油酸誘導單純性肝脂肪變性細胞模型的建立及應用
2017-11-01 22:00:16董麗紅張瑞芬魏振承張名位
中國藥理學通報 2017年11期
關鍵詞:模型
董麗紅,張瑞芬,黃 菲,魏振承,張名位
(廣東省農業科學院蠶業與農產品加工研究所,農業部功能食品重點實驗室,廣東省農產品加工重點實驗室,廣東 廣州 510610)
油酸誘導單純性肝脂肪變性細胞模型的建立及應用
董麗紅,張瑞芬,黃 菲,魏振承,張名位
(廣東省農業科學院蠶業與農產品加工研究所,農業部功能食品重點實驗室,廣東省農產品加工重點實驗室,廣東 廣州 510610)
目的探討人肝癌HepG2細胞單純肝脂肪變性細胞模型的建立方法與模型的應用。方法體外培養HepG2細胞,以不同濃度的油酸處理24 h誘導細胞脂肪變性,油紅O和尼羅紅染色定性觀察細胞內脂滴蓄積情況,并測定細胞內甘油三酯(TG)、丙二醛(MDA)、超氧化物歧化酶(SOD)以及培養液中谷丙轉氨酶(ALT)和谷草轉氨酶(AST)水平,確定最佳模型濃度;同時,利用此模型觀察黃酮類化合物對脂肪變性肝細胞內TG的影響。結果油酸終濃度為0.4 mmol·L-1時,肝細胞內有大量脂滴形成,且細胞內TG含量明顯增加,而MDA含量、SOD活性及培養液中ALT、AST釋放量較正常組無明顯變化;槲皮素和表兒茶素可明顯降低模型細胞內TG含量。結論采用0.4 mmol·L-1油酸可成功誘導HepG2細胞發生脂肪變性,建立單純性肝細胞脂肪變性模型,該模型適用于預防非酒精性脂肪肝病的天然降脂藥物研究。
油酸;HepG2細胞;脂肪變性;甘油三脂;細胞模型;黃酮類化合物
非酒精性脂肪肝病(nonalcoholic fatty liver disease, NAFLD)是目前世界上廣泛流行的一種慢性疾病,其發病過程可由早期階段的單純性脂肪肝逐漸發展到非酒精性肝炎(nonalcoholic steatohepatitis, NASH),甚至肝纖維化和肝硬化等嚴重病癥。根據NAFLD形成機制的“二次打擊”學說,肝細胞內甘油三酯(triglyceride, TG)過度沉積是“第一次打擊”,導致單純性脂肪肝的發生;隨之,氧化應激和炎癥因子上演“第二次打擊”,促使NASH的發展[1]。可見,在無“第二次打擊”的情況下,單純性脂肪肝不一定會轉化成NASH,其臨床表現為肝臟單純性脂肪變性,是預防NAFLD發生和發展的最佳時期。鑒于膳食療法的預防和早期治療原則以及無毒副作用等優點,尋求新的食療方法,以預防肝脂肪變性及防止其向NASH的轉化成為食品營養學領域的研究熱點。因而,首先要建立一種方便、實用的單純性肝脂肪變性模型。目前,細胞模型由于實驗條件可控、影響因素少而被廣泛用于NAFLD的體外研究[2],但大部分研究采用的細胞模型并非單純性肝脂肪變性模型,如Yin等[3]和Xie等[4]的研究分別以一定濃度游離脂肪酸誘導L02或HepG2細胞脂肪變性,在TG蓄積的同時,谷丙轉氨酶(alanine aminotransferase, ALT)、谷草轉氨酶(aspartate aminotransferase, AST)、丙二醛(malondialdehyde, MDA)、超氧化物歧化酶(superoxide dismutase, SOD)等指標亦明顯變化,引起細胞氧化應激損傷。因此,本研究擬采用油酸誘導HepG2建立單純性肝脂肪變性細胞模型,并利用該細胞模型,評價黃酮類化合物預防肝脂肪變性的作用,旨在建立一種早期NAFLD的細胞模型,并應用于膳食營養或食品功能因子等活性物質預防NAFLD的體外研究。
1 材料與方法
1.1材料
1.1.1細胞 人肝癌細胞HepG2由中國科學院干細胞庫提供。
1.1.2試劑與儀器 MEM培養液、胎牛血清、非必需氨基酸溶液,購自美國Thermo Fisher公司;油酸(oleic acid,OA)、牛血清白蛋白(bull serum albumin, BSA)、油紅O、尼羅紅染料,購自美國Sigma公司;MTT細胞增殖-毒性檢測、ALT、AST、MDA、SOD測定試劑盒,購自南京建成生物工程研究所;TG酶法檢測試劑盒、BCA蛋白測定試劑盒,購自北京普利萊技術有限公司。熒光倒置相差顯微鏡購自德國Leica公司;Infinite M200pro熒光酶標儀購自瑞士Tecan公司。
1.2方法
1.2.1細胞培養 HepG2細胞用含10%胎牛血清、1%非必需氨基酸和抗生素的MEM培養液培養于T25細胞培養瓶中,置于37℃、5%的CO2培養箱中培養,觀察并記錄細胞的生長狀態。待細胞匯合度達到80%~90%時,用1 mL 0.25%胰酶-EDTA消化細胞,1 ∶3 傳代。取對數生長期的HepG2細胞進行實驗。
1.2.2細胞存活率的測定 取對數生長期的HepG2細胞以5×108·L-1,每孔100 μL接種于96孔細胞培養板,待細胞完全貼壁后,加入不同濃度的油酸(以異丙醇作溶劑[5]),每個濃度設置6個復孔。培養24、48 h后,按照MTT法檢測試劑盒的說明書測定各組細胞活力,篩選油酸的安全濃度范圍。以酶標儀測定570 nm處吸光度值A,計算細胞存活率/%=[(A實驗組-A空白組)/(A對照組-A空白組)]×100%
1.2.3油酸誘導肝細胞脂肪變性 取對數生長期的HepG2細胞以5×108·L-1,每孔2 mL接種于6孔細胞培養板,待細胞匯合至70%~80%時,棄去舊的培養液,用含1% BSA的無血清MEM培養基饑餓處理16 h后,加入不同濃度的油酸(“1.2.2”確定的安全濃度范圍內),每個濃度重復3次,培養24 h。
1.2.4單純性肝脂肪變性細胞模型的鑒定
1.2.4.1細胞內脂滴染色觀察 細胞按“1.2.3”方法做相應處理后,棄去培養液,用PBS清洗3次,冰凍的4%多聚甲醛固定10 min,分別采用油紅O染色法[3]和尼羅紅染色法[5]進行染色,熒光顯微鏡下觀察、拍照。
1.2.4.2TG、ALT、AST、MDA和SOD的測定 細胞按“1.2.3”方法做相應處理后,收集培養液和細胞。培養液1 000 r·min-1離心5 min,取上清,以試劑盒測定ALT、AST的含量;加入細胞裂解液裂解細胞,2 000 r·min-1離心5 min,取上清,以試劑盒測定TG、MDA、SOD濃度,并以BCA 法測定蛋白濃度,分別以每克細胞蛋白中所含的量來表示。
1.2.5模型的應用 選取“1.2.4”確定的最佳油酸濃度,按“1.2.3”所述的方法建立單純性肝細胞脂肪變性模型。設置正常對照組、油酸模型組和油酸+不同濃度藥物的實驗組(槲皮素或表兒茶素),每組重復3次。處理完成后收集細胞,以試劑盒測定細胞內TG的變化。
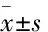
2 結果
2.1油酸對HepG2細胞活力的影響由Fig 1可知,相比于未加油酸的正常對照組,油酸濃度為0.8 mmol·L-1時,誘導24 h能明顯降低HepG2細胞活力(P<0.05),且當油酸濃度達到1.0 mmol·L-1時,存活細胞量明顯減少,僅為79%(P<0.01);誘導48 h,油酸對HepG2細胞活力的影響更大,其濃度為0.6 mmol·L-1時,HepG2細胞活力已明顯降低至正常對照組的90%(P<0.05)。由此可見,油酸濃度和誘導時間對HepG2細胞活力均有一定影響。油酸濃度≤0.6 mmol·L-1,孵育24 h時,對HepG2細胞活力無明顯影響。
Fig 1 Effect of oleic acid on HepG2 cell viability
*P<0.05,**P<0.01vscontrol group
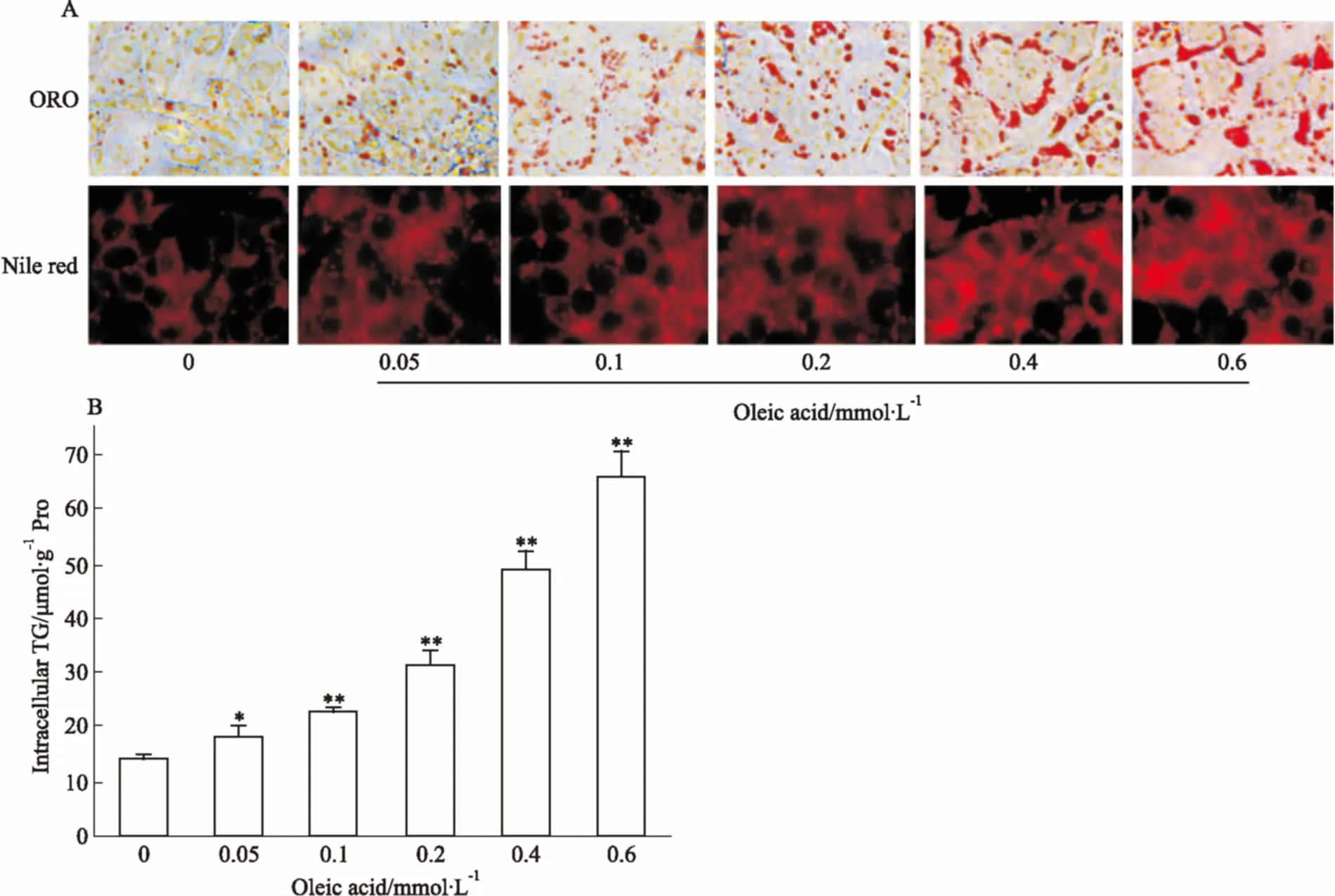
Fig2EffectofoleicacidonlipidaccumulationinHepG2cells
A:Oil red O staining and Nile red staining(×400); B:Intracellular TG content.*P<0.05,**P<0.01vscontrol group
2.2單純性肝脂肪變性細胞模型的鑒定
2.2.1油酸對HepG2細胞內脂肪沉積的影響 從Fig 2A可看出,對比2種脂質染色方法,均能很好地呈現油酸負荷下HepG2細胞內脂肪沉積的特征。隨著油酸濃度的增加,細胞內紅色脂滴越來越多,脂滴主要分布在細胞質內靠近細胞膜的部位,當其濃度達到0.4 mmol·L-1時,在靠近細胞膜的區域聚集大量的紅色脂滴且連成片狀。細胞內TG含量的測定結果與染色觀察結果一致(Fig 2B),隨著油酸濃度的增加,細胞內TG逐漸上升,呈劑量效應關系。油酸濃度為0.1 mmol·L-1時,細胞內TG含量比正常對照組中TG含量高出64%;當油酸濃度增加到0.4 mmol·L-1時,細胞內TG含量已達到正常對照組的3.5倍。表明油酸濃度為0.4 mmol·L-1時能明顯導致HepG2細胞內脂肪沉積。
2.2.2油酸對HepG2細胞氧化應激損傷的影響 由Fig 3A可知,相比于正常對照組,濃度≤0.4 mmol·L-1的油酸誘導24 h后,ALT和AST水平無明顯增加,當濃度達到0.6 mmol·L-1時,ALT水平明顯升高(P<0.05)。由Fig 3B、3C可知,相比于正常對照組,濃度≤0.4 mmol·L-1的油酸組細胞內MDA和SOD含量亦無明顯變化,而0.6 mmol·L-1油酸組細胞內MDA水平明顯增加(P<0.05),SOD活性明顯降低(P<0.05),表明高濃度油酸可引發HepG2細胞氧化應激反應,導致細胞氧化損傷,甚至死亡。
上述結果表明,油酸濃度為0.4 mmol·L-1時,細胞內聚集的甘油三酯明顯增加,且此濃度未引起細胞氧化應激損傷。因此,以0.4 mmol·L-1的油酸誘導HepG2細胞24 h可成功建立單純性肝脂肪變性細胞模型。
2.3黃酮類化合物對HepG2細胞內脂質聚集的影響以0.4 mmol·L-1油酸誘導HepG2細胞建立單純性肝細胞脂肪變性模型,在加入油酸的同時,給予不同濃度槲皮素或表兒茶素干預24 h,檢測各組細胞內TG的變化。結果如Fig 4所示,各濃度槲皮素和表兒茶素均能明顯降低細胞內TG含量(P<0.05),其中20 μmol·L-1時抑制效果最好,與油酸模型組相比,表兒茶素的抑制率為24%,槲皮素的抑制率達到45%。表明黃酮類化合物對單純性肝細胞脂肪變性具有良好的改善作用。
3 討論
建立肝脂肪變性細胞模型的關鍵在于選擇合適的細胞和誘導劑。大量研究發現,以油酸或軟脂酸誘導HepG2細胞脂肪變性是一種較合適的體外研究NAFLD模型,而其中關于單純性肝脂肪變性細胞模型的研究報道較少[2]。單純性脂肪變性的特征是細胞內TG蓄積,但無明顯氧化應激損傷,通常以肝酶ALT、AST以及MDA、SOD等脂質過氧化指標來判斷是否造成肝損傷[6]。而劉江等[7]僅通過模型組和正常組細胞內MDA無明顯差異,則認為所建模型是單純脂肪變性。殷錦錦等[8]通過細胞培養液中MDA變化來判斷模型是否存在脂質過氧化,這些研究均不能完全說明建立的模型是單純性脂肪變性模型。此外,有研究發現,不同游離脂肪酸誘導肝細胞脂肪變性時,軟脂酸更易造成細胞急性脂毒性,而油酸更易被細胞吸收而引起TG蓄積,這說明油酸比軟脂酸更適合用于建立單純性肝脂肪變性細胞模型[9]。
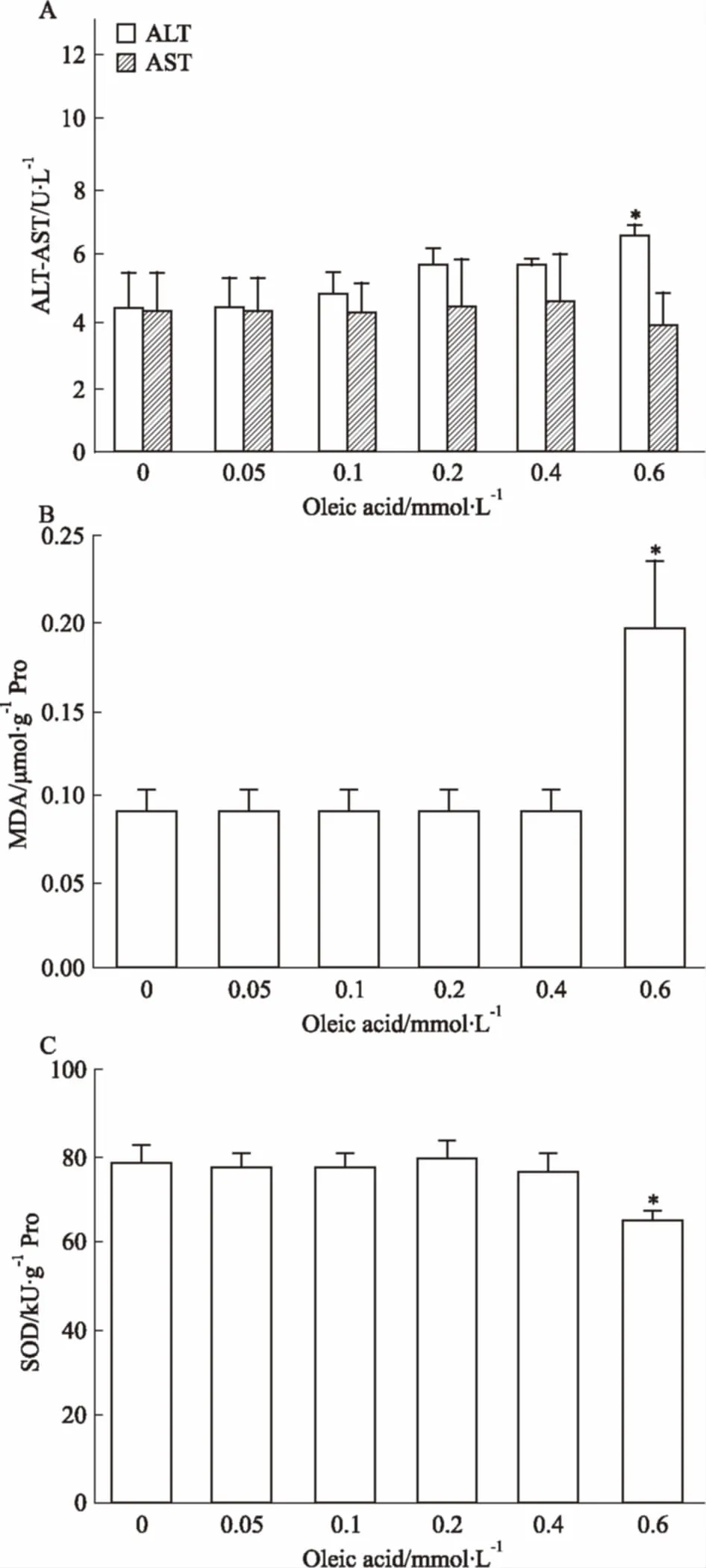
Fig 3 Effect of oleic acid on transaminaseactivities and oxidative injury in HepG2 cells
A: ALT and AST activity in cell culture medium; B:Intracellular MDA content; C: SOD activity.*P<0.05vscontrol group
本研究發現,以0.4 mmol·L-1油酸誘導HepG2細胞24 h后,細胞內TG含量明顯增加,表明脂肪變性細胞模型造模成功。同時測定發現,該模型組細胞培養液中ALT和AST水平無明顯增加,表明該模型無明顯的肝損傷;且細胞內MDA和SOD含量無明顯變化,表明該模型不會導致脂質過氧化,符合單純性脂肪變性模型的特征。Cui 等[10]的研究發現,油酸濃度不超過0.4 mmol·L-1時細胞活力無明顯影響,且細胞內TG含量呈劑量依賴增加,這與本研究結果一致。Liu等[11]的研究以1 mmol·L-1油酸誘導HepG2細胞24 h建立了單純性脂肪變性模型,這可能是由于油酸溶解的方法不同,此方法先用BSA乳化油酸,再使其溶解于培養液中,而油酸必須與BSA分離后才能被細胞吸收,導致其有效濃度降低。這提示本研究的方法簡單方便,更具可行性。
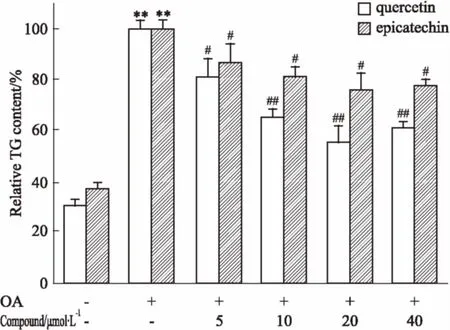
Fig 4 Inhibitory effects of flavonoids onlipid accumulation in HepG2 cells
**P<0.01vscontrol group;#P<0.05,##P<0.01vsOA model group
為了進一步確定該細胞模型適用于天然活性物質預防NAFLD的體外研究,本研究選用槲皮素和表兒茶素來評價黃酮類化合物對肝細胞脂肪變性的改善作用。槲皮素和表兒茶素是水果和蔬菜中常見的2種黃酮類化合物,研究表明其具有良好的降脂活性。本研究結果顯示,槲皮素和表兒茶素均能明顯抑制細胞內TG聚集,表明該細胞模型應用于植物活性物質預防NAFLD的體外研究是可行的[12]。此外,Zhang等[13]研究認為,槲皮素等黃酮類化合物抑制油酸負荷的肝細胞內TG聚集作用與其細胞內抗氧化活性密切相關。但在本研究模型中,HepG2細胞內并無明顯的氧化應激反應,推測槲皮素和表兒茶素等天然活性物質抑制該模型細胞內TG聚集作用可能是通過調節脂肪酸吸收、甘油三酯合成和分解相關基因的表達而實現[14-15]。
[1] Hassan K, Bhalla V, El Regal M E, et al. Nonalcoholic fatty liver disease: a comprehensive review of a growing epidemic[J].WorldJGastroenterol, 2014,20(34): 12082-101.
[2] Willebrords J, Pereira I V, Maes M, et al. Strategies, models and biomarkers in experimental non-alcoholic fatty liver disease research[J].ProgLipidRes, 2015,59:106-25.
[3] Yin J, Luo Y, Deng H, et al. Hugan Qingzhi medication ameliorates hepatic steatosis by activating AMPK and PPARα pathways in L02 cells and HepG2 cells[J].JEthnopharmacol, 2014,154(1): 229-39.
[4] Xie C, Chen Z, Zhang C, et al. Dihydromyricetin ameliorates oleic acid-induced lipid accumulation in L02 and HepG2 cells by inhibiting lipogenesis and oxidative stress[J].LifeSci, 2016,157: 131-9.
[5] Malhi H, Bronk S F, Werneburg N W, et al. Free fatty acids induce JNK-dependent hepatocyte lipoapoptosis[J].JBiolChem, 2006,281(17): 12093-101.
[6] Ucar F, Sezer S, Erdogan S, et al. The relationship between oxidative stress and nonalcoholic fatty liver disease: its effects on the development of nonalcoholic steatohepatitis[J].RedoxRep, 2013,18(4): 127-33.
[7] 劉 江, 厲有名, 陳韶華, 等. 一種實用的體外非酒精性脂肪肝細胞模型[J]. 浙江大學學報(醫學版), 2009,38(6): 626-9.
[7] Liu J, Li Y M, Chen S H, et al. Aninvitrohepatic steatosis cell model for study of non-alcoholic fatty liver disease[J].JZhejiangUniv(MedSci), 2009,38(6): 626-9.
[8] 殷錦錦, 唐外姣, 曾 璐, 等. 人肝細胞系 L-02 細胞單純肝脂肪變性細胞模型的建立與應用[J]. 南方醫科大學學報, 2014,34(6): 837-42.
[8] Yin J J, Tang W J, Zeng L, et al. Establishment of a L-02 cell model of hepatic steatosis[J].JSouthMedUniv, 2014,34(6): 837-42.
[9] Niklas J, Bonin A, Mangin S, et al. Central energy metabolism remains robust in acute steatotic hepatocytes challenged by a high free fatty acid load[J].BMBRep, 2012,45(7): 396-401.
[10] Cui W, Chen S L, Hu K Q. Quantification and mechanisms of oleic acid-induced steatosis in HepG2 cells[J].AmJTranslRes, 2010,2(1): 95-104.
[11] Liu Y, Wang D, Zhang D, et al. Inhibitory effect of blueberry polyphenolic compounds on oleic acid-induced hepatic steatosisinvitro[J].JAgricFoodChem, 2011,59(22): 12254-63.
[12] 王俊杰, 龍 婷, 曹 欣, 等. 荷葉黃酮對油酸孵育的 HepG2 細胞甘油三酯代謝的影響[J]. 中國藥理學通報, 2010,26(12): 1626-30.
[12] Wang J J, Long T, Cao X, et al. The influence of flavonoids extracted from lotus on the triglyceride metabolism of HepG2 cells incubated in oleic acid[J].ChinPharmacolBull, 2010,26(12): 1626-30.
[13] Zhang D, Xie L, Jia G, et al. Comparative study on antioxidant capacity of flavonoids and their inhibitory effects on oleic acid-induced hepatic steatosisinvitro[J].EurJMedChem, 2011,46(9): 4548-58.
[14] 徐 軍, 王國恩, 章時杰, 等. 1,3,7,9-四甲基尿酸激活SirT3/AMPK/ACC信號通路減少高脂飲食小鼠肝臟脂肪化[J]. 中國藥理學通報, 2014,30(6): 791-5.
[14] Xu J, Wang G E, Zhang S J, et al. Theacrine ameliorates high fat diet induced hepatic steatosis in mice via SirT3/AMPK/ACC pathway[J].ChinPharmacolBull, 2014,30(6): 791-5.
[15] 耿雅娜, 于 濱, 孔維佳, 等. 天麻素通過激活AMPK通路減少油酸誘導的HL-7702細胞脂肪蓄積[J]. 中國藥理學通報, 2015,31(1): 39-44.
[15] Geng Y N, Yu B, Kong W J, et al. Gastrodin ameliorates oleic acid-induced fat accumulation through activation of AMPK pathway in HL-7702 cells[J].ChinPharmacolBull, 2015,31(1): 39-44.
Establishmentandapplicationofhepatocytesteatosismodelsinducedbyoleicacid
DONG Li-hong, ZHANG Rui-fen, HUANG Fei, WEI Zhen-cheng, ZHANG Ming-wei
(SericulturalandAgri-FoodResearchInstitute,GuangdongAcademyofAgriculturalSciences/KeyLabofFunctionalFood,MinistryofAgriculture/GuangdongKeyLabofAgriculturalProductsProcessing,Guangzhou510610,China)
AimTo establish aninvitrocell model for investigating hepatic steatosis in the primary stage of non-alcoholic fatty liver disease (NAFLD).MethodsCultured HepG2 cells were divided into different group exposed to different concentrations of oleic acid for 24 h respectively. Lipid droplets in the cells were observed with Oil Red O staining and Nile red staining. The contents of intracellular TG, MDA and SOD and the levels of ALT and AST in the cell supernatant were evaluated to verify the cell model, and the effect of flavonoids on the TG accumulation was observed in oleic acid-induced HepG2.ResultsA large number of lipid droplets were found in the model group with 0.4 mmol·L-1oleic acid, which showed markedly increased level of triglyceride without significant changes of intracellular MDA content and SOD activity and the levels of ALT and AST in the cell supernatant, compared with the control group. Intervention with different doses of quercetin and epicatechin significantly decreased the TG content in the cell model.ConclusionHepatic steatosis HepG2 cell model can be established by 0.4 mmol·L-1oleic acid for the studies on lipid-lowering natural drugs in the prevention of NAFLD.
oleic acid; HepG2 cells; steatosis; triglyceride; cell model; flavonoids
A
1001-1978(2017)11-1622-05
R322.47;R329.24;R344.3;R575.5;R977.6
時間:2017-10-10 10:05 網絡出版地址:http://kns.cnki.net/kcms/detail/34.1086.R.20171010.1005.056.html
10.3969/j.issn.1001-1978.2017.11.028
2017-08-06,
2017-09-07
國家自然科學基金資助項目(No 31571828);國家自然科學基金-廣東聯合基金資助項目(No U1301211)
董麗紅(1991-),女,碩士,研究方向:功能食品,E-mail:dolify@163.com; 張名位(1966-),男,博士,研究員,博士生導師,研究方向:功能食品,通訊作者,E-mail:mwzhh@vip.tom.com
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
網絡安全與數據管理(2022年1期)2022-08-29 03:15:20
導航定位學報(2022年4期)2022-08-15 08:27:00
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
新世紀智能(數學備考)(2021年9期)2021-11-24 01:14:36
成都醫學院學報(2021年2期)2021-07-19 08:35:14
新世紀智能(數學備考)(2020年9期)2021-01-04 00:25:14
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
光學精密工程(2016年6期)2016-11-07 09:07:19