稀樹(shù)灌草叢(薩王納,Savanna)作為一類(lèi)特殊分布的森林類(lèi)群,其碳儲(chǔ)量及生產(chǎn)力受到的關(guān)注較少。另外,稀樹(shù)灌草叢植被受氣候變化的影響較為敏感。根據(jù)Zhao等[4]的模型研究認(rèn)為受到氣候變化的影響,我國(guó)稀樹(shù)灌草叢植被的面積有增大的趨勢(shì),而且其分布的界限也正逐漸北移。因此,研究稀樹(shù)灌草叢碳儲(chǔ)量及分配特征顯得更為迫切。
分布于我國(guó)的稀樹(shù)灌草叢植被中,尤以元江干熱河谷的最具代表性[5]。但是,現(xiàn)有的研究主要針對(duì)元江稀樹(shù)灌木草叢的群落特征[6- 11]、植物生理[12- 14]等方面展開(kāi),而針對(duì)元江干熱河谷稀樹(shù)灌草叢的碳儲(chǔ)量及生產(chǎn)力的研究較少。
元江干熱河谷分布的稀樹(shù)灌草叢作為世界薩王納植被的重要組成部分,其植被的碳儲(chǔ)量及其分配特點(diǎn)是怎樣的,該植被類(lèi)型的生產(chǎn)力和碳匯能力又是如何,這些問(wèn)題都急需回答。因此,本研究選擇元江干熱河谷稀樹(shù)灌草叢群落為對(duì)象,旨在研究稀樹(shù)灌木草叢群落碳儲(chǔ)量及生產(chǎn)力特征,為該區(qū)碳儲(chǔ)量估算及碳匯功能評(píng)估提供基礎(chǔ),同時(shí)可為該區(qū)植被的保護(hù)提供理論。
1 研究區(qū)概況
研究地點(diǎn)位于中國(guó)科學(xué)院西雙版納熱帶植物園元江干熱河谷生態(tài)站(102°10′40″E,23°27′56″N),海拔553 m,該區(qū)植被以稀樹(shù)灌木草叢為代表,總體上植被覆蓋較好,喬木層一般較為低矮,主要優(yōu)勢(shì)種為厚皮樹(shù)(Lanneacoromandelica)、細(xì)基丸(Polyalthiacerasoides);灌木多以宿萼木(Strophioblachiafimbricalyx)、蝦子花(Woodfordiafruticosa)、清香木(Pistaciaweinmanniifolia)、霸王鞭(Euphorbiaroyleana)等為優(yōu)勢(shì),灌木成叢現(xiàn)象比較明顯,平均高度80 cm左右,蓋度40%;草本以扭黃茅(Heteropogoncontortus)為優(yōu)勢(shì),高度在10—60 cm之間,蓋度達(dá)50%以上。該區(qū)多年平均降雨量700—800 mm左右,年平均溫度24.9℃,年日照時(shí)數(shù)2292 h,年蒸發(fā)量2750 mm,總體氣候特點(diǎn)為降水量少,平均溫度高,日照充足,干濕季明顯,終年無(wú)霜[15]。生境中裸巖率高(達(dá)61.2%),土層較薄,土壤以燥紅土為主。
2 研究方法
2.1 樣方調(diào)查
2014年10月通過(guò)踏勘,并考慮林分的特點(diǎn)及代表性,在研究區(qū)內(nèi)林分特征及立地條件相對(duì)一致的地段選取了6塊典型樣地,樣地面積不低于600 m2(20 m×30 m),樣地情況見(jiàn)表1。將樣地劃分為100 m2(10 m×10 m)的喬木樣方,分別對(duì)喬木(樹(shù)高>3 m)進(jìn)行每木檢尺,測(cè)定每株的樹(shù)高、胸徑、冠幅。在樣地中隨機(jī)的設(shè)置3個(gè)灌木樣方,大小為2 m×2 m,布設(shè)于對(duì)角線上,測(cè)定灌木樣方的物種組成、灌木的平均基徑、平均高度等。另外,在3個(gè)灌木樣方的左上角,分別設(shè)置了1個(gè)草本樣方(1 m × 1 m),測(cè)定草本樣方的總蓋度、物種組成、平均高度等指標(biāo)。最后,采用收獲法分別對(duì)灌木、草本樣方中所有植物按器官(灌木分葉、枝干、根系三部分,草本分地上、地下兩部分)全部收獲,先將灌草地上部分齊地剪斷收獲;再用全挖法將灌草的根系全部挖出,由于該區(qū)土層較薄,挖掘深度直至巖層為止,然后將灌木和草本各部分器官分別進(jìn)行稱(chēng)鮮重,同時(shí)每個(gè)部分取150 g左右的樣品帶回實(shí)驗(yàn)室,在烘箱中用105℃殺青30min后,恒溫70℃下烘干至衡重后測(cè)定各部分器官的含水率,根據(jù)灌草各部分器官的含水率,將林下植被(灌木、草本)各部分的鮮重?fù)Q算為干重生物量。最終,將各部分器官的干重生物量統(tǒng)一乘以轉(zhuǎn)換系數(shù)0.5得到各部分的碳儲(chǔ)量(t C/hm2)。

表1 樣地基本概況
2.2 數(shù)據(jù)統(tǒng)計(jì)分析
喬木層生物量估算使用每木檢尺的樹(shù)高、胸徑數(shù)據(jù),通過(guò)異速生長(zhǎng)方程計(jì)算得到。考慮研究區(qū)域的氣候、植被、樹(shù)種組成及模型使用范圍,喬木層生物量估算選用的異速生長(zhǎng)方程[16]:
Wl=0.014×(D2H)0.693
(1)
Wb=0.015×(D2H)0.844
(2)
Ws=0.091×(D2H)0.816
(3)
Wr=0.038×(D2H)0.896
(4)
式中,Wl指葉生物量(kg),Wb指枝生物量(kg),Ws指莖干生物量(kg),Wr指根生物量(kg),D為喬木1.3 m處胸徑(cm),H為樹(shù)高(m)。將計(jì)算得到喬木、各部分生物量統(tǒng)一乘以轉(zhuǎn)換系數(shù)0.5得到各部分的碳儲(chǔ)量(t C/hm2)。
凈初級(jí)生產(chǎn)力指的是生物體在單位面積、單位時(shí)間內(nèi)有機(jī)物的凈生產(chǎn)量。本研究中凈初級(jí)生產(chǎn)力(Net primary productivity,NPP)使用林分中各組分碳儲(chǔ)量的年變化量(t C hm-2a-1)來(lái)表示,其計(jì)算公式為:
NPP=ΔWt+ΔWs+ΔWh+TL
(5)
式中,ΔWt、ΔWs、ΔWh分別指代喬木、灌木、草本碳儲(chǔ)量年凈增量,TL為凋落物年掉落量。喬木的增量依據(jù)2a的群落調(diào)查數(shù)據(jù)通過(guò)模型計(jì)算得到,灌木、草本的增量依據(jù)2a通過(guò)全挖法的數(shù)據(jù)計(jì)算得到;凋落物量(TL)通過(guò)凋落物筐收集得到,乘以轉(zhuǎn)換系數(shù)0.5得到凋落物的碳儲(chǔ)量。
3 結(jié)果與分析
3.1 稀樹(shù)灌草叢喬木層碳儲(chǔ)量
稀樹(shù)灌草叢群落喬木層的總碳儲(chǔ)量為(26.68±0.88) t C/hm2(圖1)。喬木層11種木本植物的碳儲(chǔ)量在(0.05±0.03) —(16.47±2.01) t C/hm2之間,物種間碳儲(chǔ)量最大值比最小值高出339倍。碳儲(chǔ)量最大的物種是厚皮樹(shù),為16.47 t C/hm2,占到喬木層總碳儲(chǔ)量的61.71%,其次為霸王鞭,它的碳儲(chǔ)量為5.58 t C/hm2,占到20.90%;喬木層中碳儲(chǔ)量最低的物種為疏序黃荊(Vitexnegundoform.laxipaniculata),為0.04 t hm-2,僅占喬木層總碳儲(chǔ)量的0.18%。這表明喬稀樹(shù)灌草叢喬木層中優(yōu)勢(shì)物種比較突出,碳儲(chǔ)量在各物種間分布不均勻。
另外,喬木層碳儲(chǔ)量中地上部分(葉、枝、莖)的總碳儲(chǔ)量為(17.80±0.64) t C/hm2,占總碳儲(chǔ)量的66.70%,地下部分碳儲(chǔ)量為(8.89±0.26) t C/hm2,占到33.30%。而且,從地上部分碳儲(chǔ)量上看,枝和莖的碳儲(chǔ)量(17.06 t C/hm2)就占到了95.79%,但葉的生物量?jī)H為0.54 t C/hm2,只占到3.05%,減少了葉片中的碳儲(chǔ)量分配。
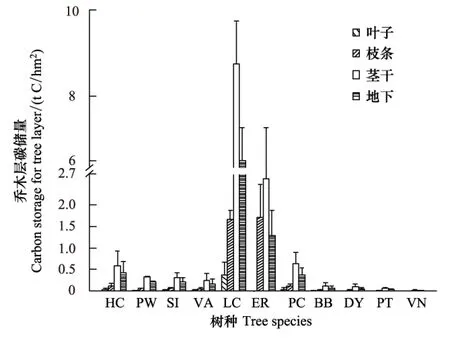
圖1 元江干熱河谷稀樹(shù)灌草叢喬木層碳儲(chǔ)量(均值+標(biāo)準(zhǔn)誤) Fig.1 Carbon storage of tree layer for Savanna in Yuanjiang dry-hot valley (Values represent means ± SE)HC:心葉木Haldina cordifolia;PW:清香木Pistacia weinmanniifolia;SI:異序?yàn)蹊闟apium insigne;VA:深裂山葡萄Vitis amurensis var. dissecta;LC:厚皮樹(shù)Lannea coromandelica;ER:霸王鞭Euphorbia royleana;PC:老人皮Polyalthia cerasoides;BB:鞍葉羊蹄甲Bauhinia brachycarpa;DY:云南柿Diospyros yunnanensis;PT:余甘子Phyllanthus taxodiifolius;VN:疏序黃荊Vitex negundo form. laxipaniculata
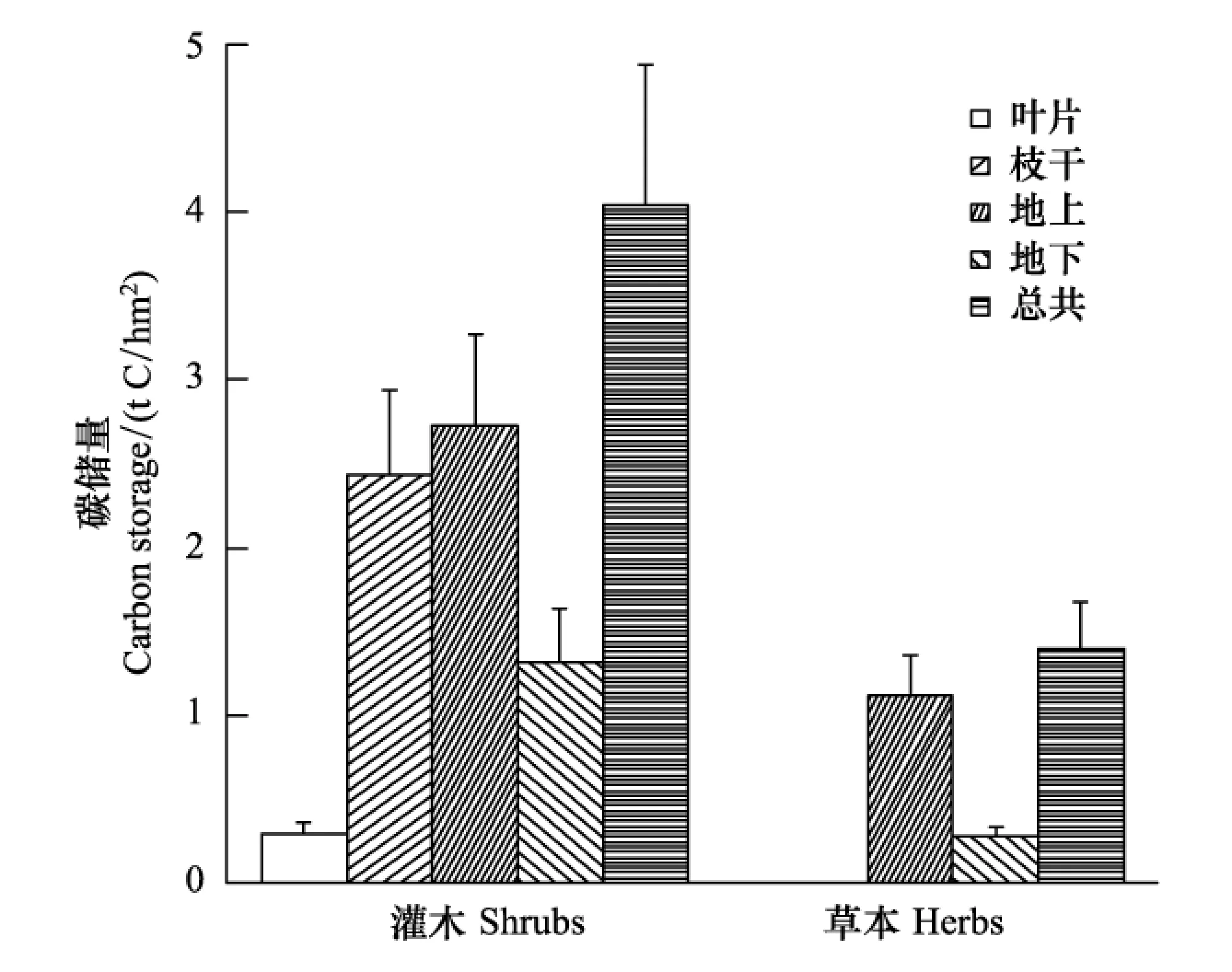
圖2 元江干熱河谷稀樹(shù)灌草叢林下植被碳儲(chǔ)量(均值+標(biāo)準(zhǔn)誤) Fig.2 Carbon storage of understory layer for Savanna in Yuanjiang dry-hot valley (mean+SE)
3.2 稀樹(shù)灌草叢林下植被碳儲(chǔ)量
元江稀樹(shù)灌草叢林下植被層總碳儲(chǔ)量為(5.44±0.99) t C/hm2,其中灌木層的總碳儲(chǔ)量為(4.04±0.84) t C/hm2,占林下植被層總碳儲(chǔ)量的74.32%,草本層碳儲(chǔ)量為(1.40±0.28) t C/hm2,占到25.70%(圖2)。
具體看來(lái),灌木層地上部分的葉、枝干碳儲(chǔ)量0.29、2.43 t C/hm2,分別占到灌木層碳儲(chǔ)量的7.21%、60.12%。而灌木層的地下碳儲(chǔ)量為(1.32±0.32) t C/hm2,占灌木層碳儲(chǔ)量的32.67%。草本層地上部分碳儲(chǔ)量為(1.12±0.24) t C/hm2,占草本層碳儲(chǔ)量的79.89%;地下部分碳儲(chǔ)量為(0.28±0.05) t C/hm2,僅占草本層碳儲(chǔ)量的20.04%。可知稀樹(shù)灌草叢林下植被碳儲(chǔ)量主要分配在地上部分。
3.3 稀樹(shù)灌草叢林分凈初級(jí)生產(chǎn)力
元江稀樹(shù)灌草叢凈初級(jí)生產(chǎn)力為3.88 t C hm-2a-1,其中林分的凈初級(jí)生產(chǎn)力為1.90 t C hm-2a-1,凋落物量為1.98 t C hm-2a-1(表2)。從林分的凈初級(jí)生產(chǎn)力看,地上部分為1.33 t C hm-2a-1,占到了林分總凈初級(jí)生產(chǎn)力的70.04%;而林分地下部分的凈初級(jí)生產(chǎn)力是0.57 t C hm-2a-1,占到林分的29.96%。表明林分中地上凈初級(jí)部分生產(chǎn)力更為明顯,當(dāng)然,29.96%比率也顯示了地下部分的凈初級(jí)生產(chǎn)力所占比重較大,凸顯了稀樹(shù)灌草叢的特征。
從林分的不同層次上看,林分中喬木層凈初級(jí)生產(chǎn)力為1.01 t C hm-2a-1、灌木層0.56 t C hm-2a-1、草本層0.33 t C hm-2a-1,分別占到林分總凈初級(jí)生產(chǎn)力的53.08%、29.49%、17.43%。而林分中林下植被層(灌木層和草本層)的凈初級(jí)生產(chǎn)力達(dá)到了46.92%,接近于喬木層的凈初級(jí)生產(chǎn)力,說(shuō)明在稀樹(shù)灌草叢中林下植被層的凈初級(jí)生產(chǎn)力對(duì)生態(tài)系統(tǒng)凈初級(jí)生產(chǎn)力的貢獻(xiàn)不可忽視。
4 討論
4.1 稀樹(shù)灌草叢碳儲(chǔ)量
森林生態(tài)系統(tǒng)作為一個(gè)動(dòng)態(tài)的碳庫(kù),準(zhǔn)確估計(jì)森林的碳儲(chǔ)量是正確評(píng)價(jià)森林碳匯功能的基礎(chǔ)[17]。元江稀樹(shù)灌草叢林分碳儲(chǔ)量為32.13 t C/hm2,在喬、灌、草3個(gè)層次中,稀樹(shù)灌草叢往地下部分分配了較多的碳儲(chǔ)量(均占到總碳儲(chǔ)量的30%以上),反映了稀樹(shù)灌草叢地下根系統(tǒng)較為發(fā)達(dá)。水分是限制稀樹(shù)灌草叢生態(tài)系統(tǒng)生長(zhǎng)的關(guān)鍵因子[15],發(fā)達(dá)的根系有利于該植被中植物能從更深更廣的區(qū)域獲取水分,這是對(duì)干旱脅迫的一種適應(yīng)[18-19]。相反,喬木層和灌木層中葉碳儲(chǔ)量的分配較少,其在總碳儲(chǔ)量中的比例不足10%,由于元江干熱河谷水熱矛盾突出,較少的葉碳儲(chǔ)量分配說(shuō)明植物葉片的葉面積及葉數(shù)量的減少,葉片也變薄[12],這有利于減少水分的散失,是適應(yīng)干旱的一種策略[20]。
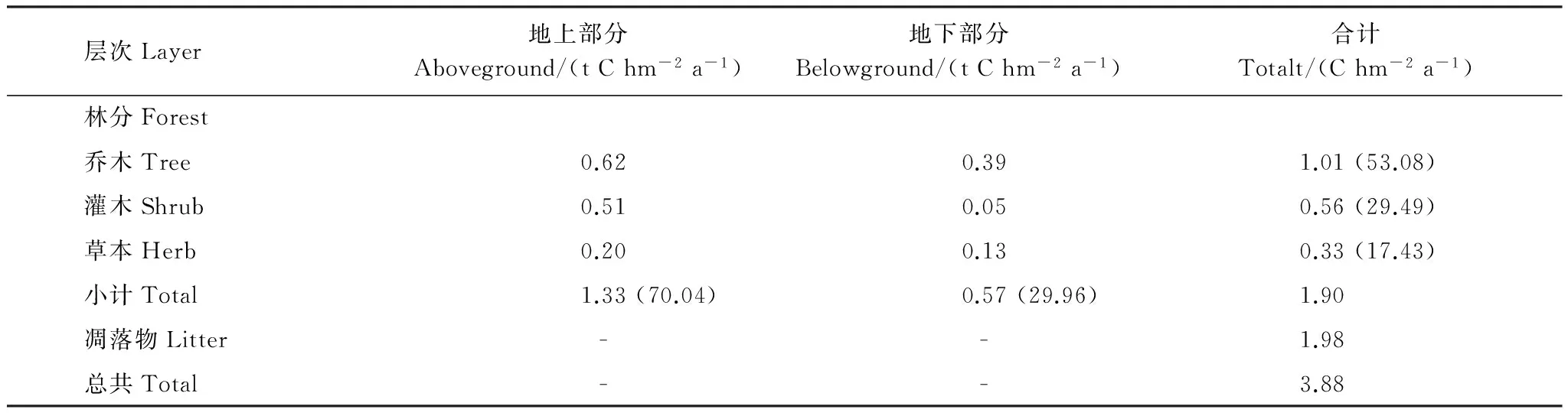
表2 元江干熱河谷稀樹(shù)灌草叢群落凈初級(jí)生產(chǎn)力
括號(hào)內(nèi)數(shù)字指代邏輯單元占林分總生產(chǎn)力的百分比
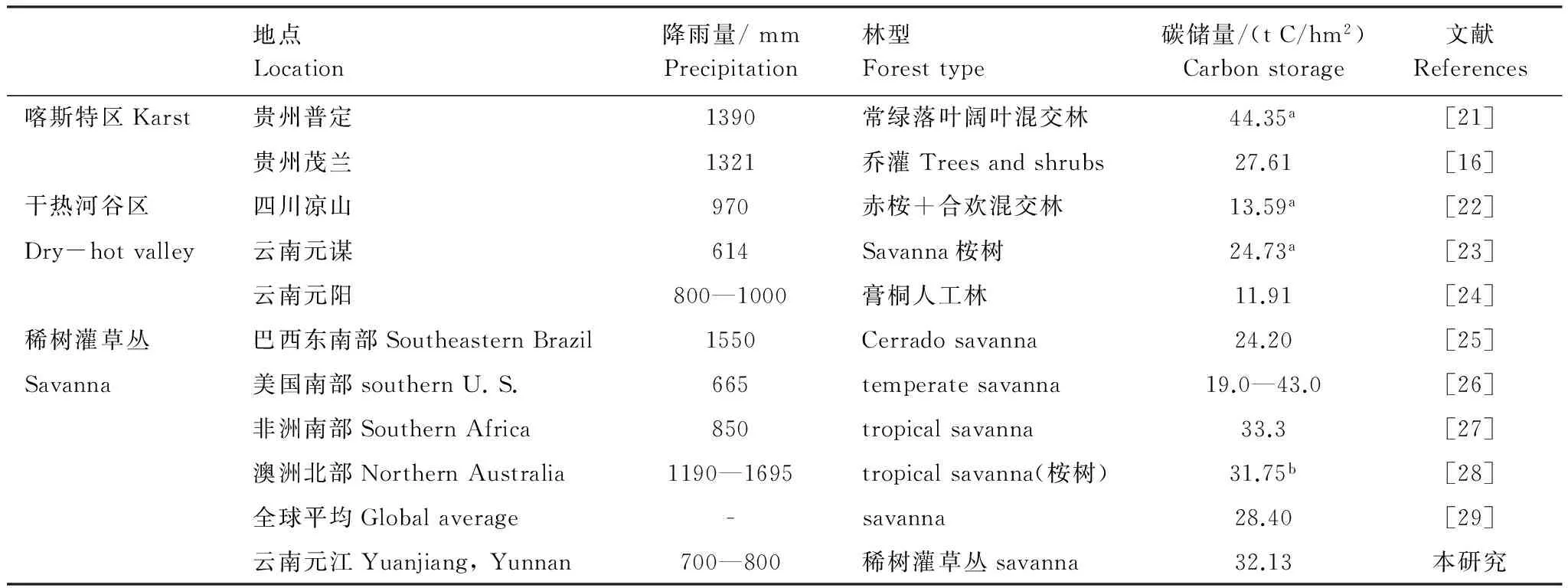
表3 不同森林類(lèi)型間碳儲(chǔ)量的差異
碳儲(chǔ)量一列中上標(biāo)a表示該值由生物量乘以轉(zhuǎn)換系數(shù)0.5得到;b表示該值為地上部分碳儲(chǔ)量
從同一地區(qū)來(lái)看,位于哀牢山河谷的元江干熱河谷稀樹(shù)灌草叢的碳儲(chǔ)量(32.13 t C/hm2)要遠(yuǎn)低于哀牢山山頂?shù)貐^(qū)中山濕性常綠闊葉林的碳儲(chǔ)量(257.90 t C/hm2)[30],這主要是因?yàn)榘Ю紊缴巾斁哂休^好的水分條件(降雨量1879.5 mm)和土壤狀況,森林生長(zhǎng)茂密,樹(shù)高可達(dá)20 m以上;而元江干熱河谷稀樹(shù)灌草叢分布于較為惡劣的石生環(huán)境中(裸巖率達(dá)到61.6%),嚴(yán)重的限制了植被的生長(zhǎng),植被一般較為低矮(喬木層平均樹(shù)高4.0 m左右,表1),制約了元江干熱河谷稀樹(shù)灌草叢植被的碳積累。
元江干熱河谷稀樹(shù)灌草叢植被的生長(zhǎng)環(huán)境與喀斯特區(qū)灌木林生長(zhǎng)環(huán)境極為相似,都生長(zhǎng)于石生環(huán)境中。比較兩種生境中植被的碳儲(chǔ)量發(fā)現(xiàn),元江稀樹(shù)灌草叢的碳儲(chǔ)量為32.13 t C/hm2,稍高于貴州茂蘭喀斯特區(qū)灌木林的碳儲(chǔ)量(27.61 t C/hm2)[16],但低于貴州普定喀斯特次生喬木林的碳儲(chǔ)量(44.35 t C/hm2)[21]。這說(shuō)明,元江干熱河谷稀樹(shù)灌草叢生態(tài)系統(tǒng)碳儲(chǔ)量與喀斯特區(qū)次生喬木林的碳儲(chǔ)量相當(dāng)。雖然,在貴州喀斯特區(qū)平均年降雨量較為充足(1200 mm),然而生境惡劣及土層淺薄也是限制喀斯特植被的生長(zhǎng)的重要因素[16]。
與國(guó)內(nèi)其他干熱河谷區(qū)植被相比,元江干熱河谷稀樹(shù)灌草叢的碳儲(chǔ)量要高(表3)。主要原因是在金沙江干熱河谷水土流失嚴(yán)重,土層中養(yǎng)分流失嚴(yán)重,限制了植被的發(fā)育,再加上該區(qū)多為人工林分,人為干擾嚴(yán)重,原生植被破壞嚴(yán)重,影響植被的碳積累[11, 23-24]。相反,元江干熱河谷稀樹(shù)灌草叢群落發(fā)育相對(duì)完善,可以明顯分出喬、灌、草三層[6, 11],有利于植被的保護(hù)及碳積累。
而與其它區(qū)域稀樹(shù)灌草叢相比,元江干熱河谷稀樹(shù)灌草叢碳儲(chǔ)量(32.12 t C/hm2)要稍高于全球稀樹(shù)灌草叢主要分布區(qū)的平均碳儲(chǔ)量(28.40 t C/hm2)(表3)和巴西東南部稀樹(shù)灌草叢(Cerrado savanna)的24.20 t C/hm2,而與美國(guó)南部(19.0—43.0 t C/hm2)、非洲南部(33.3 t C/hm2)、澳洲北部(31.75 t C/hm2)稀樹(shù)灌草叢的碳儲(chǔ)量比較接近。這表明該區(qū)稀樹(shù)灌草叢植被的碳儲(chǔ)量與世界其它稀樹(shù)灌草叢分布區(qū)域的碳儲(chǔ)量是相當(dāng)?shù)?其碳儲(chǔ)量不容忽視,應(yīng)得到重視與保護(hù)。
4.2 凈初級(jí)生產(chǎn)力
元江干熱河谷稀樹(shù)灌草叢總的凈初級(jí)生產(chǎn)力為3.88 t C hm-2a-1,其中林分的凈初級(jí)生產(chǎn)力為1.90 t C hm-2a-1,林分中林下植被的凈初級(jí)生產(chǎn)力在林分凈初級(jí)生產(chǎn)力的貢獻(xiàn)達(dá)到了47%,接近喬木層凈初級(jí)生產(chǎn)力所占比例(53%)(表2)。由于上層喬木通過(guò)控制林內(nèi)小氣候與資源利用率影響著林下植被的組成與生長(zhǎng)[31-32],元江稀樹(shù)灌草叢喬木層較為開(kāi)闊的林冠(Open-canopy,郁閉度0.5左右),為林下植被層獲取光資源和空間生態(tài)位的占領(lǐng)提供了便利,同時(shí)林下植物的多分枝(或分蘗)現(xiàn)象也會(huì)提高植物的生產(chǎn)力[33],提高了植物的固碳能力。
本研究中稀樹(shù)灌草叢總的凈初級(jí)生產(chǎn)力要低于喀斯特森林的凈初級(jí)生產(chǎn)力(4.34 t C hm-2a-1)[34]。相比之下,元江稀樹(shù)灌草叢的凈初級(jí)生產(chǎn)力是較低的,然而,元江干熱河谷植被降水偏少,土層淺薄等嚴(yán)酷環(huán)境會(huì)影響植物的發(fā)育,間接影響到植被的固碳潛力[35]。與其它區(qū)域稀樹(shù)灌草叢相比,研究區(qū)稀樹(shù)灌草叢的凈初級(jí)生產(chǎn)力僅達(dá)到全球平均值(7.2 t C hm-2a-1)[29]的一半左右。這主要是因?yàn)镚race等[29]在綜合全球稀樹(shù)灌草叢凈初級(jí)生產(chǎn)力的數(shù)據(jù)中,印度稀樹(shù)灌草叢的凈初級(jí)生產(chǎn)力普遍偏高(最高值達(dá)到22.8 t C hm-2a-1),拉高了全球的平均值,而印度這些稀樹(shù)灌草叢多為熱帶雨林退化形成,其主要樹(shù)種貢獻(xiàn)了較高生產(chǎn)力。而與其它類(lèi)型稀樹(shù)灌草叢凈初級(jí)生產(chǎn)力相比,元江稀樹(shù)灌草叢的凈初級(jí)生產(chǎn)力處于要遠(yuǎn)低于澳洲北部桉樹(shù)為主要優(yōu)勢(shì)種的稀樹(shù)灌草叢(11.0 t C hm-2a-1)[36],由于桉樹(shù)作為一種速生樹(shù)種,具有較高的生產(chǎn)力,再加上澳洲北部較好的水分條件(平均年降雨量1200—1400 mm)促進(jìn)了澳洲稀樹(shù)灌草叢較高的生產(chǎn)力;其值要高于非洲西部(2.2 t C hm-2a-1)[37]和美國(guó)南部(2.22 t C hm-2a-1)[26]稀樹(shù)灌草叢的凈初級(jí)生產(chǎn)力,更接近于南美洲稀樹(shù)灌草叢的凈初級(jí)生產(chǎn)力(3.9 t C hm-2a-1)[38]。通過(guò)比較可以看出,元江稀樹(shù)灌草叢具有一定的凈初級(jí)生產(chǎn)力,其固碳潛力是不可忽視的。
5 結(jié)論
(1)元江稀樹(shù)灌草叢植被的碳儲(chǔ)量為32.13 t C/hm2。喬木層碳儲(chǔ)量為26.70 t C/hm2,主要樹(shù)種間碳儲(chǔ)量的分配不均勻。
(2)林下植被層碳儲(chǔ)量為5.44 t C/hm2,其中灌木占74.3%,草本層碳儲(chǔ)量占25.7%。
(3)元江稀樹(shù)灌草叢植被總的凈初級(jí)生產(chǎn)力為3.88 t C hm-2a-1。林分中林下植被層的凈初級(jí)生產(chǎn)力達(dá)到46.92%。林分的地上部分凈初級(jí)生產(chǎn)力所占比重較大(70.04%),但地下部分凈初級(jí)生產(chǎn)力(29.96%)也較為明顯。
(4)元江干熱河谷區(qū)稀樹(shù)灌草叢植被的碳儲(chǔ)量和固碳潛力不可忽視,理應(yīng)受到重視與保護(hù)。
致謝:中國(guó)科學(xué)院西雙版納熱帶植物園元江干熱河谷生態(tài)站和中心實(shí)驗(yàn)室大力協(xié)助,刀萬(wàn)有、封乾元、刀新明幫助凋落物收集,特此致謝。
[1] 胡會(huì)峰, 王志恒, 劉國(guó)華, 傅伯杰. 中國(guó)主要灌叢植被碳儲(chǔ)量 植物生態(tài)學(xué)報(bào), 2006, 30(4): 539- 544.
[2] Zhang Y D, Gu F X, Liu S R, Liu Y C, Li C. Variations of carbon stock with forest types in subalpine region of southwestern China. Forest Ecology and Management, 2013, 300: 88- 95.
[3] Fang J Y, Chen A P, Peng C H, Zhao S Q, Ci L J. Changes in forest biomass carbon storage in China between 1949 and 1998. Science, 2001, 292(5525): 2320- 2322.
[4] Zhao D S, Wu S H. Responses of vegetation distribution to climate change in China. Theoretical and Applied Climatology, 2014, 117(1/2): 15- 28.
[5] 《云南植被》編寫(xiě)組. 云南植被. 北京: 科學(xué)出版社, 1987.
[6] 金振洲. 云南元江干熱河谷半薩王納植被的植物群落學(xué)研究. 廣西植物, 1999, 19(4): 289- 302.
[7] 劉方炎, 朱華. 元江干熱河谷植被數(shù)量分類(lèi)及其多樣性分析. 廣西植物, 2005, 25(1): 22- 25.
[8] 劉方炎, 朱華, 施濟(jì)普, 陳曉鳴. 元江干熱河谷植物群落特征及土壤肥力研究. 應(yīng)用與環(huán)境生物學(xué)報(bào), 2007, 13(6): 782- 787.
[9] 沈蕊, 張建利, 何彪, 李峰, 張志明, 周睿, 歐曉昆. 元江流域干熱河谷草地植物群落結(jié)構(gòu)特征與相似性分析. 生態(tài)環(huán)境學(xué)報(bào), 2010, 19(12): 2821- 2825.
[10] 朱華. 元江干熱河谷肉質(zhì)多刺灌叢的研究. 云南植物研究, 1990, 12(3): 301- 310.
[11] 金振洲, 歐曉昆. 干熱河谷植被: 元江、怒江、金沙江、瀾滄江. 昆明: 云南大學(xué)出版社, 云南科技出版社, 2000.
[12] 宋富強(qiáng), 曹坤芳. 元江干熱河谷植物葉片解剖和養(yǎng)分含量特征. 應(yīng)用生態(tài)學(xué)報(bào), 2005, 16(1): 33- 38.
[13] 張樹(shù)斌, 張教林, 曹坤芳. 干旱對(duì)干熱河谷優(yōu)勢(shì)木本植物蝦子花光能分配的影響. 云南大學(xué)學(xué)報(bào): 自然科學(xué)版, 2014, 36(5): 774- 780.
[14] Zhang J L, Zhu J J, Cao K F. Seasonal variation in photosynthesis in six woody species with different leaf phenology in a valley savanna in southwestern China. Trees, 2007, 21(6): 631- 643.
[15] 張一平, 段澤新, 竇軍霞. 岷江上游干暖河谷與元江干熱河谷的氣候特征比較研究. 長(zhǎng)江流域資源與環(huán)境, 2005, 14(1): 76- 82.
[16] 黃宗勝, 喻理飛, 符裕紅, 楊瑞. 茂蘭退化喀斯特森林植被自然恢復(fù)中生態(tài)系統(tǒng)碳吸存特征. 植物生態(tài)學(xué)報(bào), 2015, 39(6): 554- 564.
[17] 楊洪曉, 吳波, 張金屯, 林德榮, 常順利. 森林生態(tài)系統(tǒng)的固碳功能和碳儲(chǔ)量研究進(jìn)展. 北京師范大學(xué)學(xué)報(bào): 自然科學(xué)版, 2005, 41(2): 172- 177.
[18] Santiago L S, Bonal D, De Guzman M E,vila-Lovera E. Drought survival strategies of tropical trees // Goldstein G, Santiago L S, eds. Tropical tree physiology: adaptations and responses in a changing environment. Cham: Springer International Publishing, 2016: 243- 258.
[19] Brunner I, Herzog C, Dawes M A, Arend M, Sperisen C. How tree roots respond to drought. Frontiers in Plant Science, 2015, 6: 547.
[20] Farooq M, Wahid A, Kobayashi N, Fujita D, Basra S M A. Plant drought stress: effects, mechanisms and management. Agronomy for Sustainable Development, 2009, 29(1): 185- 212.
[21] 劉長(zhǎng)成, 魏雅芬, 劉玉國(guó), 郭柯. 貴州普定喀斯特次生林喬灌層地上生物量. 植物生態(tài)學(xué)報(bào), 2009, 33(4): 698- 705.
[22] 林偉宏, 陳克明, 劉照光. 川西南干熱河谷赤按人工林生物量和營(yíng)養(yǎng)元素含量. 山地研究, 1994, 12(4): 251- 255.
[23] 楊忠, 張建平, 王道杰, 張信寶. 元謀干熱河谷桉樹(shù)人工林生物量初步研究. 山地學(xué)報(bào), 2001, 19(6): 503- 510.
[24] 郭永清. 云南干熱河谷膏桐人工林生態(tài)系統(tǒng)碳庫(kù)特征[D]. 南京: 南京林業(yè)大學(xué), 2010.
[25] Lilienfein J, Wilcke W. Element storage in native, agri-, and silvicultural ecosystems of the Brazilian savanna. Plant and Soil, 2003, 254(2): 425- 442.
[26] Hughes R F, Archer S R, Asner G P, Wessman C A, McMurtry C, Nelson J, Ansley R J. Changes in aboveground primary production and carbon and nitrogen pools accompanying woody plant encroachment in a temperate savanna. Global Change Biology, 2006, 12(9): 1733- 1747.
[27] Ryan C M, Williams M, Grace J. Above- and belowground carbon stocks in a miombo woodland landscape of Mozambique. Biotropica, 2011, 43(4): 423- 432.
[28] Scheiter S, Higgins S I, Beringer J, Hutley L B. Climate change and long-term fire management impacts on Australian savannas. New Phytologist, 2015, 205(3): 1211- 1226.
[29] Grace J, José J S, Meir P, Miranda H S, Montes R A. Productivity and carbon fluxes of tropical savannas. Journal of Biogeography, 2006, 33(3): 387- 400.
[30] 張鵬超, 張一平, 楊國(guó)平, 鄭征, 劉玉洪, 譚正洪. 哀牢山亞熱帶常綠闊葉林喬木碳儲(chǔ)量及固碳增量. 生態(tài)學(xué)雜志, 2010, 29(6): 1047- 1053.
[31] Augusto L, Dupouey J L, Ranger J. Effects of tree species on understory vegetation and environmental conditions in temperate forests. Annals of Forest Science, 2003, 60(8): 823- 831.
[32] Zapfack L, Engwald S, Sonke B, Achoundong G, Madong B A. The impact of land conversion on plant biodiversity in the forest zone of Cameroon. Biodiversity & Conservation, 2002, 11(11): 2047- 2061.
[33] Sah J P, Ross M S, Koptur S, Snyder J R. Estimating aboveground biomass of broadleaved woody plants in the understory of Florida Keys pine forests. Forest Ecology and Management, 2004, 203(1/3): 319- 329.
[34] 于維蓮, 董丹, 倪健. 中國(guó)西南山地喀斯特與非喀斯特森林的生物量與生產(chǎn)力比較. 亞熱帶資源與環(huán)境學(xué)報(bào), 2010, 15(2): 25- 30.
[35] 田漢勤, 徐小鋒, 宋霞. 干旱對(duì)陸地生態(tài)系統(tǒng)生產(chǎn)力的影響. 植物生態(tài)學(xué)報(bào), 2007, 31(2): 231- 241.
[36] Chen X Y, Hutley L B, Eamus D. Carbon balance of a tropical savanna of northern Australia. Oecologia, 2003, 137(3): 405- 416.
[37] Hanan N P, Kabat P, Dolman A J, Elbers J A. Photosynthesis and carbon balance of a Sahelian fallow savanna. Global Change Biology, 1998, 4(5): 523- 538.
[38] San José J J, Montes R A. Management effects on carbon stocks and fluxes across the Orinoco savannas. Forest Ecology and Management, 2001, 150(3): 293- 311.
Carbonstorageandnetprimaryproductivityofasavannaecosysteminadry-hotvalleyinYuanjiang,Yunnan
JIN Yanqiang1,3, LI Jing1,3, ZHANG Yiping1,*, LIU Yuntong1, FEI Xuehai1,3, LI Peiguang2
1KeyLaboratoryofTropicalForestEcology,XishuangbannaTropicalBotanicalGarden,ChineseAcademyofSciences,Kunming650223,China2YuanjiangSavannaEcosystemResearchStation,XishuangbannaTropicalBotanicalGarden,ChineseAcademyofSciences,Yuanjiang653300,China3UniversityofChineseAcademyofSciences,Beijing100049,China
國(guó)家自然科學(xué)基金與云南省聯(lián)合項(xiàng)目(U1202234);國(guó)家基金重大項(xiàng)目(31290221);國(guó)家基金青年項(xiàng)目(41405143);云南省基金面上項(xiàng)目(2015FB186)
2016- 06- 01; < class="emphasis_bold">網(wǎng)絡(luò)出版日期
日期:2017- 04- 24
10.5846/stxb201606011053
*通訊作者Corresponding author.E-mail: yipingzh@xtbg.ac.cn
金艷強(qiáng),李敬,張一平,劉運(yùn)通,費(fèi)學(xué)海,李培廣.元江干熱河谷稀樹(shù)灌草叢植被碳儲(chǔ)量及凈初級(jí)生產(chǎn)力.生態(tài)學(xué)報(bào),2017,37(17):5584- 5590.
Jin Y Q, Li J, Zhang Y P, Liu Y T, Fei X H, Li P G.Carbon storage and net primary productivity of a savanna ecosystem in a dry-hot valley in Yuanjiang, Yunnan.Acta Ecologica Sinica,2017,37(17):5584- 5590.
