牛源多殺性巴氏桿菌莢膜血清型及毒力基因檢測(cè)
2017-11-09 11:07:06董文龍張喜慶田佳琪馬紅霞高云航
中國(guó)獸醫(yī)雜志 2017年9期
王 羽 , 董文龍 , 王 巍 , 張喜慶 , 田佳琪 , 馬紅霞 , 高云航
(吉林農(nóng)業(yè)大學(xué)動(dòng)物科學(xué)技術(shù)學(xué)院 , 吉林 長(zhǎng)春 130118)
牛源多殺性巴氏桿菌莢膜血清型及毒力基因檢測(cè)
王 羽 , 董文龍 , 王 巍 , 張喜慶 , 田佳琪 , 馬紅霞 , 高云航
(吉林農(nóng)業(yè)大學(xué)動(dòng)物科學(xué)技術(shù)學(xué)院 , 吉林 長(zhǎng)春 130118)
為了確定12株牛源多殺性巴氏桿菌的血清型及毒力基因的攜帶情況,本研究采用PCR技術(shù)對(duì)12株牛源多殺性巴氏桿菌(pasteurella maltocida,Pm)進(jìn)行莢膜血清型鑒定和6種毒力基因(tbpA、hgbA、hgbB、ptfA、pfhA、toxA)的檢測(cè),并對(duì)其序列進(jìn)行比對(duì)分析。結(jié)果顯示:12株多殺性巴氏桿菌均為莢膜血清A型;100%的莢膜血清A型Pm攜帶hgbA毒力基因和pfhA毒力基因;41.67%的莢膜血清A型Pm攜帶nanH毒力基因;58.33%的莢膜血清A型Pm攜帶ptfA毒力基因;而對(duì)于tbpA與toxA兩種毒力基因在12株牛源莢膜血清A型Pm中均未檢測(cè)到。
多殺性巴氏桿菌 ; 莢膜血清型 ; 毒力基因
多殺性巴氏桿菌是重要的畜禽致病菌,可引起多種動(dòng)物出現(xiàn)巴氏桿菌病。根據(jù)菌株間抗原差異,多殺性巴氏桿菌可分為多個(gè)血清型。根據(jù)被動(dòng)血凝試驗(yàn)對(duì)莢膜抗原(K抗原)可分為A、B、D、E和F 5個(gè)血清型,用凝集反應(yīng)對(duì)菌體抗原(O抗原)分類(lèi),可分為12個(gè)血清型,用瓊脂擴(kuò)散試驗(yàn)對(duì)熱浸出菌體抗原分類(lèi)可分為16個(gè)血清型[1]。多殺性巴氏桿菌血清型與致病性存在一定的相關(guān)性,如A型可以引起禽霍亂、豬肺疫;B型可以引起牛和豬等動(dòng)物的敗血癥;D和E型可以引起豬、牛、兔、羊等動(dòng)物的肺炎和敗血癥;F型主要發(fā)生于火雞,其致病作用目前尚不清楚[2]。同時(shí)相關(guān)試驗(yàn)已廣泛證實(shí),Pm的莢膜在抗吞噬作用中也起著重要作用,且菌體抗吞噬作用的敏感性與莢膜的存在及厚度有關(guān)[3]。各血清型多殺性巴氏桿菌的流行性存在較大差異,具有明顯的地方特性,故相同血清型不同地方分離的菌株毒力可能不同[1,4]。
Faham Khamesipour等從333頭健康及患病牛體內(nèi)分離了30株多殺性巴氏桿菌,并進(jìn)行了莢膜基因及23個(gè)毒力相關(guān)基因的檢測(cè),23株為莢膜A型多殺性巴氏桿菌,56.5%菌株攜帶pfhA基因,78.3%菌株攜帶tbpA和nanH基因,ptfA及nanH基因攜帶率為82.6%,87.0%菌株攜帶hgbA基因,hgbB基因攜帶率高達(dá)100%,但只有13.0%菌株攜帶toxA基因[5]。本研究對(duì)從吉林省不同地區(qū)分離得到的牛源莢膜血清A型多殺性巴氏桿菌的毒力基因進(jìn)行檢測(cè),為客觀評(píng)估牛巴氏桿菌病的危害提供依據(jù)。
1 材料與方法
1.1 菌種 經(jīng)分離并鑒定的12株牛源多殺性巴氏桿菌保存于吉林農(nóng)業(yè)大學(xué)動(dòng)物科學(xué)技術(shù)學(xué)院預(yù)防獸醫(yī)學(xué)研究室。
1.2 主要試劑 胰蛋白胨大豆肉湯培養(yǎng)基(TSB)、胰蛋白胨大豆瓊脂培養(yǎng)基(TSA),購(gòu)自青島高科園海博生物技術(shù)有限公司;胎牛血清,購(gòu)自北京元亨圣馬生物技術(shù)研究所;ExTaq酶、dNTPs、DNA Marker 2 000、6×ExBuffer等,購(gòu)自TaKaRa公司;膠回收試劑盒等,購(gòu)自上海生工生物工程技術(shù)服務(wù)有限公司。
1.3 致病性試驗(yàn) 將分離純化的菌株接種到TSA培養(yǎng)基(含50 mL//L 胎牛血清),37 ℃培養(yǎng)24 h。并進(jìn)行擴(kuò)菌培養(yǎng),并將菌液按10倍系數(shù)倍比稀釋。隨機(jī)將小鼠分為13組,每組5只。試驗(yàn)組每只注射0.3 mL 多殺性巴氏桿菌,對(duì)照組每只小鼠接種0.3 mL 生理鹽水。觀察并記錄各組小鼠的發(fā)病及死亡情況,以寇氏改良法計(jì)算半數(shù)致死量(LD50) , 同時(shí)采集死亡小鼠的肝臟、心臟進(jìn)行細(xì)菌的分離鑒定。
1.4 PCR鑒定 參考Laxmi Narayan Sarangi[5]等報(bào)道的多殺性巴氏桿菌莢膜血清型基因hyaD-hyaC、bcbD、dcbF、ecbJ、fcbD(如表1)和毒力基因tbpA、hgbA、hgbB、ptfA、pfhA、toxA設(shè)計(jì)引物(如表2),并送往(上海)生工生物工程技術(shù)服務(wù)有限公司合成。

表1 莢膜血清型基因引物序列及預(yù)計(jì)大小

表2 毒力基因引物序列及預(yù)計(jì)大小
1.5 DNA提取及毒力基因擴(kuò)增 挑取單個(gè)菌落接種于TSB液體培養(yǎng)基中(含50 mL//L 胎牛血清),置于37 ℃水平搖床(170 r/min)增菌培養(yǎng)。取1.6 mL菌液12 000 r/min離心,棄上清,運(yùn)用酚-氯仿法提取分離菌株DNA。
以提取的DNA為模板,對(duì)莢膜血清型和毒力基因進(jìn)行擴(kuò)增。PCR擴(kuò)增條件為94 ℃ 5 min;94 ℃ 45 s;退火(退火溫度如表1、2)45 s;72 ℃ 45 s;30個(gè)循環(huán);72 ℃延伸10min;4 ℃保存。隨后用瓊脂糖凝膠電泳檢測(cè)PCR產(chǎn)物。對(duì)PCR產(chǎn)物進(jìn)行膠回收純化,將純化的PCR產(chǎn)物和pMD-18T連接過(guò)夜,將連接產(chǎn)物轉(zhuǎn)化到E.coli.DH5α 感受態(tài)細(xì)胞,按照試劑盒提取質(zhì)粒進(jìn)行驗(yàn)證,并送至上海生工生物工程技術(shù)服務(wù)有限公司測(cè)序。
2 結(jié)果
2.1 致病性試驗(yàn)結(jié)果 12株多殺性巴氏桿菌的LD50分別為6.29×105CFU/mL,2.1×103CFU/mL,3.5×105CFU/mL,7.1×108CFU/mL,4.32×105CFU/mL,1.23×104CFU/mL,2.9×105CFU/mL,6.78×105CFU/mL,1.42×104CFU/mL,9.3×105CFU/mL,1.9×106CFU/mL,4.5×105CFU/mL 。剖檢病死小鼠,觀察病變,并進(jìn)行細(xì)菌分離及分子生物學(xué)鑒定,確定為多殺性巴氏桿菌。
2.2 莢膜血清型PCR擴(kuò)增結(jié)果 以基因組DNA為模板,采用PCR技術(shù)對(duì)12株P(guān)m進(jìn)行莢膜型基因進(jìn)行擴(kuò)增,均擴(kuò)增出1 000 bp大小左右的片段。擴(kuò)增序列與GenBank上已公布的莢膜血清型特異性基因hyaD-hyaC多殺性巴氏桿菌(AY225345.1)序列同源性達(dá)到99%,據(jù)此可鑒定該菌為A型多殺性巴氏桿菌。hyaD-hyaC基因擴(kuò)增結(jié)果如圖1。

圖1 hyaD-hyaC 基因擴(kuò)增結(jié)果M:DL-2 000 Marker; Pm-1—Pm-2:多殺性巴氏桿菌
2.3 毒力基因擴(kuò)增結(jié)果
2.3.1hgbA基因PCR擴(kuò)增及序列檢測(cè)結(jié)果 10株菌擴(kuò)增出約700 bp的片段,與預(yù)期片段大小相符,根據(jù)BLAST對(duì)比結(jié)果可知,菌株P(guān)m-1、Pm-3、Pm-5、Pm-6、Pm-7、Pm-9、Pm-11擴(kuò)增序列與GenBank上已公布的Pm毒力基因hgbA(登錄號(hào)AF237923.1)同源性為100%,Pm-2、Pm-4、Pm-8、Pm-10、Pm-12擴(kuò)增序列與登錄號(hào)AF237923.1同源性為99%。hgbA基因擴(kuò)增結(jié)果如圖2。

圖2 hgbA 基因擴(kuò)增結(jié)果M:DL-2 000 Marker; Pm-1—Pm-2:多殺性巴氏桿菌
2.3.2nanH基因PCR擴(kuò)增及序列檢測(cè)結(jié)果 菌株P(guān)m-1、Pm-3、Pm-7、Pm-11、 Pm-12株P(guān)m均擴(kuò)增出900 bp左右的片段,該基因片段測(cè)序后使用BLAST對(duì)其堿基序列進(jìn)行分析,其結(jié)果顯示,所擴(kuò)增的nanH相應(yīng)序列與GenBank上已公布的Pm毒力基因nanH(登錄號(hào)AF274869.1)序列同源性為95%。nanH基因擴(kuò)增結(jié)果如圖3。
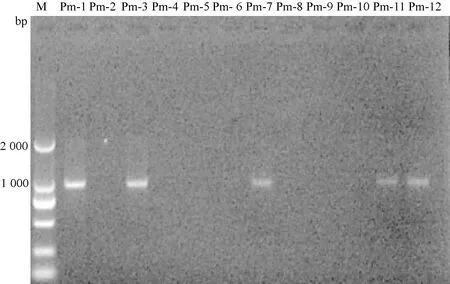
圖3 nanH基因擴(kuò)增結(jié)果M:DL-2 000 Marker; Pm-1—Pm-2:多殺性巴氏桿菌
2.3.3pfhA基因PCR擴(kuò)增及序列檢測(cè)結(jié)果 12株P(guān)m均擴(kuò)增出800 bp左右的片段,根據(jù)BLAST對(duì)比結(jié)果可知,菌株P(guān)m-1、Pm-2、Pm-4、Pm-5、Pm-10、Pm-12與GenBank上已公布的Pm毒力基因pfhA(登錄號(hào)AF237929.1)同源性為100%,Pm-3、Pm-6、Pm-7、Pm-8、Pm-9、Pm-11與登錄號(hào)AF237929.1同源性為99%。12株P(guān)m的pfhA基因擴(kuò)增結(jié)果如圖4。
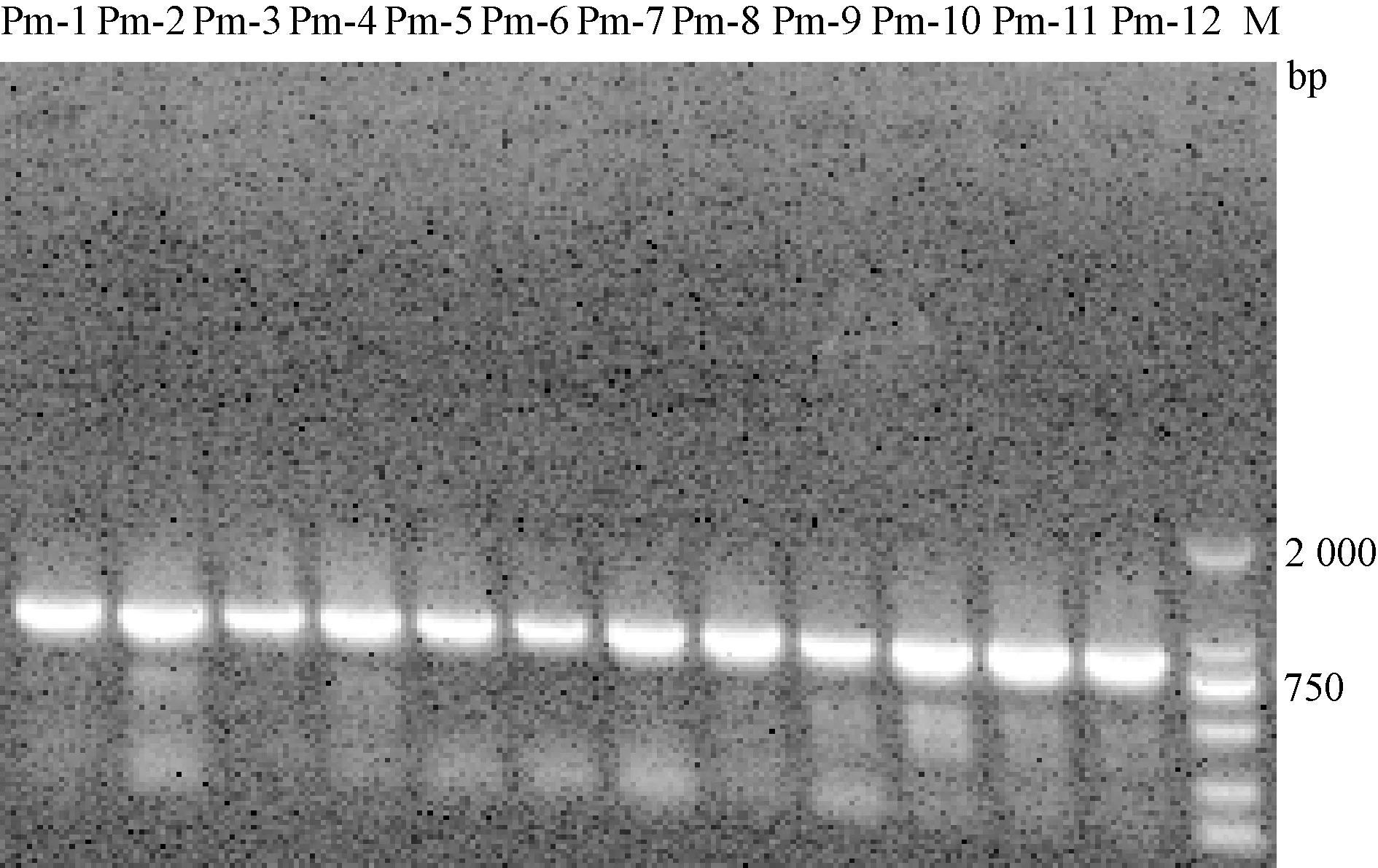
圖4 pfhA基因擴(kuò)增結(jié)果M:DL-2 000 Marker;Pm-1—Pm-2:多殺性巴氏桿菌
2.3.4ptfA基因PCR擴(kuò)增及序列檢測(cè)結(jié)果 菌株P(guān)m-1、Pm-2、Pm-4、Pm-5、Pm-9、Pm-10、Pm-11均擴(kuò)增出約400 bp左右大小的片段,進(jìn)行BLAST分析,其結(jié)果顯示7株菌擴(kuò)增序列與GenBank上已公布的Pm毒力基因ptfA(登錄號(hào)KP726888.1)同源性均為100%。ptfA基因擴(kuò)增結(jié)果如圖5。

圖5 ptfA基因擴(kuò)增結(jié)果M:DL-2 000 Marker;Pm-1—Pm-2:多殺性巴氏桿菌
2.3.5tbpA基因及toxA基因PCR擴(kuò)增結(jié)果 以基因組DNA為模板,采用PCR技術(shù)對(duì)tbpA基因與toxA基因進(jìn)行擴(kuò)增,均未檢測(cè)到。
3 討論
多殺性巴氏桿菌是一種有莢膜的革蘭陰性桿菌,由于莢膜血清型決定病原菌的免疫原性,各血清型之間交叉免疫保護(hù)性較低,對(duì)該致病菌進(jìn)行莢膜分型就顯得尤為重要[6]。1984 年Carter G R 等[7]提出多殺性巴氏桿菌血清型的標(biāo)準(zhǔn)定名,分為A、B、D、E 和F 5 個(gè)莢膜血清型。近年來(lái)在牛群中流行的主要是以引起肺部感染為主的莢膜血清A型巴氏桿菌[8]。本研究對(duì)在不同地區(qū)分離到12株多殺性巴氏桿菌進(jìn)行血清分型,12株菌均為莢膜A型。
自從1959年世界衛(wèi)生組織(WHO)將巴氏桿菌病定為一類(lèi)重要?jiǎng)游飩魅静。琍m就一直受到各國(guó)學(xué)者的關(guān)注[9]。在本研究中對(duì)6種毒力基因的檢測(cè)結(jié)果顯示,12株牛源莢膜A型多殺性巴氏桿菌均擴(kuò)增出pfhA基因和基因hgbA,菌株P(guān)m-1、Pm-2、Pm-4、Pm-5、Pm-9、Pm-10、Pm-11含有基因ptfA,菌株P(guān)m-1、Pm-3、Pm-7、Pm-11和Pm-12含有nanH基因。12株菌株均未擴(kuò)增出toxA和tbpA基因。菌株P(guān)m-1、Pm-2、Pm-4、Pm-9有相同的毒力基因,動(dòng)物試驗(yàn)結(jié)果表明不同菌株間其毒力存在一定差異,菌株2毒力最強(qiáng),菌株4毒力最弱。通過(guò)本試驗(yàn)可得出兩種推斷:(1)菌株2和4毒力差異可能與基因表達(dá)程度以及調(diào)控不同而造成;(2)可能還有其他基因參與。毒力因子之間的相互作用還有待于進(jìn)一步研究。
[1] 李一經(jīng).獸醫(yī)微生物學(xué)[M].北京:高等教育出版社,2011.
[2] 陸承平.獸醫(yī)微生物學(xué)[M].3版.北京:中國(guó)農(nóng)業(yè)出版社,2001.
[3] Truscott W M, Hirsh D C. Demonstration of an outer membrane protein with antiphagocytic activity fromPasteurellamultocidaof avian origin[J]. Infect Immun, 1988, 56: 1 538-1 544.
[4] Ahmad T A, Rammah S S, Sheweita S A,etal. Development of immunization trials againstPasteurellamultocida[J]. Vaccine, 2014, 32(8):909-917.
[5] Khamesipour F, Momtaz H, Mamoreh M A. Occurrence of virulence factors and antimicrobial resistance inPasteurellamultocidastrains isolated from slaughter cattle in Iran[J]. Front Microbiol, 2014, 5:536.
[6] Sarangi L N, Priyadarshini A, Kumar S,etal. Virulence Genotyping ofPasteurellamultocidaIsolated from Multiple Hosts from India[J]. Scientific World Journal, 2014, 2014:814 109-814 109.
[7] 馬文戈,于力. 牛源莢膜血清A型多殺性巴氏桿菌的分離鑒定[J]. 中國(guó)預(yù)防獸醫(yī)學(xué)報(bào),2008,30(10):747-750+754.
[8] Carter G R,Genus I.Pasteurella.In:Bergey’s manual of systematic bacteriology[M].Baltimore:Williams and Wilkins,1984:554-557.
[9] Confer A W, Nutt S H, Dabo S M,etal. Antibody responses of cattle to outer membrane proteins ofPasteurellamultocidaA:3.[J]. Am J VET RES, 1996, 57(10):1 453-1 457.
[10] Arashima Y, Kumasaka K.Pasteurellosisas zoonosis[J].Intern Med,2005,44:692-693.
CapsularSerotypeandVirulenceGeneDetectiononBovineSourcePasteurellamultocida
WANG Yu , DONG Wen-long , WANG Wei , ZHANG Xi-qing , TIAN Jia-qi , MA Hong-xia , GAO Yun-hang
(College of Animal Science and Technology,Jilin Agricultural University, Changchun 130118,China)
To determine the serotype and virulence genes of 12 strains ofPasteurellamultocida. We use PCR technology for detection and sequence analysis of 12 Bovine sourcePasteurellamultocida(Pm)of capsular serotype and 6 virulence genes (ptfA,pfhA,hgbA,hgbB,tbpA,toxA).The results showed that all 12Pmstrains are capsular serotype A, 100% of the bovine source capsular serotype APmstrains carry virulence genespfhAand virulence genes hgbA;41.67% of bovine source capsular serotype APmstrains carry virulence genes nanH; 58.33% of bovine source capsular serotype APmstrains carry virulence genes ptfA; tbpA and toxA was not detected in 12 bovine source capsular serotype APmstrains.
Pasteurella multocida; capsular serotype;virulence genes
GAO Yun-hang
S852.65+3 文獻(xiàn)杯志碼:A
0529-6005(2017)09-0016-04
2017-03-12
王羽(1993-),女,碩士生,研究方向?yàn)閯?dòng)物疫病防治,E-mail:1846020084@qq.com
高云航,E-mail:gaoyunhang@163.com
