非靶標代謝組研究溫度對黃曲霉菌代謝的影響
2017-11-29 08:15:16謝華里李培武王秀嬪張良曉汪雪芳
分析測試學報 2017年11期
謝華里,李培武,5*,王秀嬪,張 奇,張良曉,王 同,張 文,5,汪雪芳,5
(1.中國農業科學院油料作物研究所,湖北 武漢 430062;2.農業部油料作物生物學與遺傳育種重點實驗室,湖北 武漢 430062;3.農業部生物毒素檢測重點實驗室,湖北 武漢 430062;4.農業部油料產品質量安全風險評估實驗室,湖北 武漢 430062;5.農業部油料及制品質量監督檢驗測試中心,湖北 武漢 430062)
非靶標代謝組研究溫度對黃曲霉菌代謝的影響
謝華里1,2,3,4,李培武1,2,3,4,5*,王秀嬪1,3,4,5*,張 奇1,2,3,4,張良曉1,3,4,5,王 同1,2,3,張 文1,2,3,5,汪雪芳1,2,3,5
(1.中國農業科學院油料作物研究所,湖北 武漢 430062;2.農業部油料作物生物學與遺傳育種重點實驗室,湖北 武漢 430062;3.農業部生物毒素檢測重點實驗室,湖北 武漢 430062;4.農業部油料產品質量安全風險評估實驗室,湖北 武漢 430062;5.農業部油料及制品質量監督檢驗測試中心,湖北 武漢 430062)
采用基于超高效液相色譜-高分辨質譜聯用的非靶標代謝組學方法來研究溫度對黃曲霉菌生理代謝的影響,使用交互偏最小二乘判別分析(OrthoPLS-DA)等化學計量學方法對代謝組數據進行多元統計分析,使用二級質譜信息和譜庫檢索定性黃曲霉菌代謝特征信息。使用內標結合混合質控樣品的方法對非靶標代謝組方法進行質量控制。將該方法應用于研究溫度對黃曲霉菌代謝組的影響,發現不同溫度下有3 593個(T檢驗p<0.01)差異表達代謝特征,篩選出20個候選差異代謝物。研究結果表明,溫度顯著影響三羧酸循環、脂肪酸、苯丙氨酸、色氨酸、絡氨酸等生物合成路徑,并調控黃曲霉毒素、黃匹阿尼酸、曲酸等次生代謝物生物合成路徑酶活性。研究發現曲酸和黃曲霉毒素前體化合物與黃曲霉毒素的累積變化規律相似,可作為候選靶標進行驗證。該研究為開展我國黃曲霉毒素風險評估和分子預警研究提供了新的路徑和方法。
黃曲霉菌;代謝組;高分辨質譜;黃曲霉毒素生物合成;溫度
黃曲霉毒素是迄今發現毒性最強的一類污染花生和玉米等農產品的生物毒素,對我國農產品質量安全構成了很大威脅,特別是近年來對花生等油料農產品的出口貿易造成重大影響。當前,研究已經明確遺傳、生物和非生物因素將影響黃曲霉毒素的形成,并已明確了參與黃曲霉毒素生物合成路徑上的骨架基因,調控因子及生物合成酶[1]。此外黃曲霉菌全基因組測序已經完成,也對其次生代謝基因簇進行了生物信息學預測[2],黃曲霉毒素生物合成區室化分布[3]和光照如何調控黃曲霉毒素合成也已報道[4],這些研究基礎為基于質譜技術結合基因挖掘策略進行黃曲霉次生代謝研究提供了較為充足的理論基礎。溫度和水分活度是生產和糧油倉儲中影響黃曲霉毒素產生最重要的環境因子。當前對于以溫度調控黃曲霉毒素形成的機制仍不明確,僅停留于最適宜溫度為28~30 ℃,低溫(<20 ℃)和高溫(>35 ℃)抑制黃曲霉毒素產生的表觀層面。而通過研究溫度顯著調控的靶點,可為黃曲霉毒素的防控提供參考靶點和理論支撐,也為開展黃曲霉污染早期分子預警和風險評估研究提供技術支撐。
代謝組學(Metabolomics或Metabonomics)是繼基因組學、轉錄組學和蛋白質組學等系統生物學之后興起的一個分支[5]。當前代謝組學主要分為靶向代謝組學和非靶向代謝組學,靶向代謝組學是針對特定數量或種類的代謝分析策略,具有主觀目的性。而非靶向代謝組學是一種盡可能多的檢測樣本中代謝物的分析策略,從總體上對生物學問題在代謝水平上實現導航,具有無偏性。由此可見,非靶向代謝組學研究的代謝物集合更大,這對分析儀器平臺提出了更高的精度要求。目前,代謝組學主要基于核磁共振和質譜分析平臺并結合化學計量學進行代謝差異研究。其中高分辨質譜在非靶向代謝分析方面扮演著重要的角色。該平臺主要通過一級精確質量數和二級質譜數據作為定性判據,為分析海量的代謝信息提供高分辨工具平臺。目前主流的高分辨質譜有飛行時間質譜(TOF-MS)、靜電場軌道阱(Orbitrap-MS)質譜和傅立葉變換離子回旋共振FTICR-MS質譜。靜電場軌道阱(Orbitrap-MS)質譜(MS error<2 ppm)相比于飛行時間(TOF-MS)質譜(MS error<5 ppm)具有更低的測量質量誤差,同時在儀器穩定性和重現性上顯著優于飛行時間(TOF-MS)質譜[6],FTICR-MS雖是目前測量精度最高的質譜,但其掃描速度較前兩者更慢,在代謝組學研究中應用較少。此外,將高分辨質譜技術應用于黃曲霉菌等真菌的研究報道較為缺乏,主要集中在利用高分辨質譜技術對黃曲霉菌新次生代謝物通路的結構鑒定研究方面[7-8]。
本文通過利用代謝組研究建立以黃曲霉菌為代表的絲狀真菌的非靶標代謝組學方法,并利用該方法研究溫度對黃曲霉菌生理代謝的影響,為深入理解黃曲霉菌響應環境因子機制提供一個有效的方法途徑。
1 實驗部分
1.1 試劑與材料
甲醇(MeOH)、乙酸乙酯(EA)、二氯甲烷(DCM)、乙腈(ACN)(色譜純,美國Fisher 公司),甲酸購于Sigma-Aldrich 公司(Bornem,Belgium)。2-氯苯丙氨酸購于百靈威公司。超純水通過Milli-Q Gradient System(Millipore,Brussels,Belgium)產生。PDA固體培養基(200 g馬鈴薯,洗凈去皮切碎,加水1 000 mL蒸煮0.5 h,紗布過濾,再加葡萄糖10 g,充分溶解后趁熱用紗布過濾,分裝于三角錐形瓶中,加入瓊脂15 g,搖勻。121 ℃滅菌20 min,冷卻后貯存備用)。沙氏培養基購于海博生物公司。冷甘油緩沖溶液(-30 ℃):丙三醇和NaCl溶液(13.5 g/L)以3∶2(體積比)混合;清洗溶液:丙三醇和NaCl溶液(13.5 g/L)以1∶1(體積比)混合。PBS緩沖溶液:0.01 mol/L PBS緩沖液(pH 7.4):稱取NaCl 8.0 g、Na2HPO4·12H2O 2.9 g、KCl 0.2 g、KH2PO40.2 g溶于900 mL去離子水中,用鹽酸或氫氧化鈉溶液調至pH 7.4,補加去離子水并定容至1 L。
1.2 菌株及培養條件
AF73菌株由中國農業科學院油料作物研究所質檢中心(武漢)提供。黃曲霉菌在PDA固體培養基上28 ℃培養10 d后,用無菌水(含0.1%的吐溫-80)將孢子從培養基上洗下,用血球板計數法計數。沙氏液體培養基中接種黃曲霉孢子懸液,使接種量達5×105個/mL,將培養瓶置于28 ℃恒溫搖床(200 r/min)培養6 d,得到黃曲霉菌樣品。
1.2 取樣與樣品準備
1.2.1黃曲霉菌猝滅方法采用改進的冷甘油緩沖溶液法猝滅[9],具體步驟為:快速轉移黃曲霉菌株樣品到預先加入冷甘油緩沖溶液猝滅劑的50 mL離心管中,混合液中樣品體積和猝滅劑體積比保持在1∶4~1∶6之間。快速均質混合液(渦旋10 s)后,置于 -30 ℃冷卻5 min。真空抽濾,快速過濾猝滅樣品,吸取5 mL PBS清洗黃曲霉菌絲球,重復1次,收集得到樣品,冷凍干燥8 h。
1.2.2黃曲霉菌細胞被膜破碎方法及代謝物提取采用非機械破碎方法中的有機溶劑穿透破碎結合超聲機械破碎對細胞進行破碎和代謝物提取。提取溶劑為混合溶劑MeOH-DCM-EA(1∶1∶1,體積比),同時加入1%甲酸以提高化合物離子化效率,加入內標2-氯苯丙氨酸配成500 μg/L溶液。混合提取溶液配制好后,取2 mL提取液加入樣品中進行代謝物超聲(超聲功率100%)提取20 min。
1.3 UHPLC-HRMS儀器條件
在高效液相色譜(Dionex,Sunnyvale,CA,USA)上進行色譜分離,色譜柱為C18反向色譜柱(Hypersil Gold,100 mm×2.1 mm i.d.3 μm,Thermo Fisher Scientific,USA)。使用Orbitrap Fusion靜電軌道阱高分辨質譜(Thermo Scientific,USA)做質譜分析。
流動相:A為甲醇-水(95∶5,體積比,含0.1%甲酸和10 mmol/L甲酸銨)混合溶液;B為水-甲醇(95∶5,體積比,含0.1%甲酸和10 mmol/L甲酸銨)混合溶液。梯度洗脫程序為:0~2 min,85%B;2~8 min,85%~50%B;8~10 min,50%B;10~12 min,50%~30%B;12~13 min,30%B;13~15 min,30%~0%B;15~16 min,0%B;16~24 min,0%~85%B;24~28 min,85%B。流速為0.3 mL/min。柱溫為40 ℃,自動進樣室溫度為15 ℃,進樣量為2 μL。
高分辨質譜條件:離子源加熱溫度為300 ℃;噴霧電壓:正離子模式下為3.5 kV,負離子模式下為3.0 kV;鞘氣為40 Arb;輔氣為5 Arb。離子傳輸毛細管溫度為320 ℃,毛細管電壓-1.9 kV。主要的一級精確質量數全掃參數如下:檢測器選用Orbitrap,分辨率選擇120 000 FWHM(半峰寬),掃描范圍為100~1 000m/z,自動增益控制設定為1.0e6,注入時間為100 ms。一級掃描與二級掃描間主要的過濾參數如下:強度閾值為1.0e4,帶電荷為1~2個,動態排除設定為1。數據依賴采集選擇Top speed模式,循環時間為1 s。主要的二級質譜掃描(dd-ms2)參數如下:碎裂模式為選擇高能碰撞誘導裂解模式(HCD),碰撞能正離子模式下設為35 eV,負離子模式設為30 eV。檢測器類型為Orbitrap,分辨率設為30 000 FWHM,自動增益控制設為5.0e4,最大注入時間為100 ms,四極桿隔離寬度為1 Da。
1.4 數據預處理與多元統計分析
使用SIEVE 3.0(Thermo Scientific,USA)對原始數據分別進行峰對齊,提取,代謝物譜庫檢索定性得到原始峰表。經過處理后導入SIMCA14.1或在線代謝組學數據處理網站MetaboAnalyst 3.0(www.metaboanalyst.ca)[10]或XCMS[11](https://xcmsonline.scripps.edu)進行數據過濾、標準化等數據預處理和多元統計分析。使用代謝組數據庫METLIN(https://metlin.scripps.edu/)、mzCloud(https://www.mzcloud.org/)、YMDB( http://www.ymdb.ca/)和KEGG (http://www. kegg. com/ )和本地庫進行代謝物定性。
1.5 溫度對黃曲霉菌代謝組的影響
分別以不同溫度(18、28、38 ℃)培養的3組黃曲霉菌作為樣品,使用上述的前處理步驟、檢測條件和數據處理方法,對黃曲霉菌生理及次生代謝情況進行全組分分析,以期初步揭示不同溫度條件對黃曲霉生理及其產毒的影響機制。
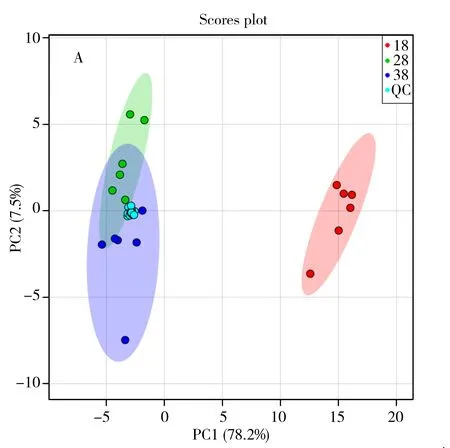
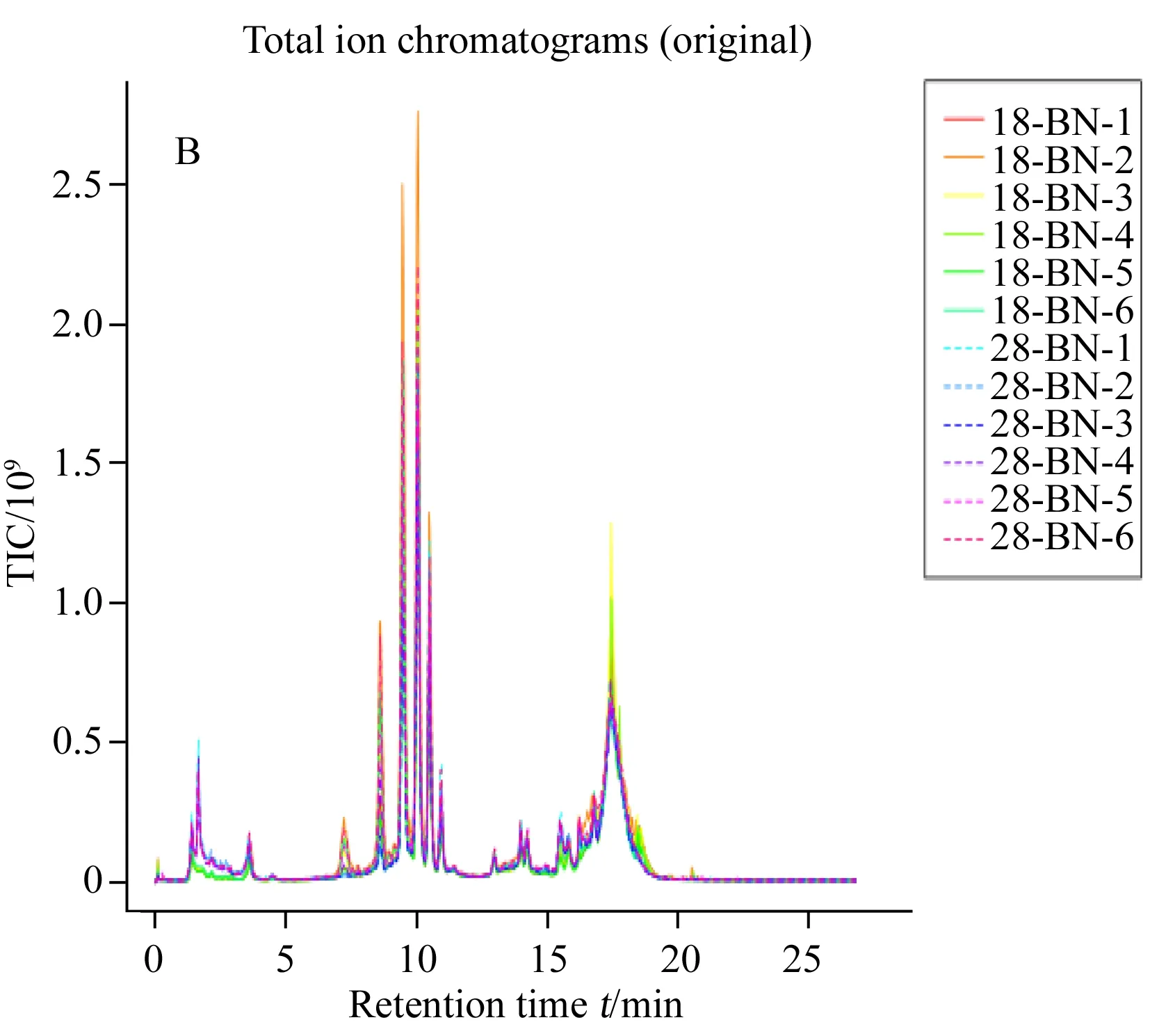
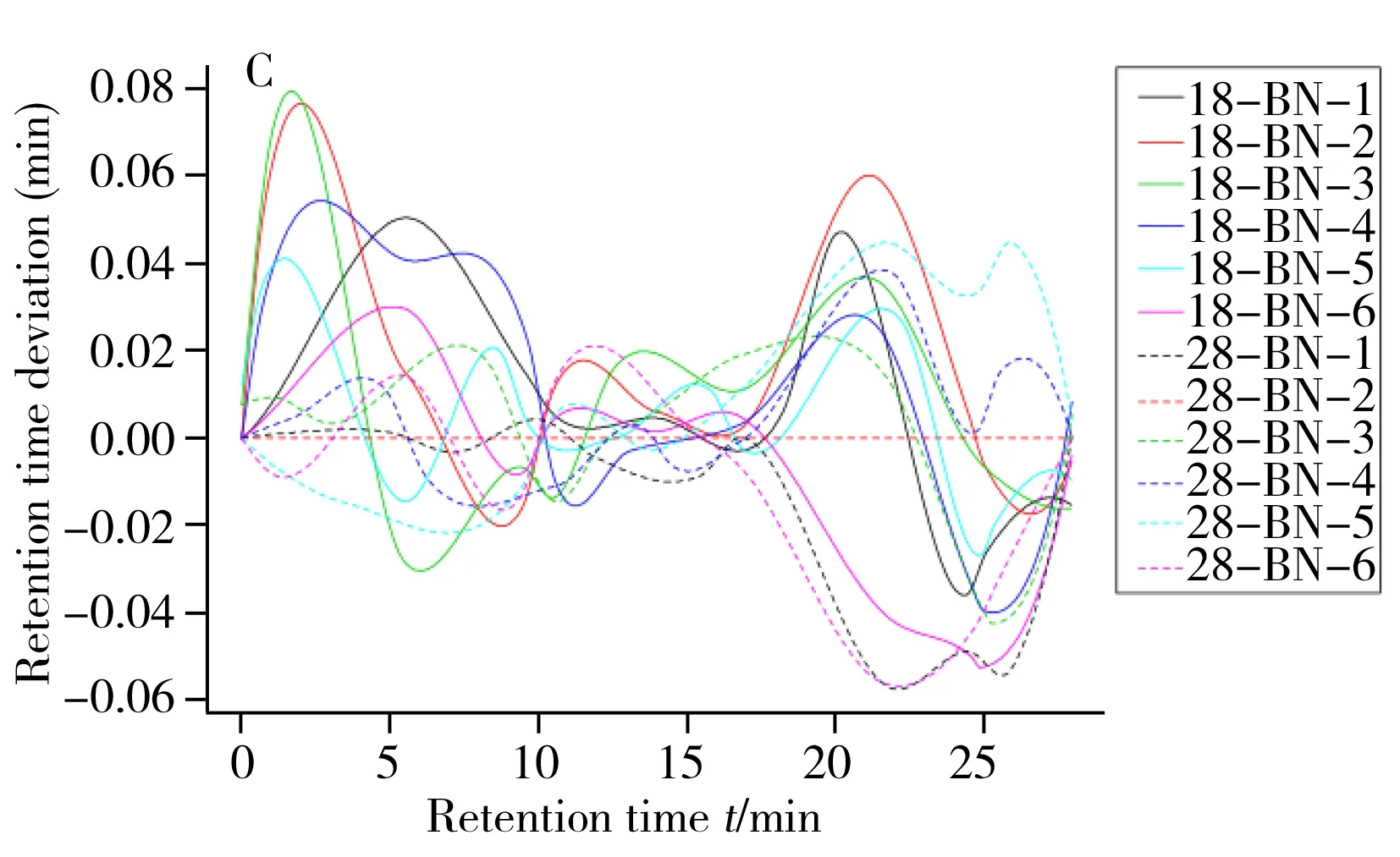
圖1 非靶標代謝組方法確證圖示Fig.1 Method validation plot involved in non-targeted metabolomicsaA.rincipal component analysis scores plot of three temperature and quality control sample;B.8 ℃ and 28 ℃ groups of samples were aligned with the original peak of total ion current;C.lot of retention time deviation for 18 ℃ and 28 ℃ groups of samples;18-BN and 28-BN represent the intracellular group of 18 ℃ and 28 ℃(18-BN和28-BN代表18,28 ℃組樣品胞內的6個重復)
2 結果與討論
2.1 非靶標代謝組學方法確證
非靶標代謝組學方法分析目標是數以千計的未知形式代謝物,其方法確證有別于目標物分析方法確證,更具挑戰性[12]。為確保研究發現的差異是生物學差異而非測定誤差帶來的技術差異,需進行有效的質量控制。目前國際上常用方法是使用混合QC質控樣品,即從樣本中取少量、等量混勻用作QC樣本,將其插入運行序列中用于監測儀器穩定性和可重復性并用于數據過濾[13]。本研究使用2-氯苯丙氨酸作為內標結合混合QC質控樣品對數據質量進行監測,并用于方法確證[12,14]。分析結束后使用QC樣品數據進行以下工作:對分析性能進行系統評價,判斷進一步數據分析的必要性;進行代謝特征選取,濾除在QC 中變異大的代謝特征,通常LC-MS分析中過濾閾值RSD為20%[13,15-16]。將3種不同溫度培養條件下樣品(分胞內和胞外樣品共36個)、3個空白樣本和17個混合質控樣,共56個原始樣本數據經過數據預處理后進行主成分分析,結果見圖1A。由圖可見,17個質控樣品較好地聚集在原點附近,圖1B總離子流圖中原始峰對齊效果也較好,未出現顯著的峰漂移,由圖1C可見保留時間變異在±0.08 s內,以上數據表明儀器的穩定性和重現性較好。
2.2全組分分析溫度對黃曲霉菌生理及其產毒的影響
2.2.1溫度對黃曲霉菌生理代謝輪廓分析采用UHPLC-HRMS分別在正負離子模式下對樣本進行數據采集,在18、28、38 ℃培養條件下樣本分別用正負離子模式采集到總離子流圖(TIC)。從總離子流圖可以總體上觀察3個溫度下的樣品是否存在差異,但具體差異需進一步提取分析。
2.2.2潛在差異代謝物篩選由圖2可見,質荷比分布云圖可以對采集到的特征化合物質量分布和出峰時間有直觀了解。在p≤0.01和差異表達倍數≥1.5情況下發現3 593個特征化合物,圖中包含p值可視化、定向差異表達倍數值和特征化合物質荷比信息,且包含了每個樣本的總離子流圖和保留時間。圖中上半部分綠色部分代表化合物強度增強的組分,下半部分代表了化合物減弱的部分,氣泡的大小與特征值的差異表達倍數取log值后的大小相關,即氣泡大者,差異表達倍數取log值后也大。本研究中,特征化合物主要分布為出峰時間較早的極性化合物,質量分布在100~600 Da左右,5~10 min出峰的中等極性化合物質量分布在200~800 Da左右,10~18 min出峰的弱極性化合物質量分布在200~1 000 Da。從氣泡大小可看出,大部分特征化合物的差異表達倍數(Fold change)值較大,表明篩選的特征化合物差異顯著。
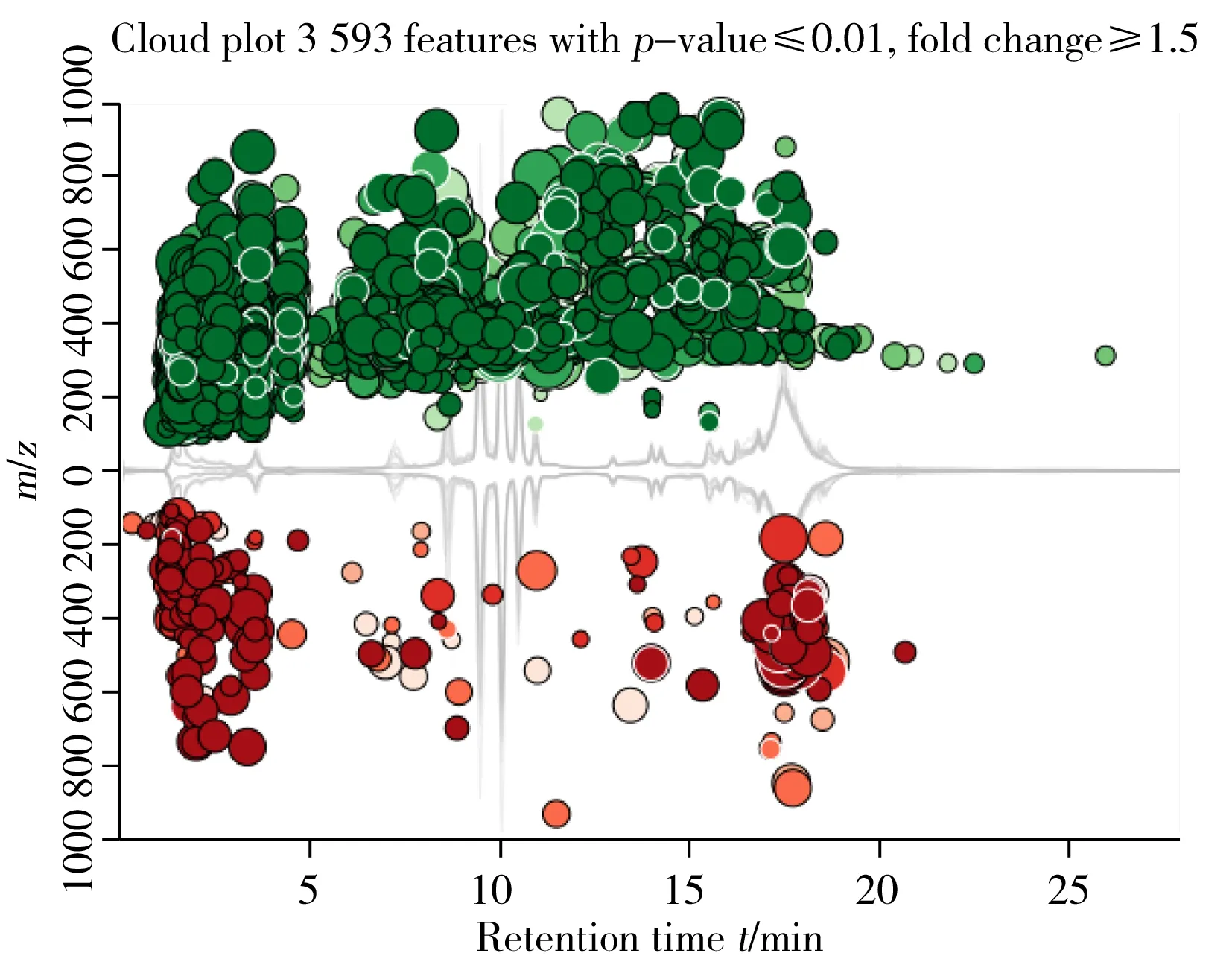
圖2 黃曲霉菌數據集質荷比分布云圖Fig.2 Cloud plot of aspergillus flavus data set
OrthoPLS-DA是一種有監督的模式識別分析方法,與PCA相比,可在不降低模型預測能力的前提下,增強模型的解釋能力。本研究中,對18 ℃與28 ℃培養樣品進行OrthoPLS-DA分析比較,得到該2個溫度下培養組的OrthoPLS-DA得分圖3A,同樣,38 ℃與28 ℃培養組的比較也經過OrthoPLSDA分析,其得分圖見圖4A。從圖3A,4A可看出,這兩組都可以與28 ℃培養組明顯區分。R2Y、Q2分別代表模型OrthoPLS-DA的解釋能力和預測能力。本研究中18、28 ℃培養組模型的R2Y、Q2分別為0.98和0.96。28、38 ℃培養組模型的R2Y、Q2分別為0.99和0.92。說明兩個模型都具有較高的解釋能力和預測能力,分類效果明顯。由于OrthoPLS-DA分析模型易出現過擬合,常采用如交叉檢驗(Cross-validation)、置換檢驗(Permutation test)或刀切法(Jackknife)來檢驗模型是否過擬合。本研究采用200次置換檢驗對模型進行驗證(圖5)。在置換檢驗驗證中,R2-intercept和Q2-intercept兩個參數用來檢驗OrthoPLS模型是否過擬合,經驗表明,R2-intercept值不宜超過0.3~0.4,Q2-intercept不宜超過0.05。而對于OrthoPLS-DA模型是否過擬合,主要以Q2-intercept值為依據[18]。本研究中,18、28 ℃培養組置換檢驗(Permutation test)得到的R2-intercept和Q2-intercept值分別為0.607和-0.818。28、38 ℃培養組置換檢驗(Permutation test)得到的R2-intercept和Q2-intercept值分別為0.986和-0.252。從圖5可見,兩組中所有R2-intercept和Q2-intercept左邊的點均低于右邊的點,表明模型的預測能力大于任何一次隨機排列y變量的預測能力,回歸線的斜率也大于1,證明建立的兩個OrthoPLS-DA模型沒有出現過擬合。同時,對兩個OrthoPLS-DA模型進行CV-ANOVA驗證,p<0.001(分別為0.000 8和5.18×10-5),這說明采用OrthoPLS-DA模型篩選本研究中的差異代謝物具有統計學依據,可繼續進行后續差異代謝物篩選。
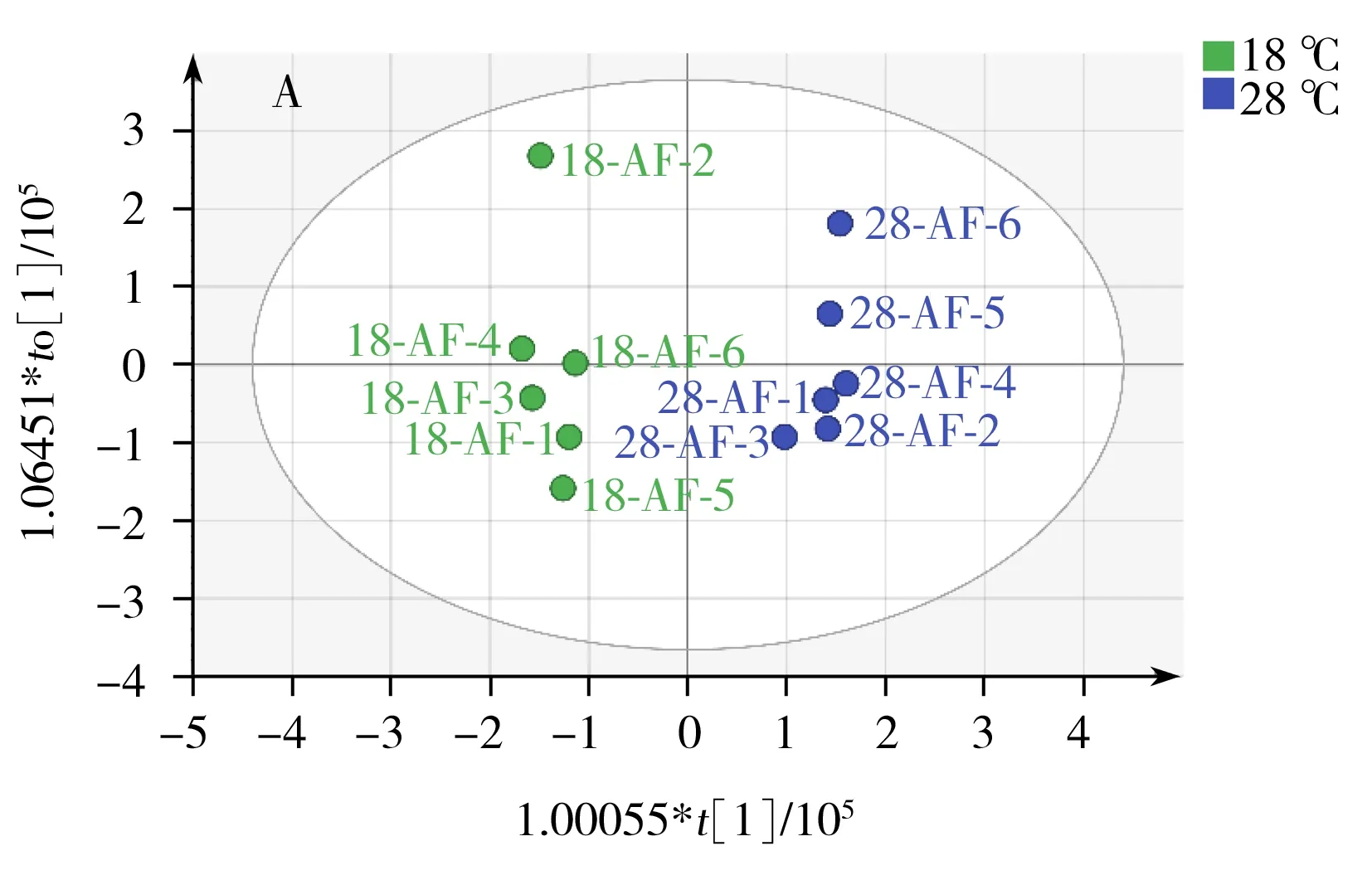

圖3 18 ℃和28 ℃培養組的OrthoPLS-DA分析得分圖(A)和差異化合物特征變量圖(S-plot)(B)Fig.3 OrthoPLS-DA score plot of 18 ℃group and 28 ℃group(A) and the plot of feature importance(S-plot)(B)
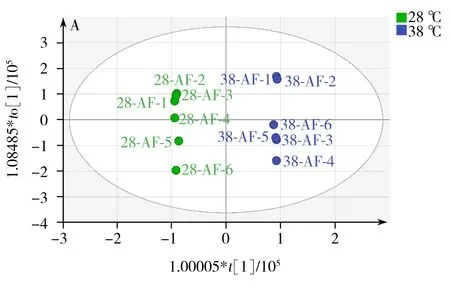
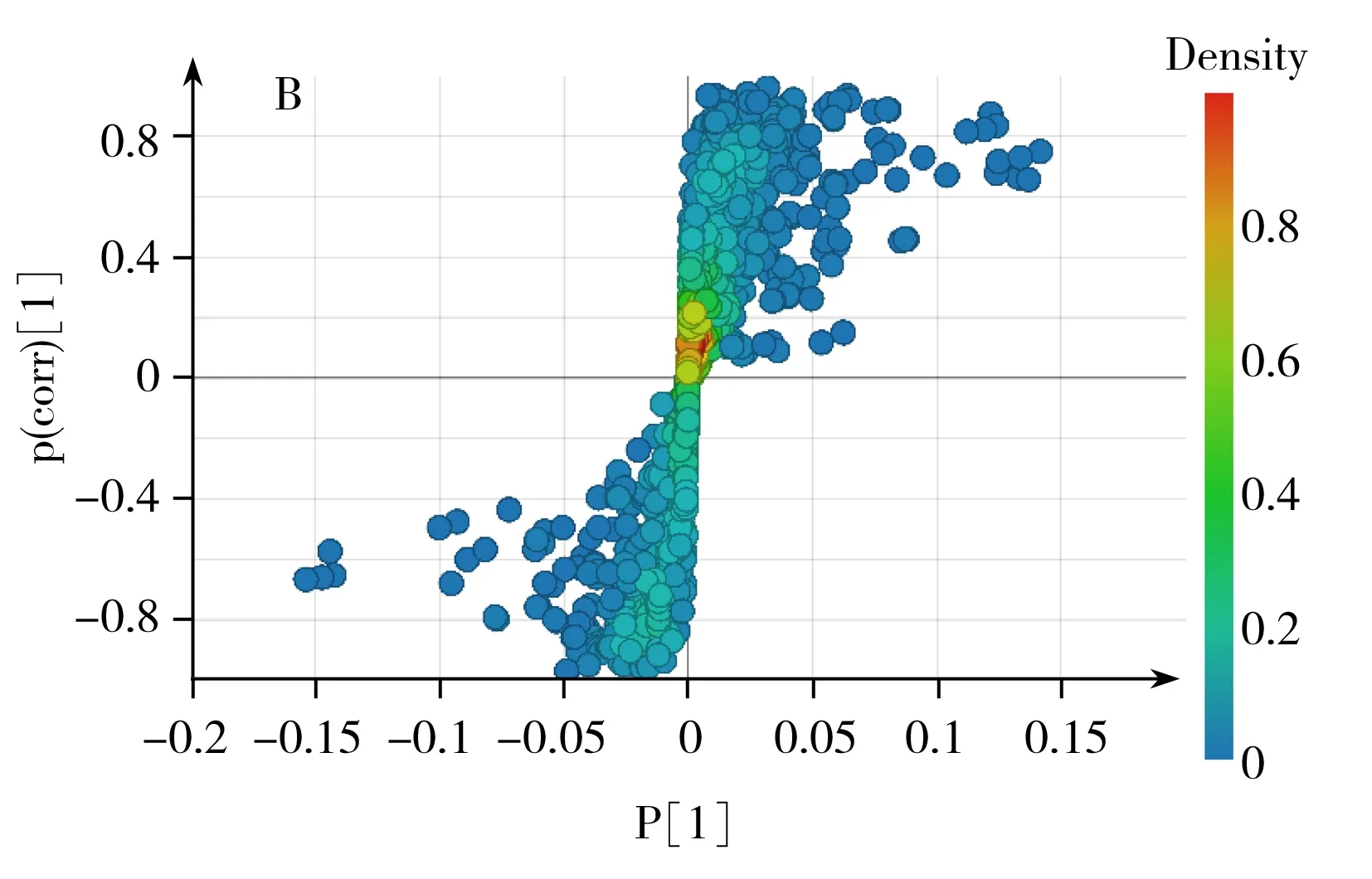
圖4 38 ℃和28 ℃培養組的OrthoPLS-DA分析得分圖(A)和差異化合物特征變量圖(S-plot)(B)
Fig.4 OrthoPLS-DA score plot of 38 ℃group and 28 ℃group(A) and the plot of feature importance(S-plot)(B)
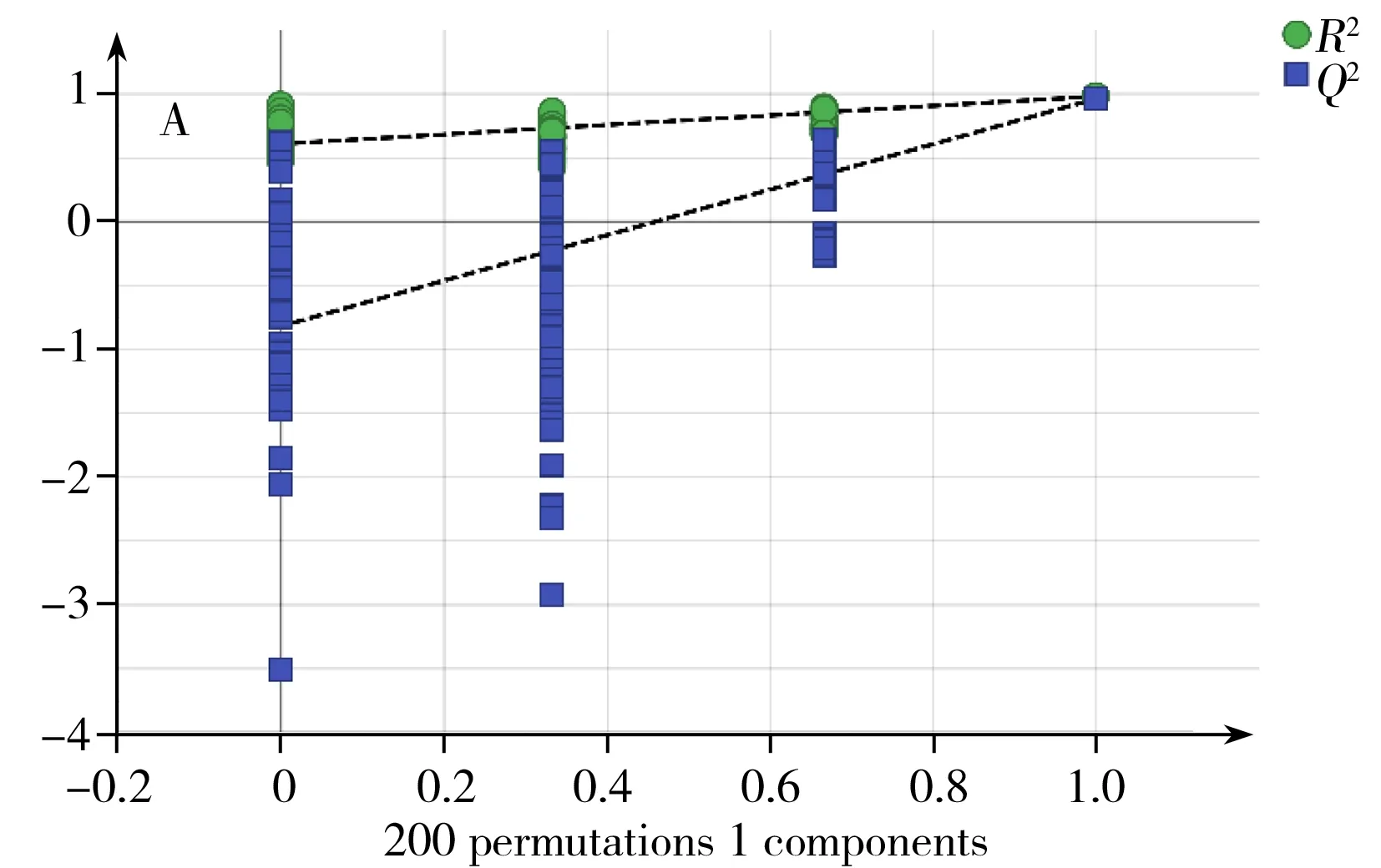
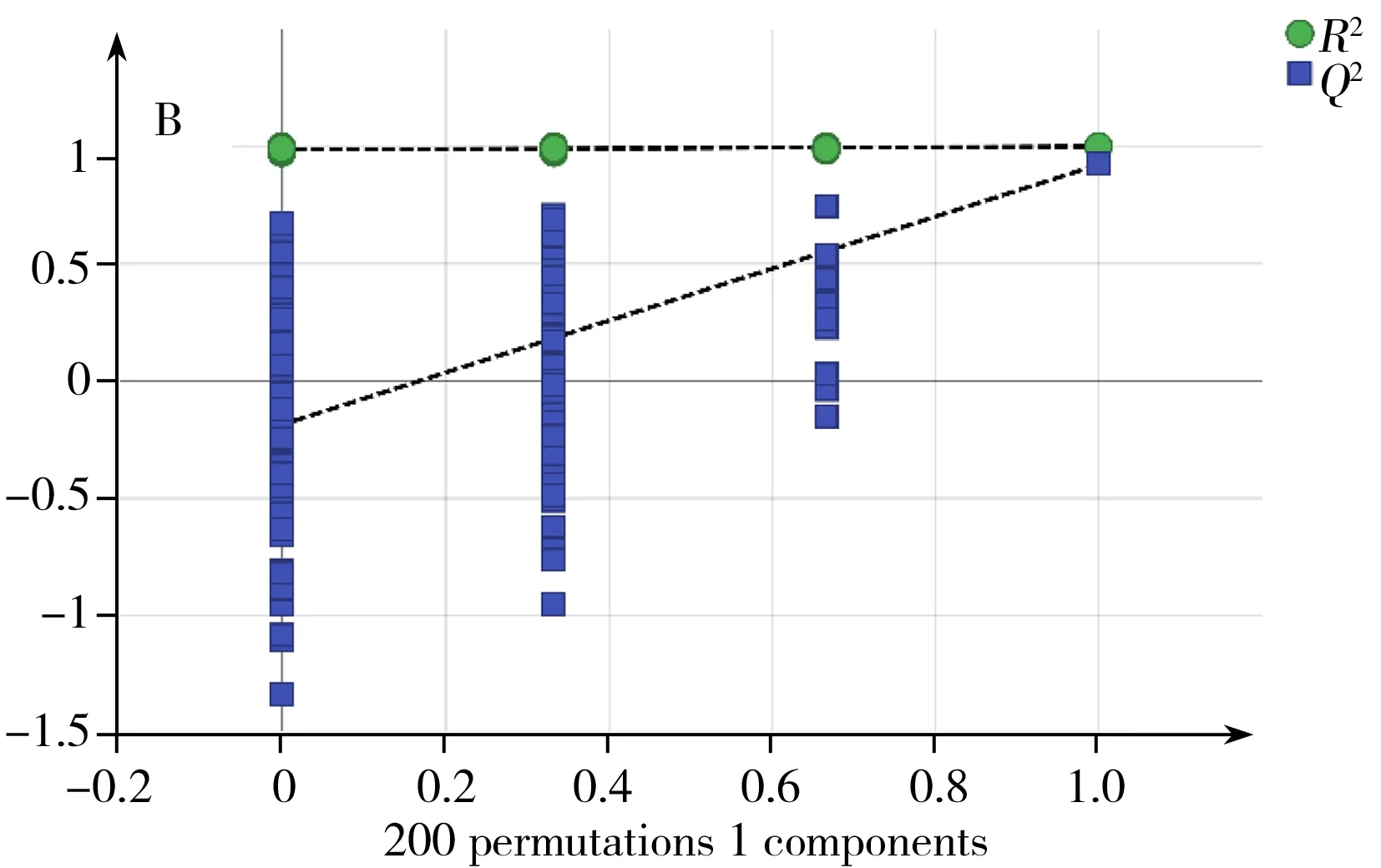
圖5 18 ℃和28 ℃培養組(A)以及28 ℃和38 ℃培養組(B)的OrthoPLS-DA模型驗證圖Fig.5 OrthoPLS-DA permutationtest model validation plot of 18 ℃group and 28 ℃group(A) and 28 ℃group and 38 ℃group(B)
2.2.3潛在差異代謝物鑒定對重要特征變量進行選取,其中圖3B和4B圖中遠離原點的化合物為差異顯著的潛在的差異代謝物,同時以有統計學意義的變量(p<0.05)作為差異化合物判斷標準,并通過MZcloud、Metlin和本地庫等數據庫進行二級質譜信息匹配及部分標準品驗證,篩選出20個溫度影響黃曲霉菌生理代謝的潛在差異代謝物(表1),其中纈氨酸、亮氨酸、苯丙氨酸、色氨酸、酪氨酸、硬脂酸、油酸、曲酸、黃匹阿尼酸、3a,3a′-二聚吡咯吲哚生物堿、黃曲霉毒素B1、黃曲霉毒素B2、氧甲基雜色曲霉毒素、降散盤衣酸、雜色曲菌素 B經過標品驗證,對其相對含量進行了比較分析(圖6)。
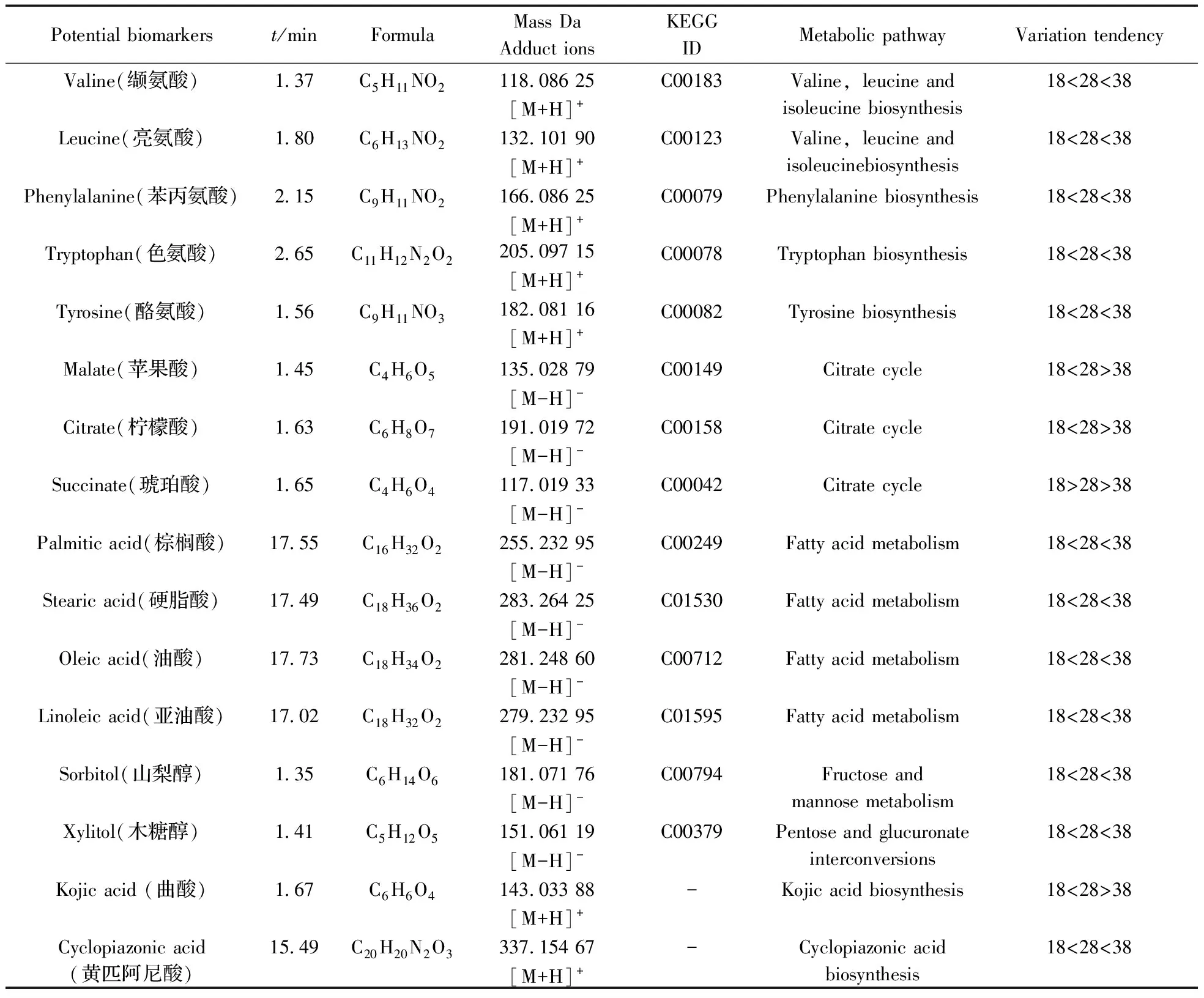
表1 溫度影響黃曲霉菌生理代謝的潛在差異代謝物Table 1 Potential biomarkers of physiological metabolism of aspergillus flavus affected by temperature
(續表1)
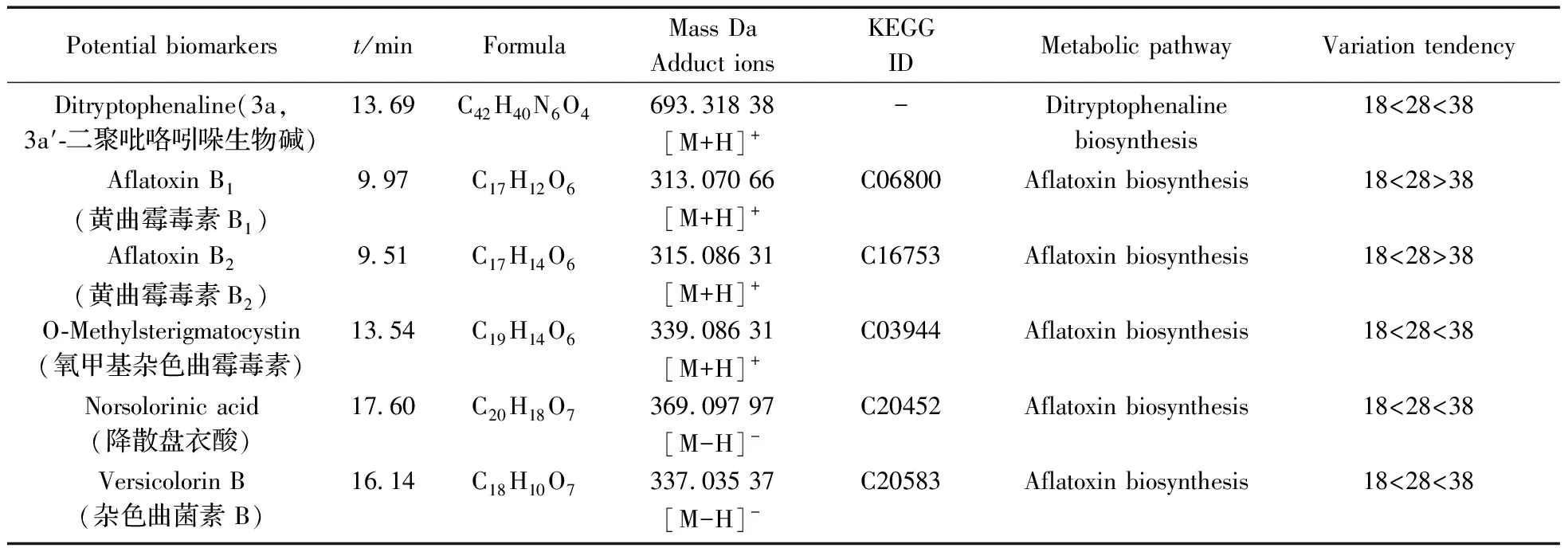
Potentialbiomarkerst/minFormulaMassDaAdductionsKEGGIDMetabolicpathwayVariationtendencyDitryptophenaline(3a,3a′?二聚吡咯吲哚生物堿)13 69C42H40N6O4693 31838[M+H]+-Ditryptophenalinebiosynthesis18<28<38AflatoxinB1(黃曲霉毒素B1)9 97C17H12O6313 07066[M+H]+C06800Aflatoxinbiosynthesis18<28>38AflatoxinB2(黃曲霉毒素B2)9 51C17H14O6315 08631[M+H]+C16753Aflatoxinbiosynthesis18<28>38O?Methylsterigmatocystin(氧甲基雜色曲霉毒素)13 54C19H14O6339 08631[M+H]+C03944Aflatoxinbiosynthesis18<28<38Norsolorinicacid(降散盤衣酸)17 60C20H18O7369 09797[M-H]-C20452Aflatoxinbiosynthesis18<28<38VersicolorinB(雜色曲菌素B)16 14C18H10O7337 03537[M-H]-C20583Aflatoxinbiosynthesis18<28<38
-:no data
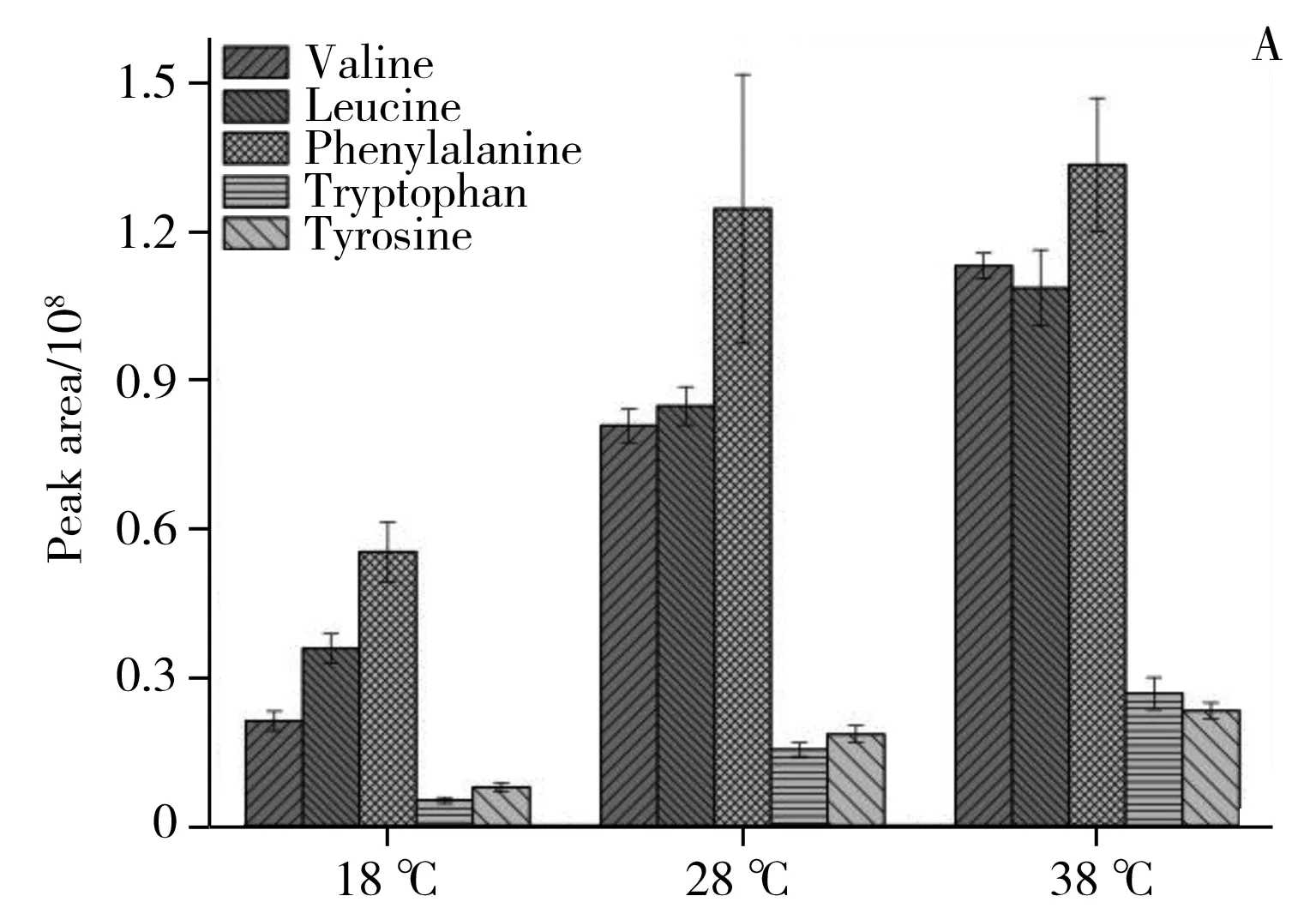
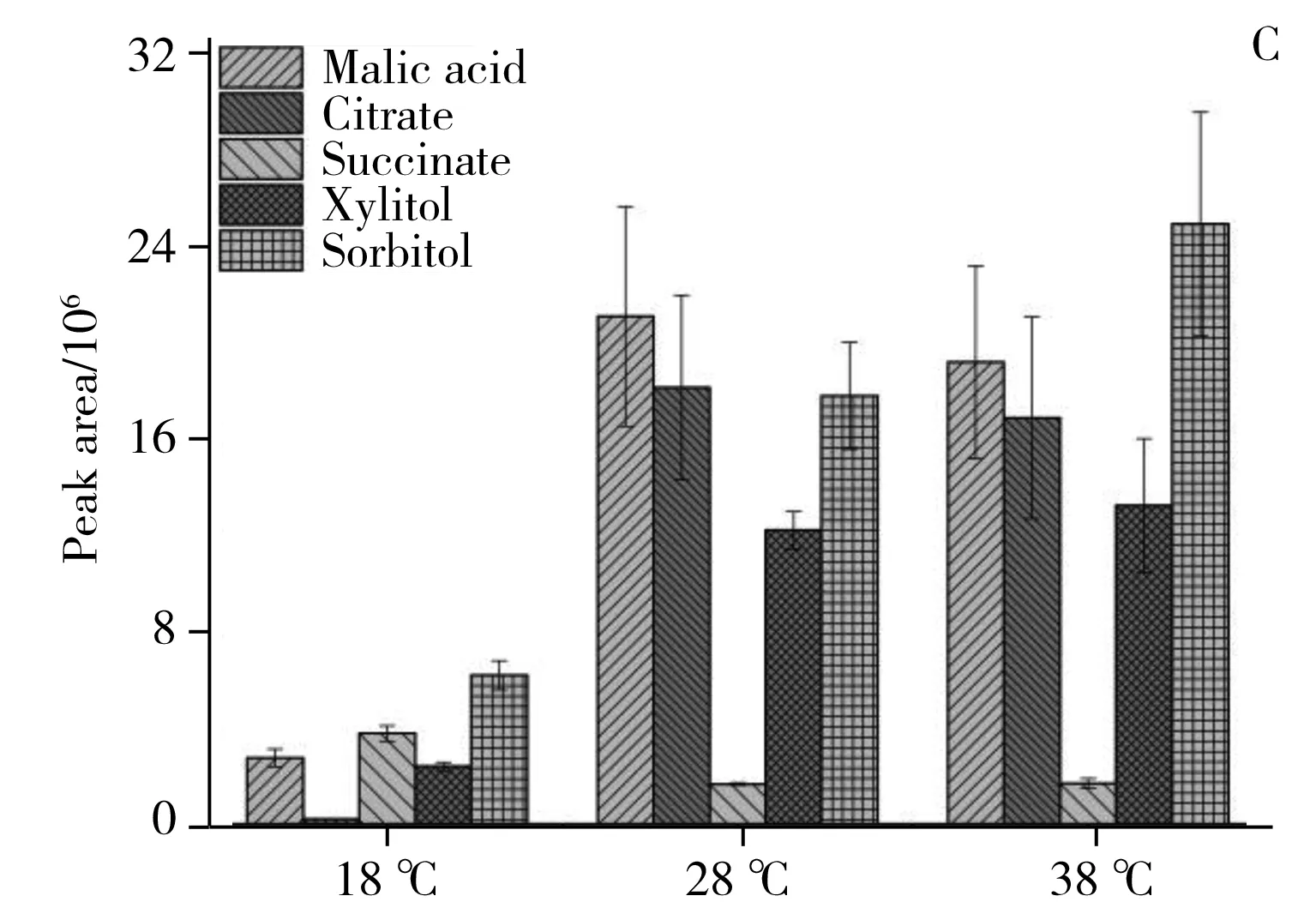
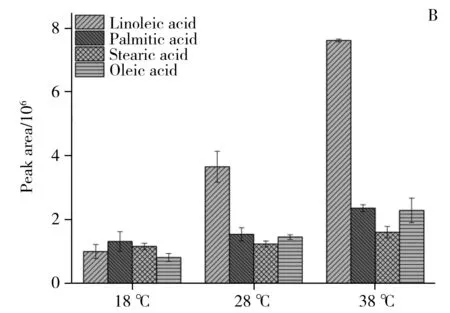
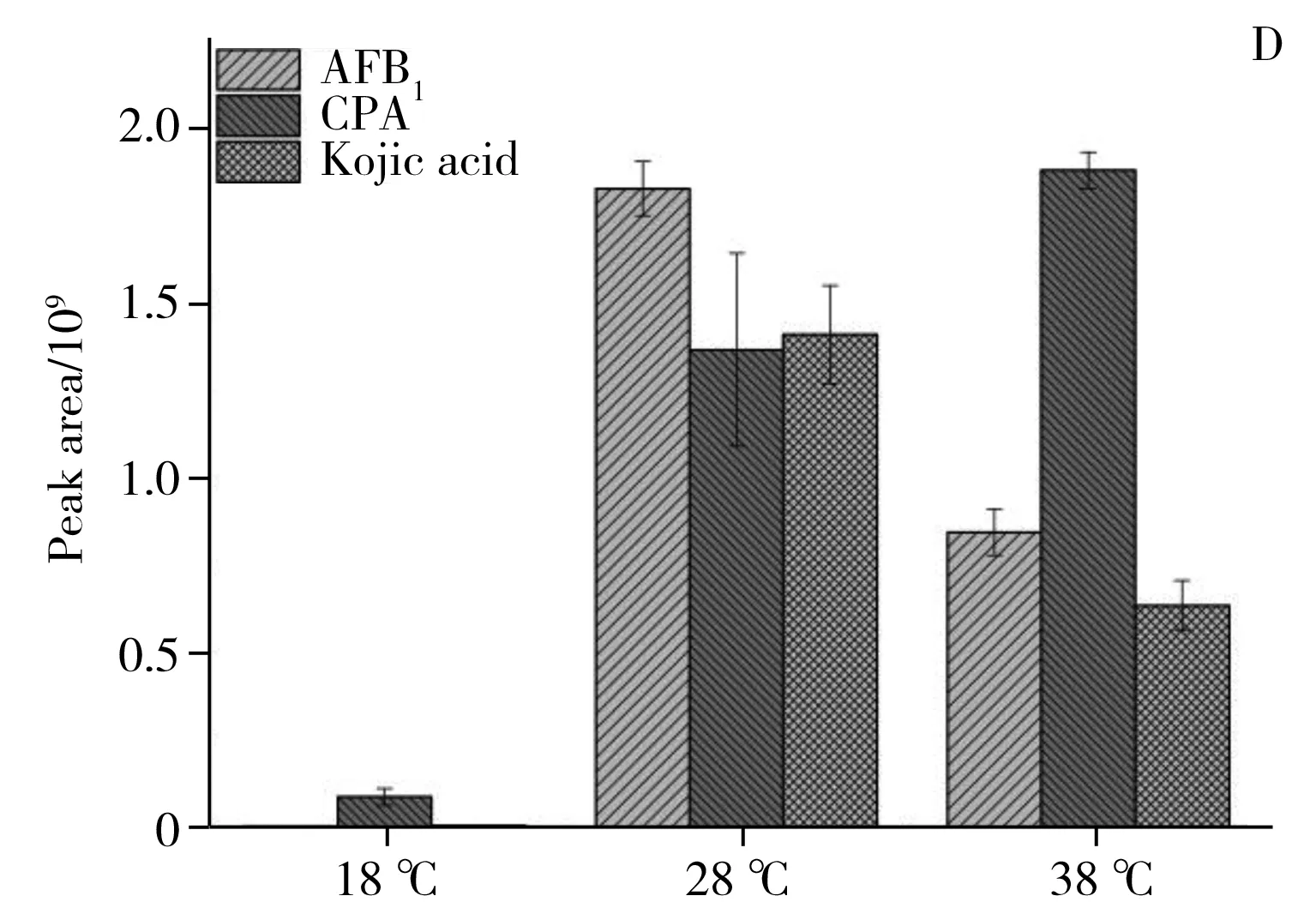
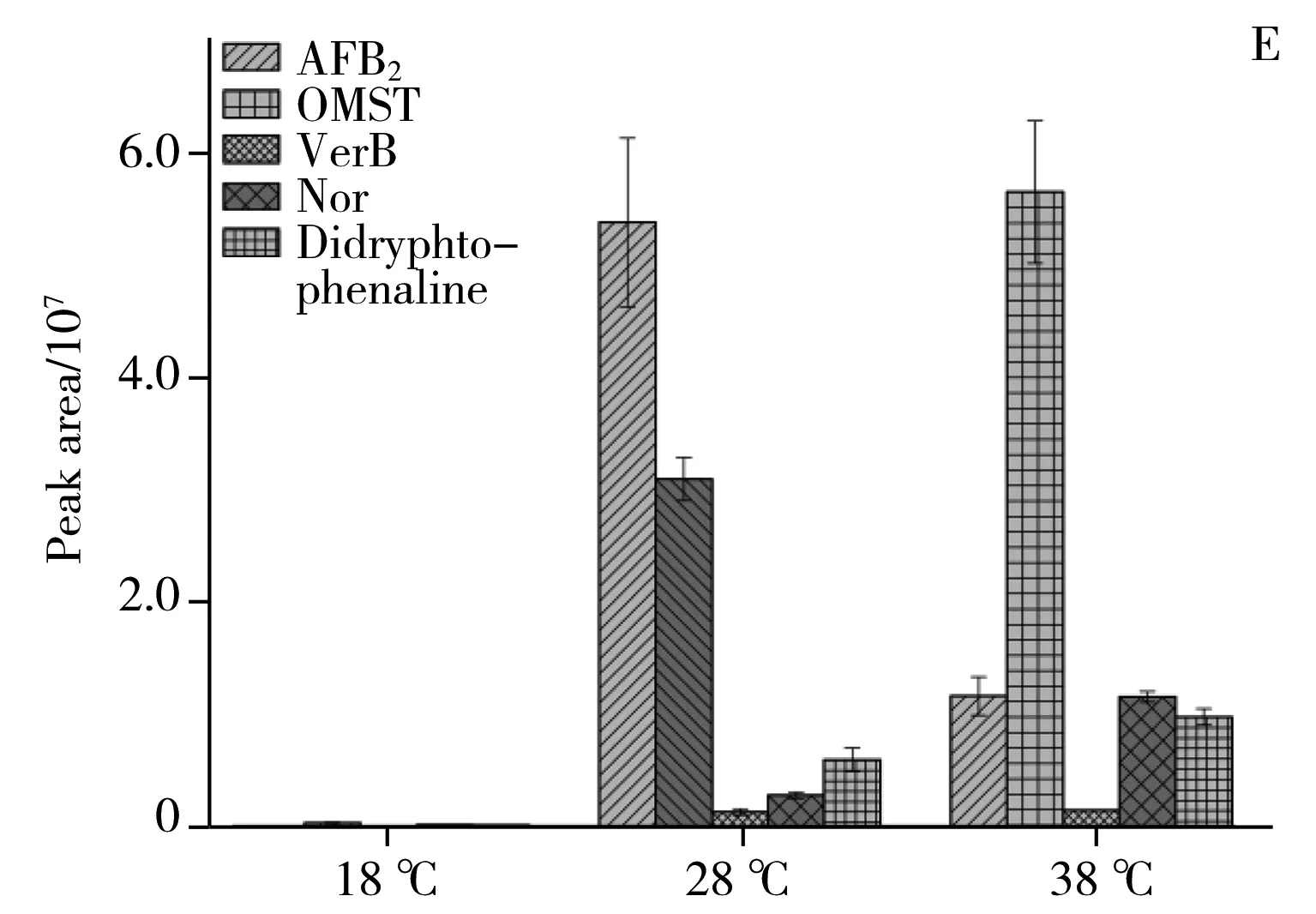
圖6 3個溫度條件下20個潛在差異代謝物的相對含量比較Fig.6 The relative content comparison of 20 potential differential metabolites under the 18,28,38 ℃
2.2.4潛在差異代謝物的代謝通路分析利用MBROLE 2.0[19]對鑒定的潛在差異代謝物進行代謝通路分析,結合表1和圖6A、B、C可看出,各潛在差異代謝物變化趨勢如下:18 ℃相較于28 ℃組,三羧酸循環中的蘋果酸、檸檬酸,脂肪酸生物合成中的軟脂酸(Palmitic acid)和硬脂酸(Stearic acid),油酸(Oleic acid)和亞油酸(Linoleic acid),纈氨酸(Valine),亮氨酸(Leucine),苯丙氨酸(Phenylalanine),色氨酸(Tryptophan),絡氨酸(Tyrosine)下調。其中色氨酸是必需氨基酸,在生物體中起著至關重要的作用,在能量代謝中是蛋白質合成或分解的中間體。這些化合物的下調說明低溫抑制了黃曲霉菌的能量代謝,同時也降低了對黃曲霉毒素合成[20]提供乙酰輔酶A等前體化合物反應原料供應。從圖6D、E可以發現,低溫顯著下調了黃曲霉毒素B1、B2、黃匹阿尼酸、曲酸、3a,3a′-二聚吡咯吲哚生物堿等次生代謝物。黃匹阿尼酸的生物合成[21]前體物質包括色氨酸,而色氨酸的下調導致黃匹阿尼酸的合成下調。此外,經過標品鑒定的黃曲霉毒素生物合成路徑上的前體物質降散盤衣酸、雜色曲菌素 B在低溫18 ℃下都實現了下調,但并未完成阻斷。推測原因如下:低溫(18 ℃)降低了黃曲霉毒素生物合成路徑上的合成酶活性;低溫(18 ℃)使黃曲霉菌三羧酸循環,脂肪酸合成,必須氨基酸等與能量代謝路徑有關的化合物下調,而能量代謝可產生乙酰輔酶A,該化合物才可進一步合成丙二酰輔酶A和黃曲霉毒素生物合成路徑上的首個穩定的前體物質降散盤衣酸。從圖6C可見,高溫38 ℃相比于28 ℃培養條件,黃曲霉初生代謝三羧酸循環中的蘋果酸、檸檬酸、琥珀酸相繼下調,但下調幅度較18 ℃和28 ℃組小。說明高溫稍微抑制了黃曲霉菌的呼吸代謝,降低了為黃曲霉毒素生物合成前體原料。從圖6A、B、C可見,纈氨酸、亮氨酸、苯丙氨酸、色氨酸、絡氨酸代謝,脂肪酸生物合成中的軟脂酸和硬脂酸,油酸和亞油酸,碳代謝中的山梨醇、木糖醇相繼上調。說明溫度的升高提升了黃曲霉菌碳代謝、脂肪生物合成等路徑中酶活性,溫度促進了這些代謝中的酶促反應,從而使黃曲霉生成更多的脂質并進行生物質積累,這與Elmer[22]等的研究結果相似。在次生代謝方面,從圖6D、E可見,升溫使黃匹阿尼酸出現上調,3a,3a′-二聚吡咯吲哚生物堿也出現上調,曲酸和黃曲霉毒素B1,B2在38 ℃出現顯著下調,但前體化合物降散盤衣酸、雜色曲菌素B、氧甲基雜色曲霉素卻上調,原因可能是:38 ℃條件下抑制了黃曲霉毒素合成路徑上從氧甲基雜色曲霉毒素轉化到黃曲霉毒素的酶的活性。特別地,溫度調控黃曲霉毒素和黃匹阿尼酸的生物合成的結果與Yu等[23]轉錄組研究結果相吻合,高溫使黃曲霉毒素的重要調控因子AflR和AflS出現下調,從而使黃曲霉毒素的合成量降低。總而言之,溫度調控了三羧酸循環、脂肪酸等初生代謝途徑,并影響黃曲霉毒素、黃匹阿尼酸、曲酸等次生代謝物生物合成路徑酶活性。這一研究結果從代謝水平上對影響黃曲霉菌生長和產毒的重要非生物因子——溫度如何調控黃曲霉菌生長和產毒素機制有了更深的理解。
3 結 論
本研究利用非靶標代謝組方法研究了18、28、38 ℃ 3個溫度條件對黃曲霉菌生理代謝機制的影響。共鑒定出20個溫度顯著調控黃曲霉菌生理代謝的潛在差異代謝物,發現溫度主要影響三羧酸循環、脂肪酸生物合成、糖代謝和氨基酸代謝等能量代謝通路以及黃曲霉毒素、曲酸、黃匹阿尼酸、3a,3a′-二聚吡咯吲哚生物堿等次生代謝通路。表明溫度通過影響上述代謝通路調控黃曲霉菌的生長和產生黃曲霉毒素、曲酸、黃匹阿尼酸等次生代謝物。這些差異代謝物為黃曲霉毒素風險評估研究提供了候選參考目標,為黃曲霉毒素分子預警提供了新的分析方法。
[1] Roze L V,Hong S Y,Linz J E.AnnualReviewofFoodScienceandTechnology,2013,4:293-311.
[2] Khaldi N,Seifuddin F T,Turner G,Haft D,Nierman W C,Wolfe K H,Fedorova N D.FungalGeneticsandBiology,2010,47(9):736-741.
[3] Chanda A,Roze L V,Kang S,Artymovich K A,Hicks G R,Raikhel N V,Calvo A M,Linz J E.ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica,2009,106(46):19533-19538.
[4] Bayram O,Krappmann S,Ni M,Bok J W,Helmstaedt K,Valerius O,Braus-Stromeyer S,Kwon N J,Keller N P,Yu J H,Braus G H.Science,2008,320(5882):1504-1506.
[5] Kong H W,Dai W D,Xu G W.Chin.J.Chromatogr.(孔宏偉,戴偉東,許國旺. 色譜),2014,32(10):1052-1057.
[6] Rubert J,Zachariasova M,Hajslova J.FoodAddit.Contam.,2015,32(10):1685-1708.
[7] Nielsen K F,Larsen T O.FrontiersinMicrobiology,2015,6:71.
[8] Arroyo-Manzanares N,Di Mavungu J D,Uka V,Malysheva S V,Cary J W,Ehrlich K C,Vanhaecke L,Bhatnagar D,De Saeger S.FoodAdditives&ContaminantsA,2015,32(10):1656.
[9] Smart K F,Aggio R B,Van Houtte J R,Villas-Boas S G.NatureProtocols,2010,5(10):1709-1729.
[10] Xia J G,Sinelnikov I V,Han B,Wishart D S.NucleicAcidsRes.,2015,43(W1):W251-W257.
[11] Tautenhahn R,Patti G J,Rinehart D,Siuzdak G.Anal.Chem.,2012,84(11):5035-5039.
[12] Godzien J,Alonso-Herranz V,Barbas C,Armitage E G.Metabolomics,2015,11(3):518-528.
[13] Naz S,Vallejo M,Garcia A,Barbas C.J.Chromatogr.A,2014,1353:99-105.
[14] Dunn W B,Wilson I D,Nicholls A W,Broadhurst D.Bioanalysis,2012,4(18):2249-2264.
[15] Dunn W B,Broadhurst D,Begley P,Zelena E,Francis-McIntyre S,Anderson N,Brown M,Knowles J D,Halsall A,Haselden J N,Nicholls A W,Wilson I D,Kell D B,Goodacre R.NatureProtocols,2011,6(7):1060-1083.
[16] Engskog M K R,Haglof J,Arvidsson T,Pettersson C.Metabolomics,2016,12(7):114.
[17] Zhang Y X,Yang X,Xie B J,She Y X,Du P F,Zou P,Jin F,Jin M J,Shao H,Wang S S,Zheng L F,Wang J.J.Instrum.Anal.(張艷欣,楊鑫,謝冰潔,佘永新,杜鵬飛,鄒攀,金芬,金茂俊,邵華,王珊珊,鄭鷺飛,王靜.分析測試學報),2016,35(5):501-507.
[18] Eriksson L.Multi-andMegavariateDataAnalysis.3nd ed.Byrne T,Johansson E,Trygg J,Vikstrom C.Sweden:Umetrics Inc,2013:422.
[19] Lopez-Ibanez J,Pazos F,Chagoyen M.NucleicAcidsRes.,2016,44(W1):W201-W204.
[20] Yu J J,Chang P K,Ehrlich K C,Cary J W,Bhatnagar D,Cleveland T E,Payne G A,Linz J E,Woloshuk C P,Bennett J W.Appl.Environ.Microbiol,2004,70(3):1253-1262.
[21] Seshime Y,Juvvadi P R,Tokuoka M,Koyama Y,Kitamoto K,Ebizuka Y,Fujii I.Bioorganic&MedicinalChem.Lett.,2009,19(12):3288-3292.
[22] Shih C N,Elmer H M.ZLebensmUnters.-Forsch.,1975,158:215-224.
[23] Yu J J,Fedorova N D,Montalbano B G,Bhatnagar D,Cleveland T E,Bennett J W,Nierman W C.FEMSMicrobiolLett.,2011,322(2):145-149.
Study on Effects of Temperature on Metabolism of Aspergillus Flavus Based on Untargeted Metabolomics
XIE Hua-li1,2,3,4,LI Pei-wu1,2,3,4,5*,WANG Xiu-pin1,3,4,5*,ZHANG Qi1,2,3,4,ZHANG Liang-xiao1,3,4,5,WANG Tong1,2,3,ZHANG Wen1,2,3,5,WANG Xue-fang1,2,3,5
(1.Oil Crops Research Institute of the Chinese Academy of Agricultural Science,Wuhan 430062,China;2.Key Laboratory of Biology and Genetic Improvement of Oil Crops,Wuhan 430062,China;3.Ministry of Agriculture,Key Laboratory of Detection for Mycotoxins,Wuhan 430062,China;4.Ministry of Agriculture,Laboratory of Risk Assessment for Oilseeds Products,Wuhan 430062,China;5.Ministry of Agriculture,Quality Inspection and Test Center for Oilseeds Products,Ministry of Agriculture,Wuhan 430062,China )
A method of non-targeted metabolomics coupled to ultra-high performance liquid chromatography- high resolution mass spectrometry was used to investigating effect of temperature on the metabolism of aspergillus flavus. Multivariate statistical analysis on the metabolomics data was carried out by using chemometric methods of OrthoPLS-DA(alternative partial least squares discriminant analysis) etc. The metabolism characteristics of aspergillus flavus were identified by MS/MS and library searching. In the study,the non-targeted metabolomics analytical method was validated by using internal standard and pooled quality control samples. The method was applied to investigate the effect of temperature on the metabolome of aspergillus flavus,and 3 593(T-test,p<0.01) differential metabolites were found under different temperature conditions. Twenty candidate biomarkers were screened out. The results showed that temperature significantly affected the biosynthetic pathways of tricarboxylic acid cycle,fatty acid,phenylalanine,tryptophan and tyrosine,and regulated the secondary metabolites biosynthetic pathway enzymesactivity of aflatoxin,cyclopiazonic acid and kojic acid. This study provide a new insight for risk assessment and early warning research of aflatoxin in China.
aspergillus flavus;metabolomics;high-resolution mass spectrum;aflatoxins biosynthesis;temperature
2017-02-08;
2017-08-17
國家自然科學基金(31640062);國家農產品質量安全風險評估重大項目;國家糧食行業科研專項(201513006)
*
李培武,研究員,研究方向:農產品質量與食物安全,Tel:027-86812943,E-mail:peiwuli@oilcrops.cn 王秀嬪,副研究員,研究方向:農產品質量安全檢測技術,E-mail:xiupinwang@163.com
10.3969/j.issn.1004-4957.2017.11.003
O657.63
A
1004-4957(2017)11-1304-09
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
