陜西子午嶺國家級自然保護區鞘翅目昆蟲群落的邊緣效應
2017-11-10 03:11:57徐世才王麗娟
生態科學 2017年5期
徐世才, 王麗娟
延安大學生命科學學院, 陜西延安716000
陜西子午嶺國家級自然保護區鞘翅目昆蟲群落的邊緣效應
徐世才*, 王麗娟
延安大學生命科學學院, 陜西延安716000
選取陜西子午嶺國家級自然保護區闊葉林—針葉林、農田—灌叢和草地—灌叢3種邊緣地帶為研究樣地, 在科級水平上探討邊緣效應對鞘翅目昆蟲群落多樣性的影響。共采集鞘翅目昆蟲4935只, 隸屬于28科, 其中步甲科、葉甲科和瓢蟲科的個體數量分別占個體總數的44.19%、21.95%和13.66%, 構成該地區鞘翅目昆蟲的優勢類群。3種邊緣類型中鞘翅目昆蟲的科多樣性、豐富度和均勻度均沿邊緣向相鄰生境內部降低,EH′值均大于 1, 呈現邊緣正效應。經多元回歸分析表明, 植被密度影響闊葉林—針葉林邊緣鞘翅目昆蟲群落的多樣性和豐富度, 植被密度和蓋度影響整個群落鞘翅目昆蟲的個體數量分布。以上結論可為陜西子午嶺自然保護區鞘翅目昆蟲生物多樣性的保護和合理利用提供科學依據。
鞘翅目; 群落多樣性; 邊緣效應
1 前言
頻繁的人類活動和自然災害的發生, 加速了生境的破碎化與生物棲息地的喪失。片段化的森林代替了原有大面積的自然生境, 形成不同的斑塊與邊緣。邊緣效應的存在使得森林內部到外部的生態環境產生梯度變化, 包括棲息地、食物、繁殖場所等[1]。生物由于自身及外界環境等因素的影響, 對邊緣產生不同的反應, 研究物種對邊緣的選擇趨勢或行為反應對邊緣效應的理解與生物多樣性的保護具有特定的價值。鞘翅目(Coleoptera)昆蟲是昆蟲綱中最大的一個目, 占全球已知昆蟲總數的25%[2]。鞘翅目昆蟲食性多樣, 在整個生態系統中扮演著重要的角色[3],且在維持生物多樣性等方面具有重要作用, 其中部分類群還常被選作指示性昆蟲來反映環境的變化情況[4–5], 對其的研究也逐漸受到國內外學者的重視。
由于人為的破壞, 如亂砍濫伐和過度開墾等,子午嶺森林由相對完整逐漸走向破碎化, 形成了片段化的景觀和天然次生林[6]。隨著人們逐漸認識到子午嶺在維持黃土高原地區生態穩定方面的重要性,2006年成立了陜西子午嶺國家級自然保護區, 促進了該保護區動、植物多樣性的保護[7], 生物多樣性也得到逐步恢復, 但生態恢復過程中生物多樣性的恢復狀況缺乏相應的監測與評價。本文選取子午嶺闊葉林—針葉林、農田—灌叢和草地—灌叢3種邊緣類型, 以鞘翅目昆蟲為研究對象, 通過比較不同邊緣鞘翅目昆蟲在科級水平上的群落多樣性及其邊緣效應, 分析影響鞘翅目昆蟲分布的植被特征, 研究子午嶺在生態恢復過程中生境片段化對鞘翅目昆蟲多樣性的影響, 以期為子午嶺鞘翅目昆蟲生物多樣性的保護及合理利用, 子午嶺保護區植被恢復與重建提供參考。
2 研究地區與研究方法
2.1 研究區概況
子午嶺國家級自然保護區位于陜西省富縣境內,南與渭北大平原相接, 西靠子午嶺主脊, 北臨毛烏素沙漠邊緣, 地理坐標在東經 108°29′—108°41′, 北緯 35°45′—36°01′之間, 總面積 40621 hm2。保護區氣候屬暖溫帶半濕潤氣候, 年平均氣溫7.5—8.9 ℃,晝夜溫差大, 年降水量520—610 mm, 南多北少。植被類型是從森林草原—半干早草原的過渡地帶, 地帶性植被為華北暖溫帶落葉闊葉林地帶的北部落葉闊葉林。依據《中國植被》分類原則, 保護區的植被可分為森林、灌叢和草地3種植被類型, 共20個群系, 主要樹種有遼東櫟(Quercus wutaishanica)、油松(Pinus tabulaeformis)、山楊(Populus davidiana)、小葉楊(Populus simonii)、白樺(Betula platyphylla)和側柏(Platycladus orientalis)等。此外還有大果榆(Ulmus macrocarpa)、野杏(Armeniaca vulgaris)、小葉樸(Celtis bungeana)、茶條槭(Acer ginnala)、白蠟樹(Fraxinus chinensis)、北京丁香(Syringa pekinensis)等華北樹種,常以小片狀分布或與主要樹種伴生或散生。林下及散生灌木主要有狼牙刺(Sophora viciifolia)、沙棘(Hippophae rhamnoides)、虎榛子(Ostryopsis davidiana)、文冠果(Xanthoceras sorbifolium)、連翹(Forsythia suspense)、胡枝子(Lespedeza bicolor)、土莊繡線菊(Spiraea pubescens)等。草本植物主要有菊科(Compositae)、禾本科(Gramineae)、苔草(Carex)、莎草(Cyperus)等。此外, 保護區內還存有農田和苗圃, 主要種植玉米(Zea mays)和牡丹(Paeonia suffruticosa)。
2.2 調查方法
根據陜西子午嶺保護區的自然條件, 以保護區內槐樹莊保護站至榆林站為主線, 選取闊葉林—針葉林、農田—灌叢和草地—灌叢3種類型的邊緣地帶進行研究, 依據采捕的種類及生境的具體情況, 利用網捕法、直接搜捕法和誘捕法等采捕樣地內的鞘翅目昆蟲并制作標本, 依據相關文獻專著鑒定[8–11]并記錄鞘翅目昆蟲的種類和數量。采集工作在 2016年5—9月進行, 每月中旬采一次樣。
參考 Meiners和 Pickett[12]及 Heli?l?等[13]研究邊緣效應的方法, 以距離梯度形式研究邊緣及相鄰生境內部鞘翅目昆蟲群落的差異。設計樣點時, 在與邊緣垂直方向上設置5條樣帶, 樣帶間隔為10 m, 每條樣帶以邊緣為中心, 向相鄰生境內部延伸100 m, 以每隔25 m的距離設置樣點, 大小為2 m×5 m, 具體取樣方法見圖1(示兩條樣帶)。
為研究鞘翅目昆蟲分布與植被特征之間的關系,統一在調查中期(2016年7月)調查記錄各個樣帶(以樣點中央為中心, 直徑2 m范圍內)的植被種類、密度、蓋度和凋落物厚度等特征。
2.3 數據處理與分析
以鞘翅目昆蟲在科級水平上的豐富度、個體數量、多樣性、均勻度及優勢度為數據分析對象, 將每種邊緣類型的 45個研究樣點依據與邊緣的距離聚為3組: 生境1包括A、B和C(分別距邊緣100 m、75 m和50 m)3條樣地帶, 邊緣包括D、E和F(分別距邊緣25m、0m和25m)3條樣地帶, 生境2包括余下的G、H和I(分別距邊緣50 m、75 m和100 m)3條樣地帶。
科多樣性分析主要選用 4類指數, 即 Shannon-Wiener多樣性指數、Margalef豐富度指數、Pielou均勻度指數和Simpson優勢度指數, 計算公式如下[14–15]:
Shannon-Wiener多樣性指數(H′)H′=-∑PilnPi
Margalef豐富度指數(D)D=(S-1)/lnN
Pielou均勻度指數(J)J=H′/lnS
Simpson優勢度指數(C)C=∑(ni/N)2
式中Pi=ni/N,Pi為第i科占總個體數的比率,ni是第i科的個體數,N是個體總數,S表示群落科數。
邊緣效應強度(E)根據測度模型:E=mY/∑yi, 令由m個群落組成的交錯區的這一指標為Y,m個群落的這一指標為yi(i=1, 2, 3, …,m)。再以 Shannon-Wiener多樣性指數(H′)和Simpson優勢度指數(C)這兩個定量指標去擬合上述公式, 則可得出邊緣效應值EH'和EC:

其中H′為群落組成的交錯區鞘翅目昆蟲科的多樣性指數;Hi′為各個群落里鞘翅目昆蟲科的多樣性指數;C為群落組成的交錯區內鞘翅目昆蟲科的優勢度指數;Ci為各個群落里鞘翅目昆蟲科的優勢度指數[16]。
利用Excel 2016和SPASS 19.0對實驗數據進行分析處理, 通過多元線性逐步回歸分析檢驗鞘翅目昆蟲科的多樣性、豐富度、均勻度以及優勢度與植被特征之間的關系。
3 結果與分析
3.1 研究地鞘翅目昆蟲群落組成分析
調查時間內共捕獲鞘翅目昆蟲4935號, 分屬于24 科(表1)。其中步甲科、葉甲科和瓢蟲科的個體數量最多, 分別占個體總數的 44.19%、21.95%和13.66%, 均大于 10%, 為該地區的優勢類群; 叩甲科、天牛科、金龜科、擬步甲科和芫菁科的個體數量在 1%—10%之間, 為該地區的常見類群; 虎甲、鍬甲、象甲、麗金龜、花金龜、蜣螂、隱翅蟲、小蠹蟲、豆象、吉丁甲、犀金龜、皮金龜、蜉金龜、葬甲、郭公蟲和螢科等 16科個體數量少于1%, 為該地區較不常見類群。
3.2 不同邊緣距離鞘翅目昆蟲的分布
在科級水平上, 通過比較闊葉林—針葉林邊緣、農田—灌叢邊緣和草地—灌叢邊緣不同邊緣距離鞘翅目昆蟲的個體數量與豐富度(圖 2, 3)發現, 3種邊緣類型中鞘翅目昆蟲的個體數量均沿邊緣分別向相鄰生境內部降低, 且整體上鞘翅目昆蟲的個體數量以闊葉林—針葉林邊緣>草地—灌叢邊緣>農田—灌叢邊緣, 闊葉林>針葉林>灌叢>草地>農田。闊葉林—針葉林邊緣鞘翅目昆蟲的豐富度沿邊緣向闊葉林內部先升高后降低, 沿邊緣向針葉林內部降低; 農田—灌叢邊緣鞘翅目昆蟲的豐富度變化趨勢與闊葉林—針葉林邊緣大致相同, 沿邊緣向農田內部先升高后降低, 向灌叢內部降低;草地—灌叢邊緣鞘翅目昆蟲的豐富度沿邊緣分別向草地和灌叢內部降低, 且整體上以灌叢內部的豐富度高于草地。
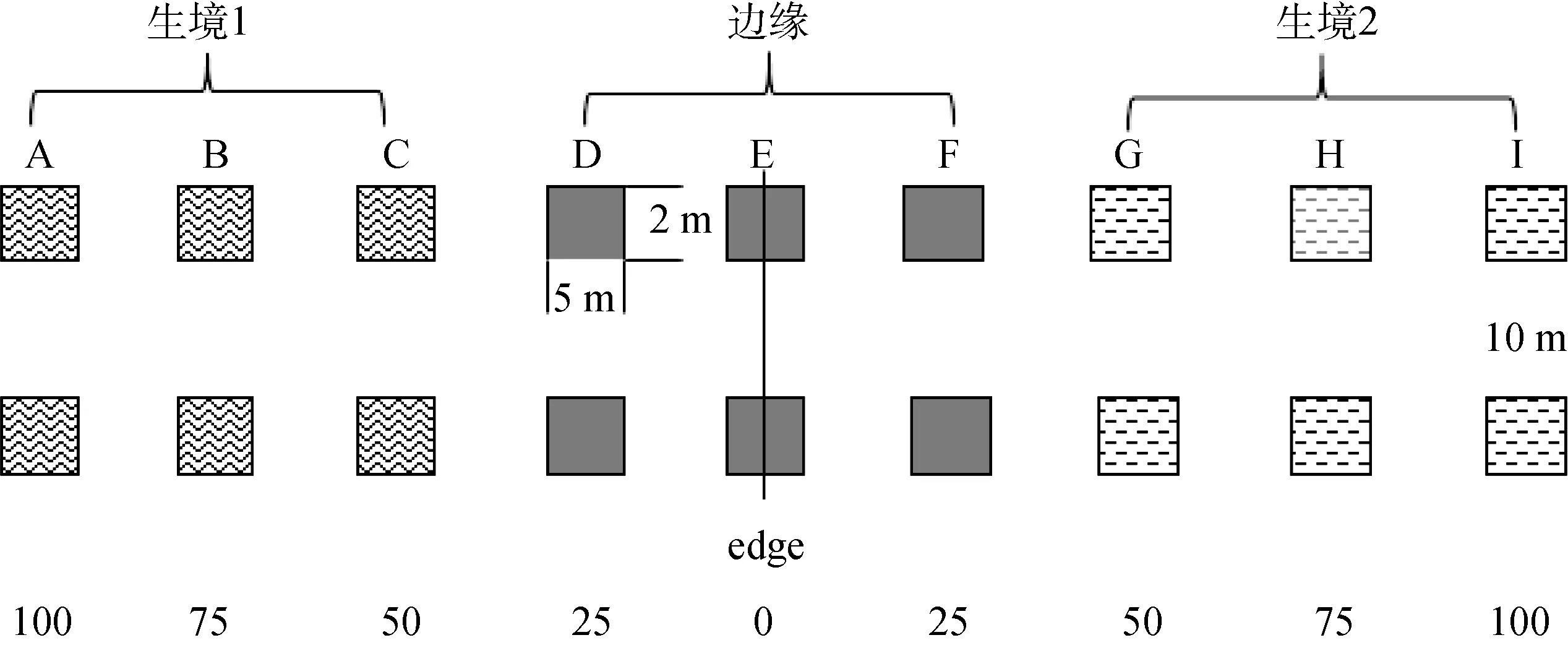
圖1 樣地設置Fig.1 The sample setting
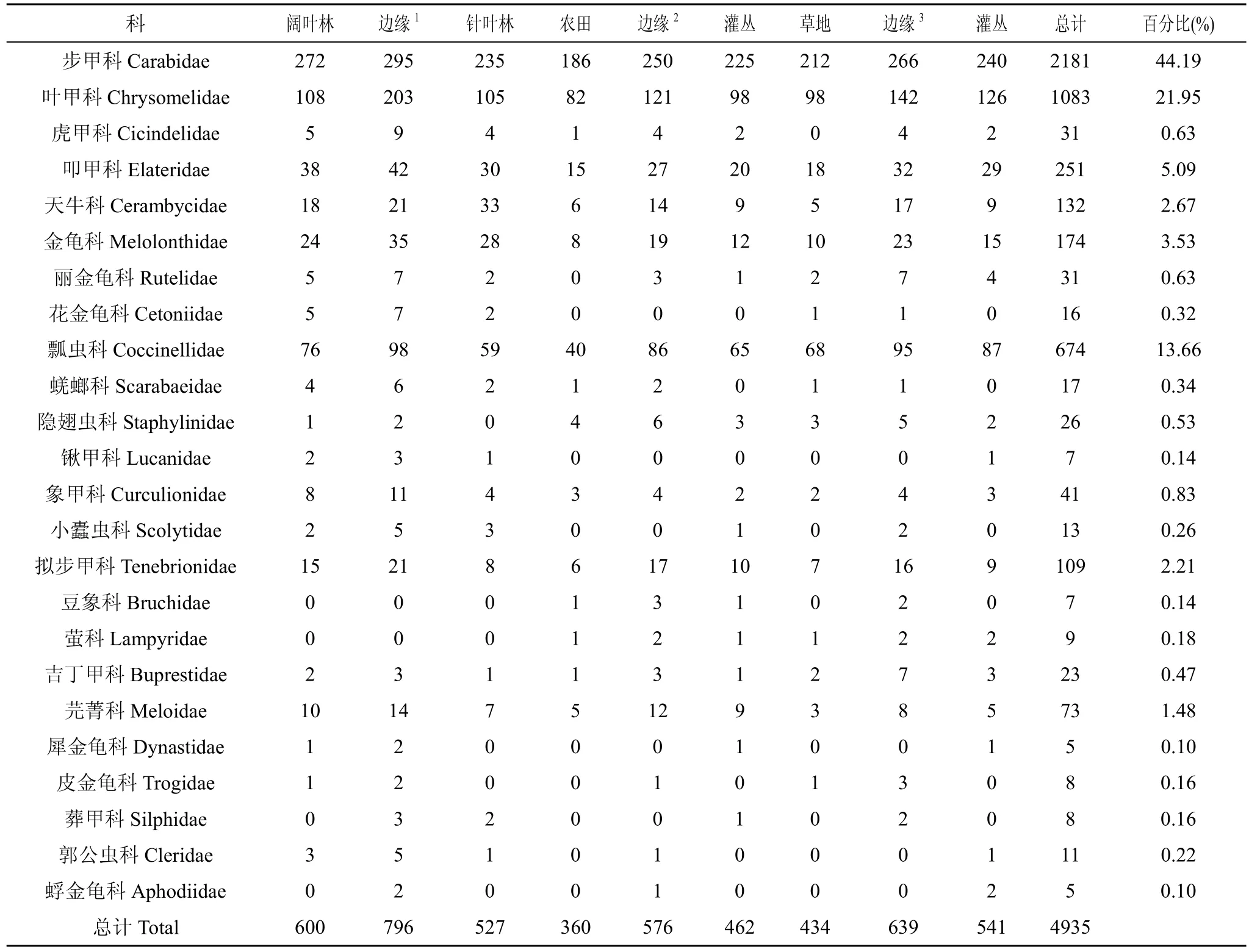
表1 各邊緣類型鞘翅目昆蟲的組成與數量分布Tab.1 Composition and quantity of Coleoptera insects in different edges
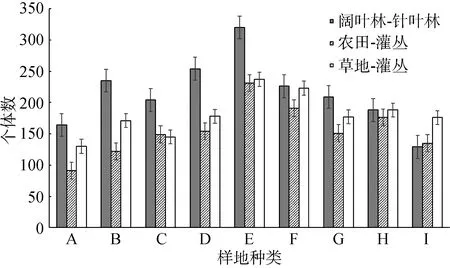
圖2 不同邊緣距離鞘翅目昆蟲個體數量Fig.2 Individuals of Coleoptera insects in different transect
3.3 不同邊緣類型鞘翅目昆蟲邊緣效應分析
本調查涉及的3種邊緣及其相鄰生境中鞘翅目昆蟲群落多樣性指數見表2。邊緣地帶鞘翅目昆蟲的Shannon-Wiener多樣性、豐富度和均勻度均高于其相鄰生境, 總體上各多樣性指數的排列順序為:闊葉林—針葉林邊緣>草地—灌叢邊緣>農田—灌叢邊緣。

圖3 不同邊緣距離鞘翅目昆蟲科豐富度Fig.3 Family richness of Coleoptera insects in different transect
在闊葉林—針葉林邊緣中, 鞘翅目昆蟲的Shannon-Wiener多樣性與豐富度變化趨勢相同, 總體上以闊葉林—針葉林邊緣>闊葉林>針葉林, 但均勻度與優勢度在闊葉林與針葉林中基本相等。農田—灌叢邊緣鞘翅目昆蟲的Shannon-Wiener多樣性與豐富度以邊緣處最高, 且灌叢內部大于農田, 均勻度農田內部稍大于灌叢, 優勢度的變化趨勢與Shannon-Wiener多樣性和豐富度相反。草地—灌叢邊緣, 鞘翅目昆蟲群落的Shannon-Wiener多樣性、豐富度和均勻度的變化趨勢為: 草地—灌叢邊緣>灌叢>草地, 與優勢度的變化趨勢相反。各生境鞘翅目昆蟲群落的 Shannon-Wiener多樣性排列順序為:闊葉林>針葉林>灌叢>農田>草地, 林地因其擁有豐富的植被類型與較高的生產力, 同時受人為干擾少, 所以鞘翅目昆蟲群落 Shannon-Wiener多樣性與豐富度高; 農田由于其特殊的生境, 種植類型單一, 且受到的人為干擾較大, 具有較低的群落豐富度, 但由于其內分布的鞘翅目昆蟲種類雖少但個體數量較多, 且分布均勻, 因此其多樣性指數H′并不低。
邊緣效應強度可對邊緣效應的強弱進行定量測量, 據王伯蓀等人所組建的測度模型[16]的生態學意義, 較大的EH′值表示具有較高的邊緣效應強度, 當EH′>1時, 可認為呈正的邊緣效應, 當EH′<1時則呈負的邊緣效應。對于EC值, 則較高的邊緣效應強度具有較低的EC值, 故在測度時EC值可變換為倒數。由邊緣效應強度分析結果(表3)得出, 闊葉林—針葉林邊緣、農田—灌叢邊緣和草地—灌叢邊緣的EH′值均大于1, 表明這3種邊緣類型均呈正的邊緣效應作用。闊葉林—針葉林邊緣地帶由于兩相鄰生境植被類型豐富, 各環境因子的分配差異較小, 鞘翅目昆蟲的種類和數量分布均勻, 因此具有較高的群落多樣性, 導致其EH′值低于農田—灌叢邊緣和草地—灌叢邊緣。用EC值來測定也能得到相同的結果, 即 3種邊緣類型均呈正的邊緣效應作用。
3.4 鞘翅目昆蟲群落多樣性與植被特征的相關性
對鞘翅目昆蟲的科多樣性、豐富度、均勻度、優勢度和個體數量與植物群落多樣性、植被密度、植被蓋度和凋落物厚度這4種植被特征進行多元線性回歸檢驗, 只有闊葉林—針葉林邊緣鞘翅目昆蟲群落的多樣性指數與植被特征有顯著的回歸關系(表4)。植被密度是影鞘翅目昆蟲 Shannon-Wiener多樣性(r2=0.511,F=7.311,P=0.030)和豐富度(r2=0.710,F=17.169,P=0.004)的決定因素, 但植被特征對鞘翅目昆蟲群落的科均勻度和優勢度影響不大, 沒有顯著的回歸關系; 植被密度和蓋度是影響鞘翅目昆蟲總個體數量分布的重要因素(r2=0.845,F=16.387,P=0.004); 對3個優勢科進行的回歸分析表明, 叩甲的個體數量主要受植被密度的影響(r2=0.566,F=9.129,P=0.019), 瓢蟲的個體數量主要由植被密度與蓋度決定(r2=0.817,F=13.423,P=0.006), 而布甲的個體數量與植被特征之間無顯著的回歸關系, 反映出不同類群對植被特征的反應存在差異。
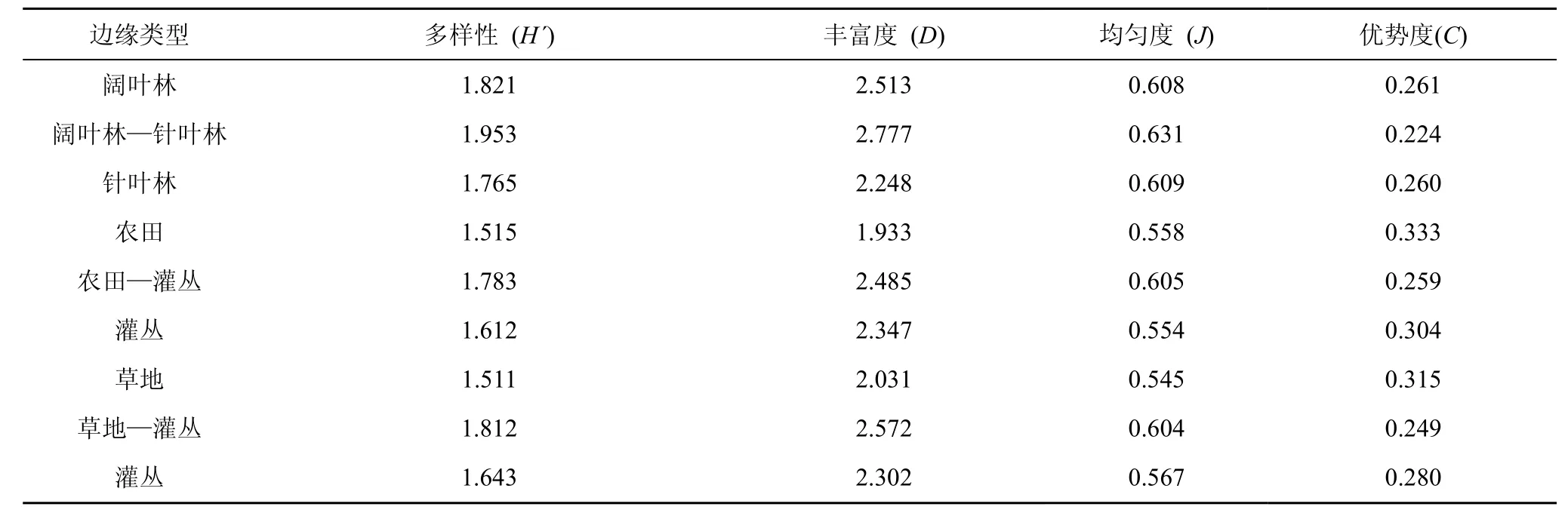
表2 不同邊緣類型鞘翅目昆蟲群落多樣性Tab.2 Comparison of Coleoptera insects community diversity indices among different type edges
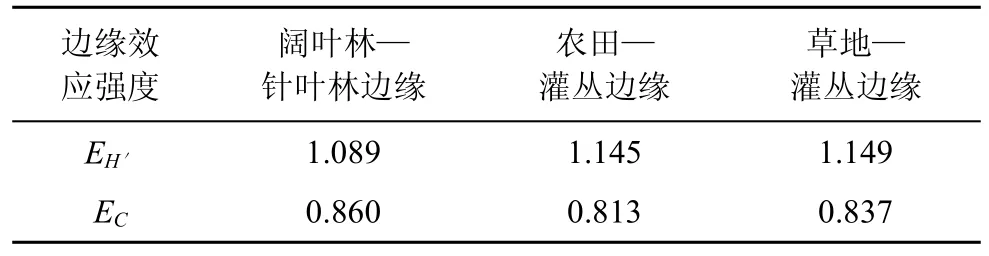
表3 不同邊緣的邊緣效應強度比較Tab.3 Comparison with the edge effect strength of different edges
4 討論
在科級水平上, 闊葉林—針葉林、農田—灌叢和草地—灌叢3種邊緣類型中鞘翅目昆蟲群落的豐富度與個體數量整體上表現為沿邊緣分別向相鄰生境內部降低的趨勢, 則對大部分鞘翅目昆蟲來說,能夠適應邊緣復雜的環境, 在邊緣很好地生存和繁殖, 但各邊緣的微環境與資源狀況不同, 使得鞘翅目昆蟲的種類和數量分布存在差異。
本文調查的 3種邊緣地帶鞘翅目昆蟲群落的Shannon-Wiener多樣性、豐富度和均勻度均高于其相鄰生境, 總體上各生境內鞘翅目昆蟲群落的多樣性指數排列順序為: 闊葉林>針葉林>灌叢>草地>農田。對比劉云慧等人研究壩上地區農田和恢復生境地表甲蟲多樣性的結果, 農田較禁牧草地和再造林地其鞘翅目科數及步甲物種數都更多, 表現出草地禁牧和再造林的生境修復活動在當前狀態下對生物多樣性恢復的作用尚不明顯[17], 此差異可能說明子午嶺保護區在其生境恢復過程中生物多樣性得到較好的恢復,但對于評估生境恢復在多樣性恢復中所起的作用,需要建立在長期監測的基礎上[18], 對此我們還需要進行長期深入地調查研究, 以期為子午嶺保護區的生態恢復提供更合理有效的措施, 提高生物多樣性恢復的效率。
邊緣效應強度EH′值均大于1, 說明3種邊緣類型均呈正的邊緣效應作用。這與王巍巍等人[4]研究荒漠景觀地表甲蟲群落的邊緣效應有正效應和負效應之分不一致, 然而實驗結果的多元化與研究對象所處的階元、邊緣類型及實驗設計等諸多因素皆有關[19]。通常在棲息地邊緣會有不同的植被結構與微環境, 如植被類型、溫度、光照、水分和土壤等生物與非生物因子[20], 這些因子將直接影響生物的取食行為與分布格局, 可能導致邊緣存在高的生物多樣性。當然任何邊緣都處于動態變化之中, 這樣的實驗結果不具有普遍代表性, 只能作為該調查區特定的生態系統管理的理論指導。因此, 對邊緣效應的動態演替進行長期監測與預報, 對邊緣效應與生物多樣性的內在關系進行本質上的探索, 是我們應該努力的方向。
調查結果顯示, 3種邊緣類型中, 只在闊葉林—針葉林邊緣鞘翅目昆蟲群落的多樣性指數與植被特征有顯著的回歸關系。植被密度影響鞘翅目昆蟲群落的 Shannon-Wiener多樣性和豐富度, 植被密度和蓋度影響鞘翅目昆蟲的個體數量分布。由此看出, 植被特征對鞘翅目昆蟲群落多樣性的影響存在差異, 不僅表現在不同邊緣類型上, 同一邊緣類型不同位置植被特征的影響也不同, 這可能與鞘翅目昆蟲不同類群對特定環境變量的選擇和敏感程度有關[21]。

表4 植被特征與鞘翅目昆蟲科多樣性、豐富度、均勻度、優勢度及個體數量間的回歸分析Tab.4 Regression analysis of vegetation characteristics and family diversity, richness, evenness, dominance and abundance of Coleoptera insects
[1]楊貴軍, 王新譜, 賈彥霞, 等.人工檸條—荒漠草地交錯帶擬步甲昆蟲群落多樣性[J].生態學報, 2016, 36(3):608–619.
[2]FLOHRE A, FISCHER C, AAVIK T, et al.Agricultural intensification and biodiversity partitioning in European landscapes comparing plants, carabids and birds[J].Ecological Applications, 2011, 21(5): 1772–1781.
[3]魯佳雄, 朱麗得孜·艾山, 伊 龍, 等.阿爾金山自然保護區鞘翅目昆蟲區系調查分析[J].干旱區研究, 2015, 32(6):1207–1212.
[4]王巍巍, 賀達漢, 張大治.荒漠景觀地表甲蟲群落邊緣效應研究[J].應用昆蟲學報, 2013, 50(5): 1383–1391.
[5]婁巧哲, 徐養誠, 馬吉宏, 等.古爾班通古特沙漠南緣地表甲蟲物種多樣性及其與環境的關系[J].生物多樣性,2011, 19 (4): 441–452.
[6]劉立品.子午嶺木本植物志[M].蘭州: 蘭州大學出版社,1998: 4–13.
[7]黨坤良, 宋小民.陜西子午嶺自然保護區綜合科學考察[M].楊凌: 西北農林科技大學出版社, 2004: 1–185.
[8]鄭樂怡, 歸鴻.昆蟲分類[M].南京: 南京師范大學出版社, 2010: 564–652.
[9]張巍巍, 李元勝.中國昆蟲生態大圖鑒[M].重慶: 重慶大學出版社, 2011: 239–378.
[10]喬治·C.麥加文.昆蟲[M].北京: 中國友誼出版公司,2007: 109–131.
[11]楊星科.中國動物志·昆蟲綱(第 61卷): 鞘翅目·葉甲科·葉甲亞科[M].北京: 科學出版社, 2014: 1–641.
[12]MEINERS S J, PICKETT S T A.Changes in community and population responses across a forest-field gradient[J].Ecography, 1999, 22(3): 261–267.
[13]HELIOLA J, KOIVULA M, NIEMELA J.Distribution of carabid beetles (Coleoptera, Carabidae) across a boreal forest–clearcut ecotone[J].Conservation Biology, 2001,15(2): 370–377.
[14]趙志模, 郭依泉.群落生態學原理與方法[M].重慶: 文獻和科學技術出版社, 1990: 147–279.
[15]馬克平, 劉玉明.生物群落多樣性的測度方法: Ⅰα多樣性的測度方法(下)[J].生物多樣性, 1994, 2(4): 231–239.
[16]王伯蓀, 彭少麟.鼎湖山森林群落分析——Ⅹ.邊緣效應[J].中山大學學報自然科學版, 1986, (4): 52–56.
[17]劉云慧, 宇振榮, 王長柳, 等.壩上地區農田和恢復生境地表甲蟲多樣性[J].生態學報, 2011, 31(2): 0465–0473.
[18]DAVIS C A, UTRUP J S.Response of terrestrial invertebrates to high-and low-diversity grassland restorations in south-central Nebraska[J].Restoration Ecology, 2010,18(S2): 479–488.
[19]廉振民, 于廣志.邊緣效應與生物多樣性[J].生物多樣性, 2000, 8(1): 120–125.
[20]楊貴軍, 賈彥霞, 王新譜.苜蓿—荒漠草地交錯帶步甲昆蟲多樣性[J].環境昆蟲學報, 2015, 37(3): 483–491.
[21]HUMPHREY J W, HAWES C, PEACE A J, et al.Relationships between insect diversity and habitat characteristics in plantation forests[J].Forest Ecology & Management,1999, 113(1): 11–21.
Edge effects of Coleoptera insect communities in Ziwuling National Nature Reserve, Shaanxi
XU Shicai*, WANG Lijuan
College of Life Sciences,Yan’an University,Yan’an Shaanxi716000,China
Three edge zones, broad-leaved-coniferous forest, farmland-shrub and grassland-shrub of Ziwuling National Nature Reserve in Shaanxi Province were selected as the sample plots, and the impact of the edge effect on the diversity of Coleopteran insect community was discussed at family level.A total of 4935 Coleopteran insects were collected, belonging to 28 families, among which the individual amount of Carabidae, Chrysomelidae and Coccinellidae accounted for 44.19%,21.95% and 13.66% of the total number of individuals respectively, which constituted the dominant group of Coleopteran insects in this area.The family diversity, richness and evenness of Coleopteran insects in the three edge types decreased from the edge to the adjacent habitats, with theEH′values greater than 1, showing a positive edge effect.The multiple regression analysis showed that the vegetation density affected the diversity and richness of Coleopteran insect community in the broad-leaved-coniferous forest, and the vegetation density and coverage affected the individual population distribution of the entire Coleopteran insects community.The above conclusions provide scientific evidence for the conservation and rational utilization of the biodiversity of Coleopteran insects in Shaanxi Ziwuling National Nature Reserve.
Coleoptera; community diversity; edge effect
10.14108/j.cnki.1008-8873.2017.05.026
Q968.1
A
1008-8873(2017)05-204-07
徐世才, 王麗娟.陜西子午嶺國家級自然保護區鞘翅目昆蟲群落的邊緣效應[J].生態科學, 2017, 36(5): 204-210.
XU Shicai, WANG Lijuan.Edge effects of Coleoptera insect communities in Ziwuling National Nature Reserve, Shaanxi[J].Ecological Science, 2017, 36(5): 204-210.
2017-04-14;
2017-06-12
陜西省高水平大學建設專項資金項目“生態學”(2012SXTS03); 延安大學引導項目“陜北煙區煙粉虱的發生規律及其有機防控”(YD2015-11)
徐世才(1973—), 男, 陜西延安人, 碩士, 副教授, 碩士生導師, 主要從事昆蟲生態與綜合防治研究, E-mail: shicaixu@163.com
*通信作者:徐世才
