同一養殖環境下不同規格三疣梭子蟹競食能力的比較研究
2017-12-18 02:45:30史會來許文軍王庚申謝建軍
水產科學 2017年6期
何 杰,史會來,許文軍,王庚申,汪 瑋,施 慧,謝建軍
( 1.浙江省海洋水產研究所,浙江省海水增養殖重點實驗室,浙江 舟山 316021;2.浙江海洋大學,浙江 舟山 316021 )
同一養殖環境下不同規格三疣梭子蟹競食能力的比較研究
何 杰1,2,史會來1,許文軍2,王庚申1,汪 瑋1,2,施 慧1,謝建軍1
( 1.浙江省海洋水產研究所,浙江省海水增養殖重點實驗室,浙江 舟山 316021;2.浙江海洋大學,浙江 舟山 316021 )
將3種不同規格的三疣梭子蟹[平均體質量分別為(101.26±5.36) g、(62.37±4.10) g和(24.37±2.10) g]各10只飼養于2.6 m×1.8 m×1 m的室內水泥池單元格內,水深40 cm,重復3組。每日17:00投喂鮮活的帶魚塊,通過攝像機觀察不同規格三疣梭子蟹的競食行為,每組重復3次。結果表明:(1)不同規格三疣梭子蟹發現、抱持食物所需的時間相近,無顯著差異(P>0.05);(2)個體間競食激烈,(101.26±5.36) g組蟹搶食的比率最高,搶奪(62.37±4.10) g組或(24.37±2.10) g組蟹所持有的食物;(101.26±5.36) g組蟹搶食的次數顯著多于(62.37±4.10) g組或(24.37±2.10) g組蟹的搶食次數(P<0.05),(24.37±2.10) g組蟹搶食的次數最少;(24.37±2.10) g組蟹被(101.26±5.36) g、(62.37±4.10) g、(24.37±2.10) g組蟹搶食的次數及總次數最高,顯著多于(101.26±5.36) g組蟹(P<0.05),(62.37±4.10) g組蟹次之,(101.26±5.36) g組蟹最少,后兩組蟹無顯著差異(P>0.05),表現出明顯的“大欺小”現象;(3)(101.26±5.36) g組蟹的食物占有率(45%)顯著高于(24.37±2.10) g組蟹(近20%)(P<0.05),而(62.37±4.10) g組蟹占有的食物數量介于(101.26±5.36) g組和(24.37±2.10) g組之間,(101.26±5.36) g組蟹啃食完食物所用的時間僅約為(24.37±2.10)g組蟹的1/5倍。
三疣梭子蟹;規格;競爭;食物
當同種或異種動物對有限的公共資源有共同需要時就會發生種內或種間的競爭。競爭的主要目標包括食物、窩巢或配偶等,而對食物的競爭最為常見[1]。目前,國內外有關動物競爭食物的研究主要見于陸上昆蟲、鼠類以及部分大型哺乳動物[2-3],而有關水生動物的競食效應研究較少,且主要集中于自然水域條件下魚、蟹、蝦類的種間食物競爭關系研究等[4-5],有關人工養殖條件下的種內或種間食物競爭研究極為鮮見。
蝦蟹類甲殼動物依靠蛻殼(或蛻皮)實現體型的增大和體質量的增加,呈現出階梯式的生長模式。由于自身遺傳和社群控制的作用,即使在同一環境下,同一群體的蝦蟹類蛻殼也難以同步,致使其在養殖中后期規格分化現象極其嚴重[6-8]。然而,迄今為止,尚未見有關蝦蟹類規格分化(個體大小差異)對其種內競爭食物的行為能力研究。鑒于此,本文以生性好斗、領域性極強的海水養殖的三疣梭子蟹(Portunustrituberculatus)為對象,初步探討了在同一養殖環境下規格分化對競食能力的影響,旨在為豐富甲殼動物的行為生態學、種群生態學和養殖生態學資料,為三疣梭子蟹的科學養殖提供參考。
1 材料與方法
1.1 材料
本試驗于2016年8月在浙江省海洋水產研究所西閃島試驗場進行。在池塘養殖三疣梭子蟹去雄期間,從池塘內誘釣、挑選十足健全的雄梭子蟹200余只運回實驗室,暫養于8 m×1.8 m×1 m的室內水泥池,水深40 cm,連續充氧,每日17:00投喂鮮活帶魚(Trichiuruslepturus)塊,次日8:00清除殘餌,換水1/3。暫養3 d后,從中挑選相對較大規格(101.26±5.36) g、中等規格(62.37±4.10) g和較小規格(24.37±2.10) g的蟹各30只用于正式試驗。
1.2 方法
選1口8 m×1.8 m×1 m的室內水泥池,用漁網、磚塊等材料等分成3個2.6 m×1.8 m×1 m的單元格,單元格間的水流相通,水深40 cm,連續充氧。在各單元格上空中心位置安裝一不銹鋼支架,用于固定攝像機(索尼HDR-PJ410)。之后向各單元格內放養(101.26±5.36) g組、(62.37±4.10) g組、(24.37±2.10) g組三疣梭子蟹各10只,每日17:00投喂鮮活的帶魚塊,次日8:00清除殘餌,換水1/2,試驗期間的水溫(28±2) ℃,pH 8.8±0.1,溶解氧>5 mg/L。4 d后,待蟹基本適應試驗環境后開始正式試驗。試驗期間,調準好攝像機焦距后,向單元格中間區域投喂10塊(份)大小相近(1.52±0.13) g的帶魚塊,待所有蟹啃食完畢后關閉攝像機(15~30 min),按同樣方法每日依次記錄3個單元格內蟹的攝食和競食行為,持續3 d,即整個試驗重復9次。拍攝結束后,使用聯想G425筆記本電腦讀取攝像機SD卡內的影像資料,統計每次攝食期間蟹索食所用時間、魚塊被不同蟹的占有率、啃完食物所用時間,并從每個重復中隨機挑選6~8只蟹,分別記錄每只蟹在整個啃食過程中被同類搶食的次數、是否被成功搶走以及其攜帶食物更換空間位置(以水平位移大于20 cm為標準)的次數等。
1.3 數據處理
試驗數據用平均值±標準差表示,不同試驗組數據間采用單因素方差分析,對檢測達到顯著水平的均值用Duncan檢驗,用SPSS 11.5進行方差分析和多重比較,在Sigmaplot 10.0下繪制相關圖表。
2 結 果
2.1 不同規格三疣梭子蟹索食所需的時間
投喂食物后,蟹反應積極,索食行為明顯,一旦發現食物,用螯足迅速抱持食物,送入口中,第一大顎不斷切碎和磨損食物,第一和第二對顎足護住食物,以防流失。由圖1可見,不同規格三疣梭子蟹發現并抱持食物所需的時間均約為0.28 min,差異不顯著(P>0.05)。
2.2 不同規格三疣梭子蟹搶食的行為特征
試驗期間投喂的餌料不足,同類之間發生了激烈的爭奪食物行為。其行為可分為5個階段:(1)相遇階段,雙方身體姿態不變,沒有額外使用附肢進攻或防御(圖2a);(2)示威階段,搶食者向食物持有者靠近,抬高身體、展開大螯向對方示威,食物持有者用其中一個螯足攜持食物,高舉另一螯足防御,并試圖側身而逃(圖2b,c);(3)搶奪階段,搶食者直接用螯足搶奪對方所持有的食物,雙方均用螯足夾住食物,互不相讓(圖2d);(4)拳(螯)擊階段,雙方不斷使用大螯推擠,并努力翻轉或扭曲對方的螯足,企圖損傷或卸去對方的螯足(圖2e,f),雙方打斗極為激烈,水波的干擾致拍攝的照片較為模糊;(5)撤退階段,食物完全被對方搶走后,失敗方撤離,搶奪結束(圖2g,h)。

圖1 不同規格三疣梭子蟹索食所需的時間
40%~80%持有食物的試驗蟹曾被(101.26±5.36) g組蟹搶奪,35%~60%的個體曾被(62.37±4.10) g組蟹搶奪,20%~50%的個體曾被(24.37±2.10) g組蟹搶奪,被(101.26±5.36) g組蟹搶食的比率最高,被(62.37±4.10) g組蟹搶食的比率次之,被(24.37±2.10) g組蟹搶食的比率最低,說明大規格蟹表現出最明顯的主動搶食行為;另外,(101.26±5.36) g組蟹更傾向于搶奪(62.37±4.10) g或(24.37±2.10) g組蟹所持有的食物,而(24.37±2.10) g組蟹更多地選擇向與其規格相近的(24.37±2.10) g組蟹發起搶食攻擊,但其中也有部分個體向(101.26±5.36) g和(62.37±4.10) g組蟹搶奪食物,其比率顯著小于向(24.37±2.10) g組蟹搶食的比率(P<0.05)(圖3)。各組蟹被同類搶食的次數相似(圖4),即(101.26±5.36) g、(62.37±4.10) g、(24.37±2.10) g組蟹均被(101.26±5.36) g組蟹搶食的次數最多,被(24.37±2.10) g組蟹搶食的次數最少,其中(101.26±5.36) g和(24.37±2.10) g組蟹被(101.26±5.36) g組蟹搶食的次數顯著多于被(62.37±4.10) g或(24.37±2.10) g組蟹搶食的次數(P<0.05);而各組蟹之間相比,(24.37±2.10) g組蟹分別被(101.26±5.36) g、(62.37±4.10) g、(24.37±2.10) g組蟹搶食的次數及總次數均為最多,(62.37±4.10) g組蟹次之,(101.26±5.36) g組蟹最少,且(24.37±2.10) g組蟹被同類搶食的次數顯著多于(101.26±5.36) g組蟹(P<0.05),(62.37±4.10) g組蟹與(101.26±5.36) g組和(24.37±2.10) g組蟹均無顯著差異(P>0.05)。
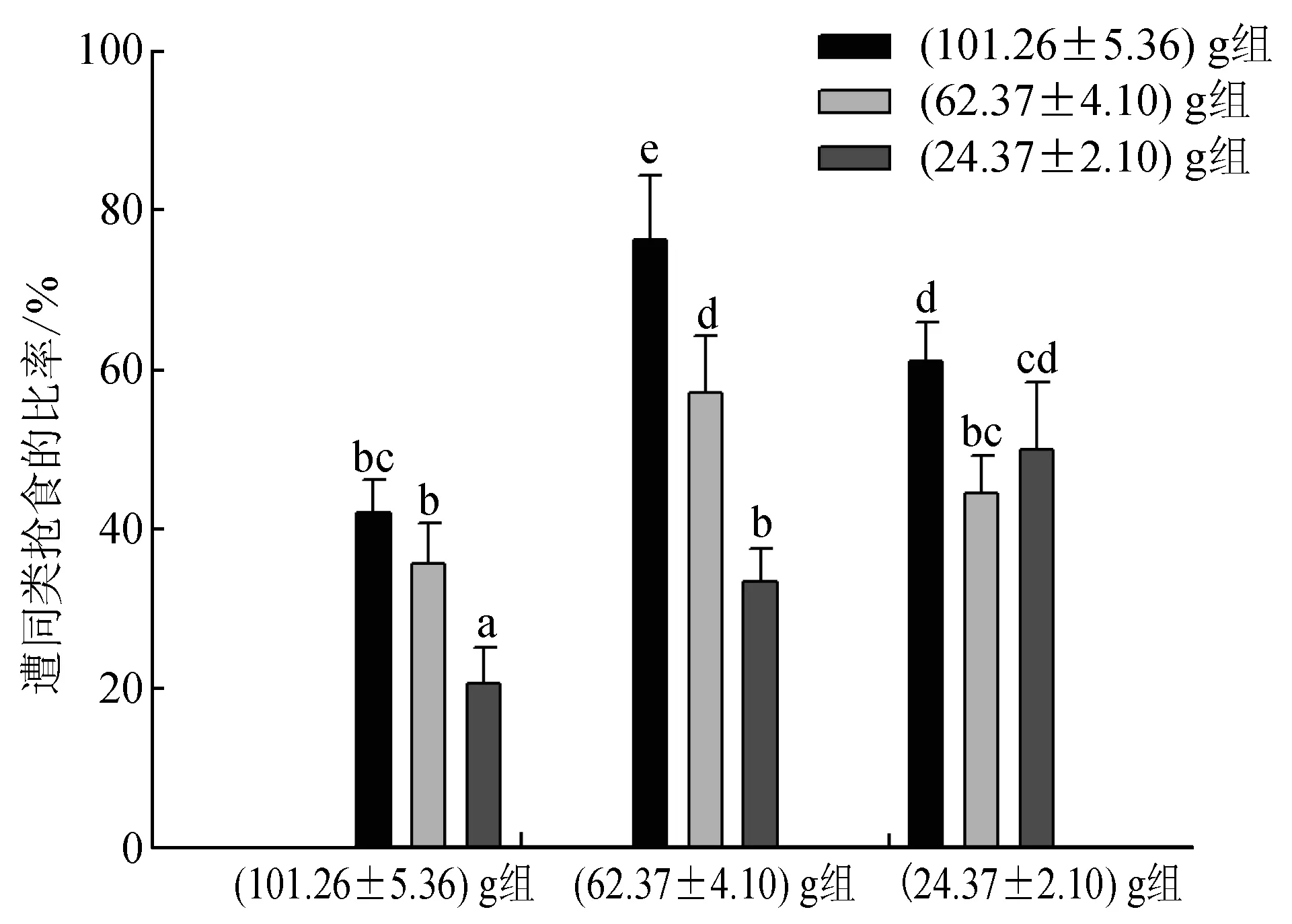
圖3 不同規格三疣梭子蟹遭同類搶食的比率同一系列上標有相同字母表示差異不顯著(P>0.05),標有相異字母表示差異顯著(P<0.05),下同.
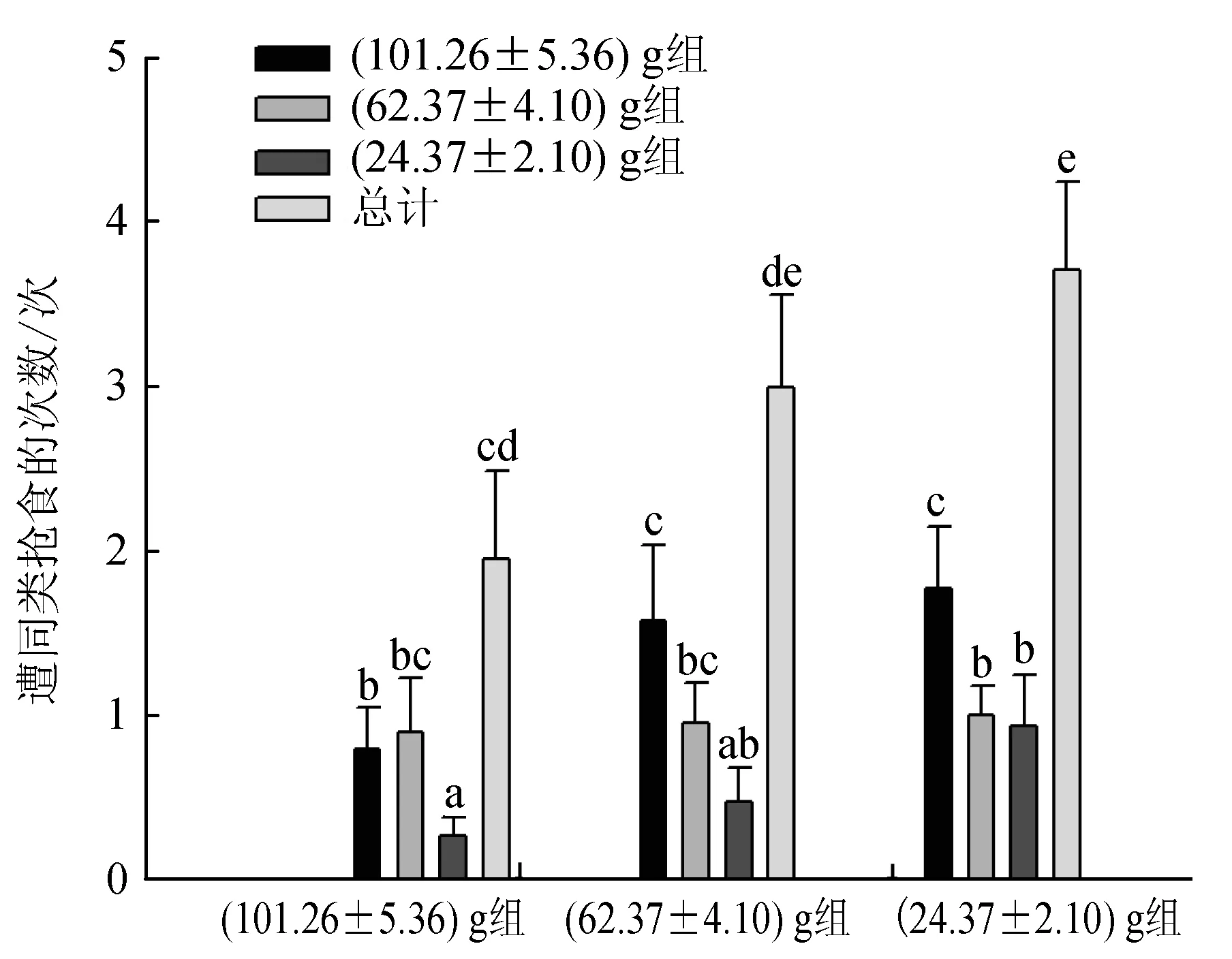
圖4 不同規格三疣梭子蟹遭同類搶食的次數
三疣梭子蟹在啃食期間不停更換空間位置來降低食物被同類發現或被搶奪的概率,其中(24.37±2.10) g組蟹最為明顯,在整個啃食過程中更換了約10次空間位置,顯著高于(62.37±4.10) g組(約6次)和(101.26±5.36) g組(約4次)蟹(P<0.05)(圖5)。無論是(101.26±5.36) g、(62.37±4.10) g還是(24.37±2.10) g組,三疣梭子蟹所持有的食物均可能被同類搶走。(24.37±2.10) g組蟹中50%的個體所持有的食物被同類搶走,顯著高于(62.37±4.10) g組(約30%)和(101.26±5.36) g組(約10%)的蟹(P<0.05)(圖6)。而且(24.37±2.10) g組蟹所持食物均可能被任何蟹成功搶奪,被(101.26±5.36) g組蟹搶走的比率最高,被(24.37±2.10) g組蟹搶走的比率最低,而(101.26±5.36) g組的蟹僅能被與其規格相近的(101.26±5.36) g組蟹搶走,不能被比其規格小的(62.37±4.10) g組和(24.37±2.10) g組蟹搶走,同樣,(62.37±4.10) g組蟹所持有的食物僅能被(101.26±5.36) g組和(62.37±4.10) g組的蟹搶走,不可能被(24.37±2.10) g組的蟹搶走。
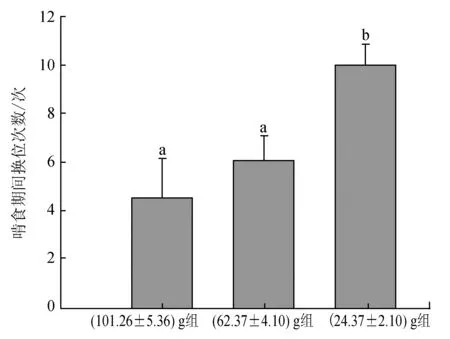
圖5 不同規格三疣梭子蟹在啃食期間身體移動次數
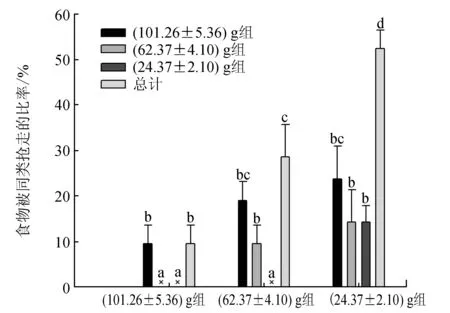
圖6 不同規格三疣梭子蟹所持食物被同類搶走的比率
2.3 不同規格三疣梭子蟹對有限食物的占有率及其啃食所需時間
不同規格三疣梭子蟹對有限食物的占有率不同,其中(101.26±5.36) g組三疣梭子蟹占有率最高,侵占了約45%的食物,顯著高于(24.37±2.10) g組蟹(近20%)(P<0.05),而(62.37±4.10) g組蟹占有的食物數量介于(101.26±5.36) g和(24.37±2.10) g之間(圖7)。啃食完食物所用的時間越短,啃食速度越快,花費時間越短,小規格蟹啃食所需時間約為大規格蟹的5倍(圖8)。

圖7 不同規格三疣梭子蟹對食物的占有率

圖8 不同規格三疣梭子蟹啃食食物所需的時間
3 討 論
動物種內競爭主要表現在同種對食物、生存空間、配偶等的爭奪與保衛[1]。水生動物的種內競爭強度由各種錯綜復雜的因素綜合引起,棲息環境惡劣、個體間大小懸殊、餌料不足以及種群擁擠等約能加劇種內競爭行為[9-10],在高密度、封閉的人工飼養環境下,養殖動物在獲取有限的生物資源時更易發生激烈的競爭[11]。養殖動物通過攝取食物來獲得能量,在餌料投喂不充足條件下,動物種內的食物競爭直接關系到個體的生存和生長,影響養殖效益,因此,探討養殖條件下水生經濟動物種內食物競爭行為的發生機制以及調節措施,對養殖對象的穩產和高產具有直接的指導意義。
蝦蟹類理想的養殖效果可概括為“兩高一短”,即“成活率高,蛻殼同步率高,蛻殼周期短”[12]。Segal等[13]研究發現,單個個體隔離養殖的羅氏沼蝦(Macrobrachiumrosenbergii)的蛻殼同步率明顯高于群體混養模式,認為直接的食物競爭是導致部分個體生長快速,部分個體被抑制生長的主要原因;董鵬生等[8]發現,即使是同一家系的中華絨螯蟹(Eriocheirsinensis)在同一生境下群養后,蛻殼仍然非常不同步,隨著養殖時間的延長,規格差異日益顯現,收獲時的大、小規格個體的體質量差異可達10倍以上,甚至部分個體生長過快出現早熟現象。可見,除甲殼動物自身遺傳背景外,社群控制是導致甲殼動物規格嚴重分化的主要誘因之一,而且種內競爭(主要指食物和生存空間)是社群控制的主要表現形式。
本研究中,不同或同一規格間的三疣梭子蟹均表現出激烈的種內競食行為,其行為大致可分為5個階段:相遇、示威、搶奪、拳擊和撤退階段,整個競食過程與先前報道的小龍蝦的爭勝行為相似[14]。筆者認為,這是甲殼動物個體間格斗、競爭資源時通用的行為模式。不同規格三疣梭子蟹被同類搶食的比率和次數表明,大規格個體的搶食欲望強于小規格個體,各規格蟹均趨向搶食比其自身規格小的個體,表現出明顯的“大欺小”的行為特征,這與羅氏沼蝦的種內競爭研究結果一致[15]。而小規格蟹只能通過不斷的更換空間位置,盡量回避競爭者的搶食行為。這是動物在應對危險環境下做出的一種規避性本能反應,也是本研究中小規格蟹啃食完相同大小食物所需時間遠大于大規格蟹的原因之一。本研究還發現,小規格三疣梭子蟹也能向比其自身大的個體進行攻擊性的搶奪食物,但比率較低,其更多的選擇與其規格相近的小規格個體搶食。從食物被同類搶走的比率以及各規格蟹對食物的最終占有率來看,大規格個體的搶食能力也明顯強于小規格個體,致使其獲得更多的食物資源,所以,在食物不充足的情況下,小規格三疣梭子蟹面臨饑餓的可能性更大。眾多研究已表明,饑餓脅迫不僅直接抑制甲殼動物成活和蛻殼成功率,還不同程度地影響蛻殼后的質量增加率和蛻殼周期,加劇了養殖群體的規格分化[16-17],種內食物競爭越趨激烈,以此惡性循環的發展,最終導致養殖動物成活率和養殖產量低下[6,18]。
規格分化將加劇甲殼動物種內競爭食物行為的發生,使部分個體生長攝入營養過多,生長過快而早熟,部分個體攝入營養不足,生長滯后甚至停止生長。因此,要不斷改進養殖設施和技術,降低種群內規格分化的程度,減少種內對食物、生存空間的競爭。近年來,三疣梭子蟹單個體養殖技術應運而生。該技術可大大提高三疣梭子蟹的成活率,但由于投餌、去殘餌的自動化管理技術配套不夠等,沒有得到大范圍的推廣,所以,今后的研究應加強三疣梭子蟹單個體養殖配套技術的開發。針對現有的池塘群養技術,應通過合理的放養密度、均勻投餌等措施,盡可能減少種內對食物的競爭,提高養殖成活率和規格整齊度。
[1] 尚玉昌. 普通生態學[M]. 北京:北京大學出版社, 2010:173-174.
[2] 楊效文. 麥蚜種間競爭研究初報 [J]. 生態學雜志, 1991,10(1):1-5.
[3] Farjana T, Tuno N, Higa Y, et al. Effects of temperature and diet on development and interspecies competition inAedesaegyptiandAedesalbopictus[J]. Medical and Veterinary Entomology, 2012, 26(2):210-217.
[4] 張波, 唐啟升, 金顯仕, 等. 東海和黃海主要魚類的食物競爭[J]. 動物學報, 2005, 51(4):616-623.
[5] 郭斌, 張波, 戴芳群, 等. 海州灣小黃魚幼魚和黃鯽幼魚的食物競爭[J]. 漁業科學進展, 2011, 32(1):8-15.
[6] Gebauer P, Paschke K, Anger K. Delayed metamorphosis in decapod crustaceans:evidence and consequences[J]. Revista Chilena de Historia Natural,2003,76(2):169-175.
[7] He J, Gao Y, Xie J J, et al. Limb autotomy patterns in the juvenile swimming crab (Portunustrituberculatus) in earth ponds[J]. Aquaculture,2016,463(3/4):189-192.
[8] 董鵬生, 劉青,吳旭干, 等.中華絨螯蟹一齡性早熟和二齡成熟家系扣蟹階段生長和早熟的比較研究[J].上海海洋大學學報,2016,25(1):51-60.
[9] Thomas C W, Carter C G, Crea B J. Feed availability and its relationship to survival, growth, dominance and the agonistic behaviour of the southern rock lobster,Jasusedwardsiiin captivity[J]. Aquaculture,2003, 215(1/4):45-65.
[10] 陳學雷, 林瓊武, 李少菁, 等.日本對蝦仔蝦相殘的實驗研究[J]. 廈門大學學報:自然科學版,2003,42(3):358-361.
[11] 李玉全, 孫霞. 水生動物的爭勝行為[J]. 動物學研究, 2013,34(3):214-220.
[12] Figueiredo J, Lopes G P, Narciso L, et al. Effect of starvation during late megalopa stage ofMithraculusforceps(Brachyura:Majidae) on larval duration, synchronism of metamorphosis, survival to juvenile, and newly metamorphosed juvenile size[J]. Aquaculture, 2008,274(1):175-180.
[13] Segal E, Roe A. Growth and behaviour of post-juvenileMacrobrachiumrosenbergii(de Man) in close confinement[J].Proceedings of the World Mariculture Society, 1975, 6(1/4):67-88.
[14] Moore P A. Agonistic behavior in freshwater crayfish——the influence of intrinsic and extrinsic factors on aggressive encounters and dominance[G]//Duffy J E, Thiel M. Evolutionary Ecology of Social and Sexual Systems-Crustaceans as Model Organisms, New York:Harvard University Press, 2007:90-114.
[15] Karplus I. Social control of growth inMacrobrachiumrosenbergii(De Man):a review and prospects for future research[J]. Aquaculture Research, 2005, 36(3):238-254.
[16] 吳立新, 董雙林, 姜志強.饑餓對甲殼動物生理生態學影響的研究進展[J].應用生態學報, 2004, 15(4):723-727.
[17] 何杰,王春,邵路暢,等.饑餓脅迫對中華絨螯蟹(Eriocheirsinensis)仔蟹的影響[J].生態學雜志, 2013,32(8):2077-2084.
[18] Polis G A. The evolution and dynamics of intraspecific predation[J].Annual Review of Ecology and Systematics,1981,12(12):225-251.
ComparisonofCompetitionforFoodamongSwimmingCrabPortunustritubercatuswithVariousSizesinSameHabitat
HE Jie1,2, SHI Huilai1, XU Wenjun2, WANG Gengshen1, WANG Wei1,2, SHI Hui1, XIE Jianjun1
( 1. Key Laboratory of Marine Culture and Enhancement of Zhejiang Province, Marine and Fisheries Research Institute of Zhejiang Province, Zhoushan 316021, China; 2. Zhejiang Ocean University, Zhoushan 316021, China )
A total of 30 swimming crabPortunustrituberculatuswith three different sizes [average body weight of (101.26±5.36) g (LC), (62.37±4.10) g (MC) and (24.37±2.10) g (SC)] were raised in the same indoor tanks of each 2.6 m×1.8 m×1 m with water depth of 40 cm with triplication. After crabs were fed fresh ribbon fish at 17:00 daily, camera was used to record their eating and competing behavior for food with triplication. The results showed that swimming crab with different body weight had similar foraging time for food (P>0.05). Intraspecific competition for food among swimming crab individuals was highly fierce and the behaviors were approximately divided into five stages:meet stage, demonstration stage, snatch stage, pugilism stage, and withdraw stage. The proportion and frequency of competition for food exhibited that the larger-sizes individuals had a stronger competition desire and were inclined to compete for food with smaller individuals. As for the proportion of being deprived of food and the ultimate occupancy of food, smaller-size individuals had relatively poorer capability of invasion and protection for food.
Portunustritubercatus; size;competition;food
10.16378/j.cnki.1003-1111.2017.06.008
S968.25
A
1003-1111(2017)06-0741-06
2016-10-10;
2017-01-16.
浙江省科技廳院所專項(2016F50042, 2015F10001, 2015F30003);浙江海洋大學博士啟動基金資助項目(22135010815 );舟山市科技計劃項目(2017C43032);舟山市普陀區科技計劃項目(2015YN209).
何杰(1985-),男,博士;研究方向:蝦蟹類健康養殖.E-mail:he_0902@126.com. 通訊作者:許文軍(1971-),男,教授;研究方向:海水健康養殖,E-mail:wjxu1971g@hotmail.com.
