不同群體中華鱉的形態多樣性分析
2017-12-18 02:49:14陸文浩韓曉磊徐建榮韓曜平
水產科學 2017年6期
關鍵詞:差異
陸文浩,韓曉磊,陳 梁,徐建榮,韓曜平
( 1. 太倉市水產指導站,江蘇 太倉 215400; 2. 常熟理工學院 生物與食品工程學院,江蘇 常熟 215500 )
不同群體中華鱉的形態多樣性分析
陸文浩1,韓曉磊2,陳 梁2,徐建榮2,韓曜平2
( 1. 太倉市水產指導站,江蘇 太倉 215400; 2. 常熟理工學院 生物與食品工程學院,江蘇 常熟 215500 )
對長江口、太湖和洪澤湖3個不同區域的中華鱉野生群體的16項生物形態特征進行測量,利用多元統計分析方法對其進行形態多樣性差異的比較研究。聚類結果顯示,長江口群體和太湖群體首先聚類,再與洪澤湖群體聚合;主成分分析中,共得到了3個主成分,其貢獻率分別為56.208%、10.894%和7.165%,累積貢獻率為74.267%,其種間形態差異主要取決于頭部的一些形態特征;判別分析中,洪澤湖群體、太湖群體和長江口群體的判別準確率分別為100%、100%和93.3%,平均為97.8%。試驗結果表明,在3個中華鱉群體中,長江口群體與洪澤湖群體之間分類水平已經達到亞種以上,而長江口群體和太湖群體還未達到亞種水平,說明采用形態差異多元分析進行中華鱉種間遺傳多樣性研究的可行性,同時也為種質保護和遺傳育種提供了一定的理論基礎。
中華鱉;多元統計分析;形態多樣性
中華鱉(Trionyxsinensis)是中國重要的特種經濟水生動物之一,隸屬于爬行綱、龜鱉目、鱉科、鱉屬,俗稱團魚、甲魚等。在我國,中華鱉分布廣泛,除西藏和青海外的其他各省均有發現,長江流域和華南地區分布較多[1]。中華鱉自古以來就是名貴的食材和藥物,具有較高的食用和藥用價值,因此在近代中華鱉的人工養殖發展非常迅速[2]。中華鱉養殖產業的高速發展也帶來了一系列問題的出現,由于中華鱉種間分類不明確,加之養殖企業引種頻繁,導致了種群混雜,群體多樣性不斷降低,致使中華鱉種質退化問題尤為突出[3]。目前研究顯示,中華鱉還沒有明確的亞種分類,但存在著一些不同的地理群體,而外部形態特征是中華鱉種質資源鑒定最簡單最直觀的重要方法[4],可以為中華鱉不同群體間的種質鑒定提供一定的理論支持。
多元統計分析能夠針對形態特征的多個參數,在多個對象和指標互相關聯的情況下分析其存在的統計規律,適合現階段農業科學研究的特點。目前,已大量應用于魚類物種確認、種群變異、性別差異等方面的研究[4-7],但是龜鱉類動物的相關報道還不多見。因此,本研究通過多元統計分析方法對中華鱉的形態參數進行研究,以期為中華鱉種質資源的管理、保護和開發利用提供有效的科學依據。
1 材料與方法
1.1 試驗材料
于洪澤湖流域、長江口流域和太湖流域3個不同地區天然水域進行野生中華鱉的樣品采集。每個群體隨機選取20~30尾中華鱉成鱉進行試驗,樣本信息見表1。

表1 中華鱉3個不同群體的樣本信息
1.2 試驗器材和試劑
游標卡尺(精度0.02 mm)、電子天平(精度0.001 kg)、塑料密閉容器(40 cm×25 cm×10 cm);磺酸間氨基苯甲酸乙酯。
1.3 試驗方法
由于活動的中華鱉難以對其進行測量,因此先將待測中華鱉放入一個密閉容器,滴入適量的磺酸間氨基苯甲酸乙酯將其麻醉后,再進行測量。測量包括中華鱉3個不同群體的16項形態指標,有體質量、體高、背甲周長、背甲長、背甲寬、腹甲長、腹甲寬、腹甲凹寬、后側裙邊寬、前肢長、后肢長、尾長、頭長、頭寬、眼間距、鼻寬。長度測量采用游標卡尺(精度為0.02 mm),體質量測量采用電子天平(精度為0.001 kg),測量方法參考國家標準(GB 21044—2007)[8]。
1.4 數據處理
試驗數據采用SPSS 19.1軟件進行處理,包括聚類分析、主成分分析、判別分析和差異系數檢測4種多元分析方法。為消除中華鱉規格大小對參數數值的影響,多元分析中均采用以背甲長為基數進行分析。本研究選取15項形態比例參數(表2),分別求出各群體形態比例參數的平均值,再用這些平均值進行相應的數據分析。

表2 形態特征比例參數
1.4.1 聚類分析
根據測量數據分別求出3個不同群體15項形態比例參數的平均值,之后通過平均值做出聚類分析。聚類方法為歐氏距離的最短距離系統聚類法[9]。
1.4.2 主成分分析
為了使多個形態指標適合小數指標的統計方法,先校正處理得出15個形態參數比例性狀,通過SPSS 19.1的主成分分析法得出3個互不相關的主成分,以及累計貢獻率和主成分貢獻率等[9]。
1.4.3 判別分析
在判別分析中,通過逐步判別法進行判別,其中判別準確率的計算參照文獻[10]的方法。
1.4.4 差異系數檢驗
形態差異的差異系數(CD)公式:
CD=(M1-M2)/(S1+S2)
式中,M1和M2分別表示兩個不同群體某項性狀指標的平均值;S1和S2是標準差。如差異系數數值大于1.28,則可視為亞種及以上水平的差異;反之,則認為差異未達到亞種水平[11]。
2 結 果
2.1 聚類分析
根據形態數據分析,洪澤湖、長江口和太湖3個不同群體形態比例參數的聚類見圖1。由圖1可見,3個群體分為兩支,其中長江口群體和太湖群體首先聚類,之后與洪澤湖群體聚合。太湖群體和長江口群體的形態相似度較高,而與洪澤湖群體的形態相似度則相對較低。

圖1 中華鱉3個不同群體的形態聚類
2.2 主成分分析
2.2.1 3個群體主成分
選用方差貢獻率較大的3個主成分,其貢獻率分別是主成分1為56.208%、主成分2為10.894%、主成分3為7.165%,累積貢獻率為74.267%。3個主成分累積貢獻率較高,包含其總變異的大部分,可以使用此3個相互獨立的因子來描述中華鱉3個群體間的形態差異。在主成分1中,x1、x6、x12、x13和x14這5個變量影響較大;在主成分2中,x3和x5兩個變量影響較大;在主成分3中變量x8影響較大(表3)。對總變差貢獻率最大的主成分1進行深入分析,發現其主要取決于背甲周長、眼間距、尾長、頭寬、頭長和鼻寬這幾個形態指標,即主成分1基本上反映的是頭部形態的特征,因此可以認為,洪澤湖、太湖、長江口3個群體在形態上的差異很大程度上是由頭部的長寬不同、眼睛和鼻子的形態差別引起的。
2.2.2 群體散布圖分析
3個流域中華鱉群體的主成分1和主成分2的散點圖見圖2,其中主成分1和主成分2為相對值。由圖2可見,洪澤湖的中華鱉與其他兩個群體間不存在重疊部分,而太湖和長江口之間有一小部分重疊,運用主成分1和主成分2就能將3個群體基本分開。由此可見,洪澤湖、太湖和長江口3個中華鱉群體在形態上差別較大,通過形態特征即可將其基本分開。
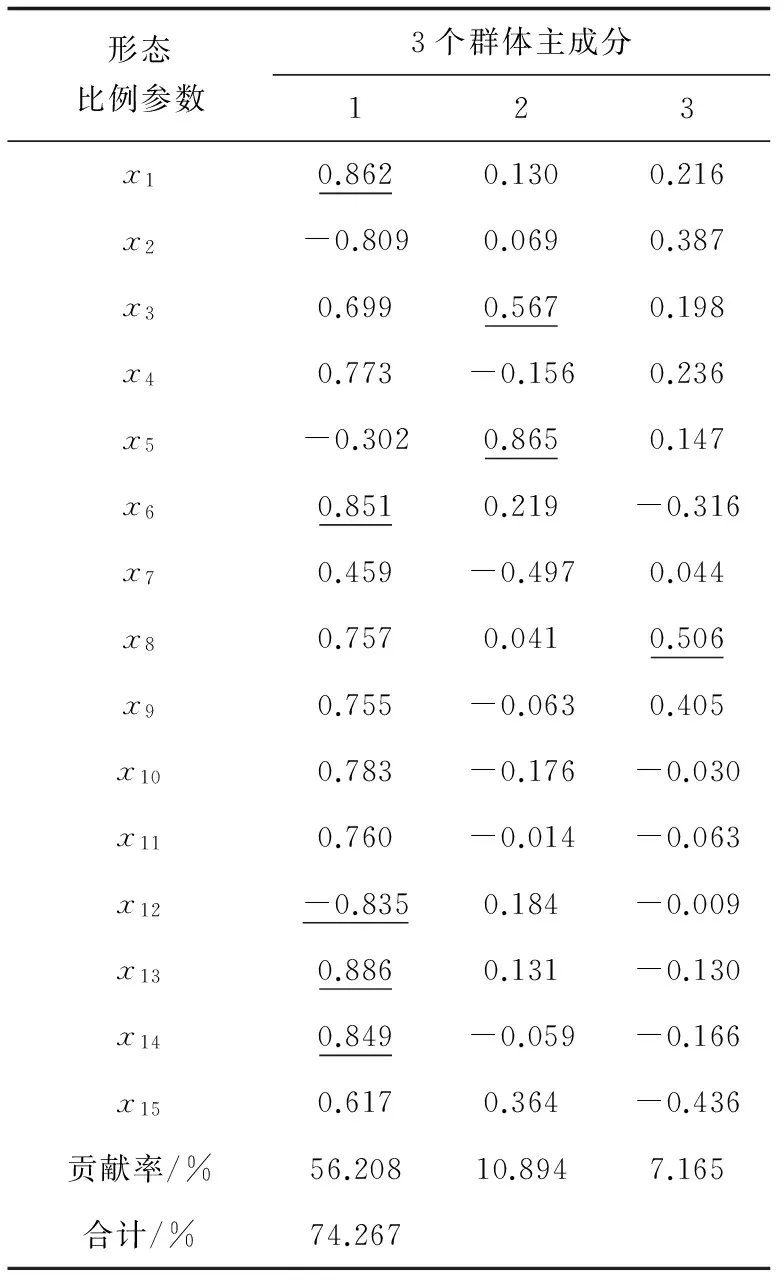
表3 3個群體形態特征因子負荷矩陣及主成分貢獻率
注:下劃線表示負荷值大于0.500000,并作為主因子.
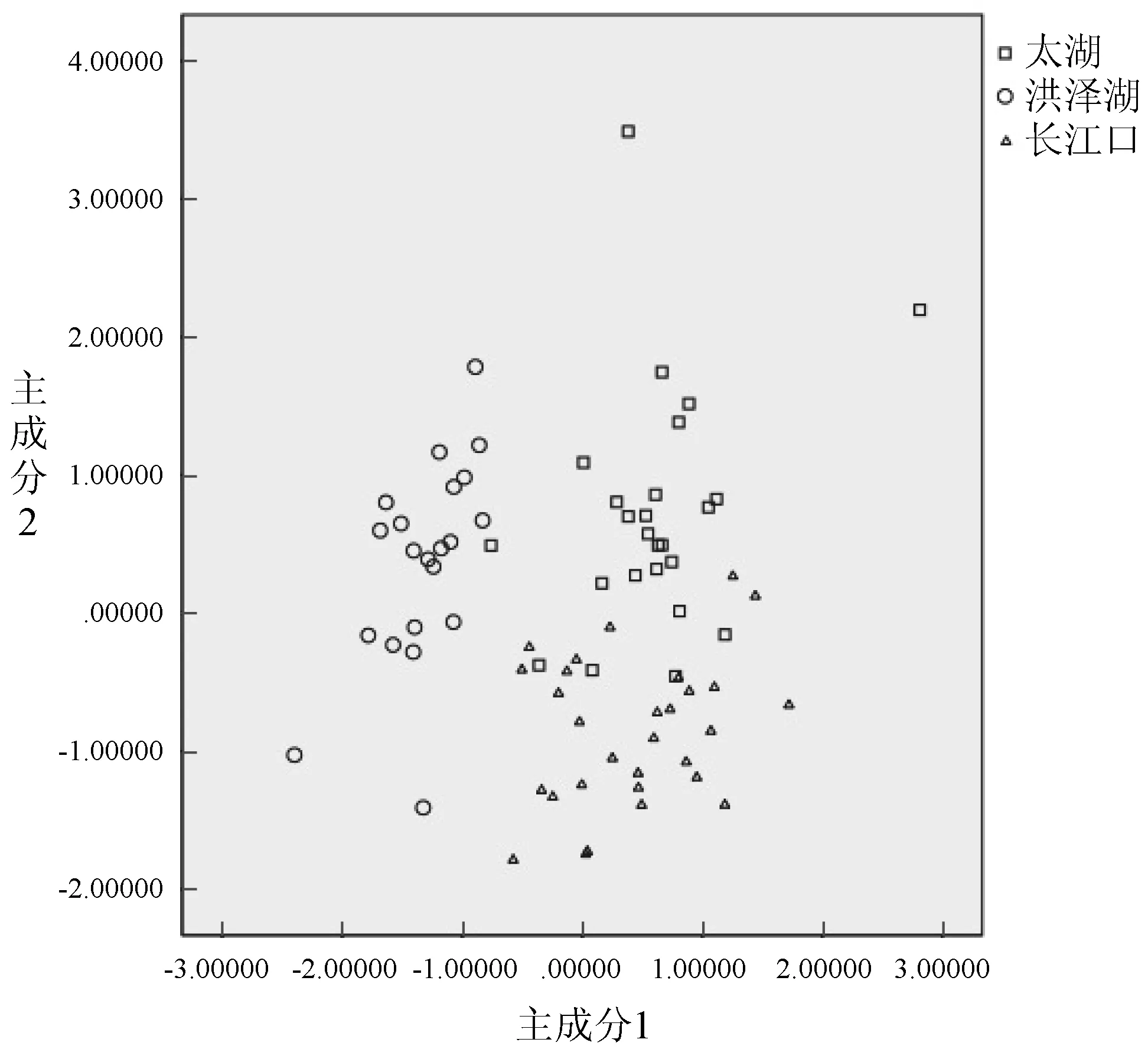
圖2 3個中華鱉群體主成分1和2的散布圖
2.3 判別分析
根據判別分析,獲得貢獻最大的3個參數:體質量/背甲長、后側裙邊寬/背甲長和前肢長/背甲長,即x2、x5和x10。根據這3項分別建立3個群體中華鱉的判別方程如下:
洪澤湖:y=4125.238x2+549.829x5+265.833x10-193.782;
太湖:y=760.663x2+631.306x5+328.143x10-118.654;
長江口:y=1628.811x2+453.206x5+352.951x10-121.181。
當判斷哪個個體的中華鱉屬于某一群體時,將所測數據校正后代入以上3個方程,計算結果最大的即為所屬,所得結果見表4。由表4可知,洪澤湖群體、太湖群體和長江口群體中華鱉的判別準確率分別為100%、100%和93.3%,平均為97.8%,判別準確率很高,說明3個群體之間的差異較大。
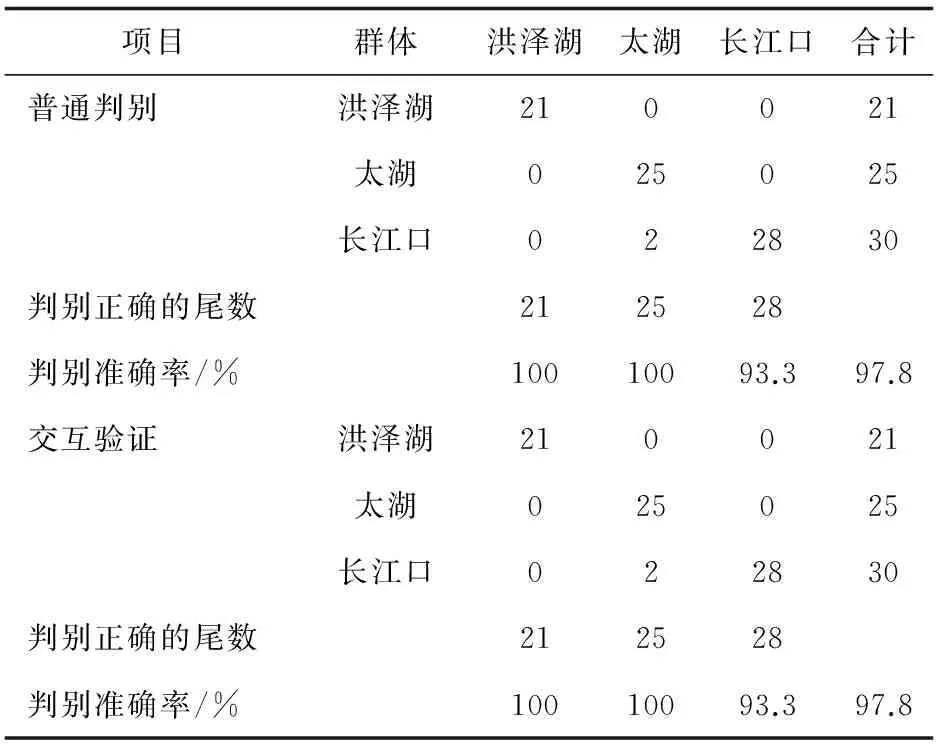
表4 3個群體中華鱉的判別結果
2.4 差異系數檢驗
通過配對樣本T檢驗得出了群體之間各項的平均值、標準差和平均值的標準誤差,然后由差異系數公式得出如下差異系數(表5)。可見洪澤湖與長江口群體間15項形態比例參數差異最大,差異系數大多大于1.28,基本可認為差異達到亞種以上水平;太湖和長江口的形態比例參數差異則比較小,差異未達到亞種以上水平;洪澤湖與太湖群體間差異則處于兩者之間。
3 討 論
本文采用多元分析方法,對中華鱉15項形態比例參數進行研究,從多個層面得出不同地區野生中華鱉群體間的形態多樣性差異。
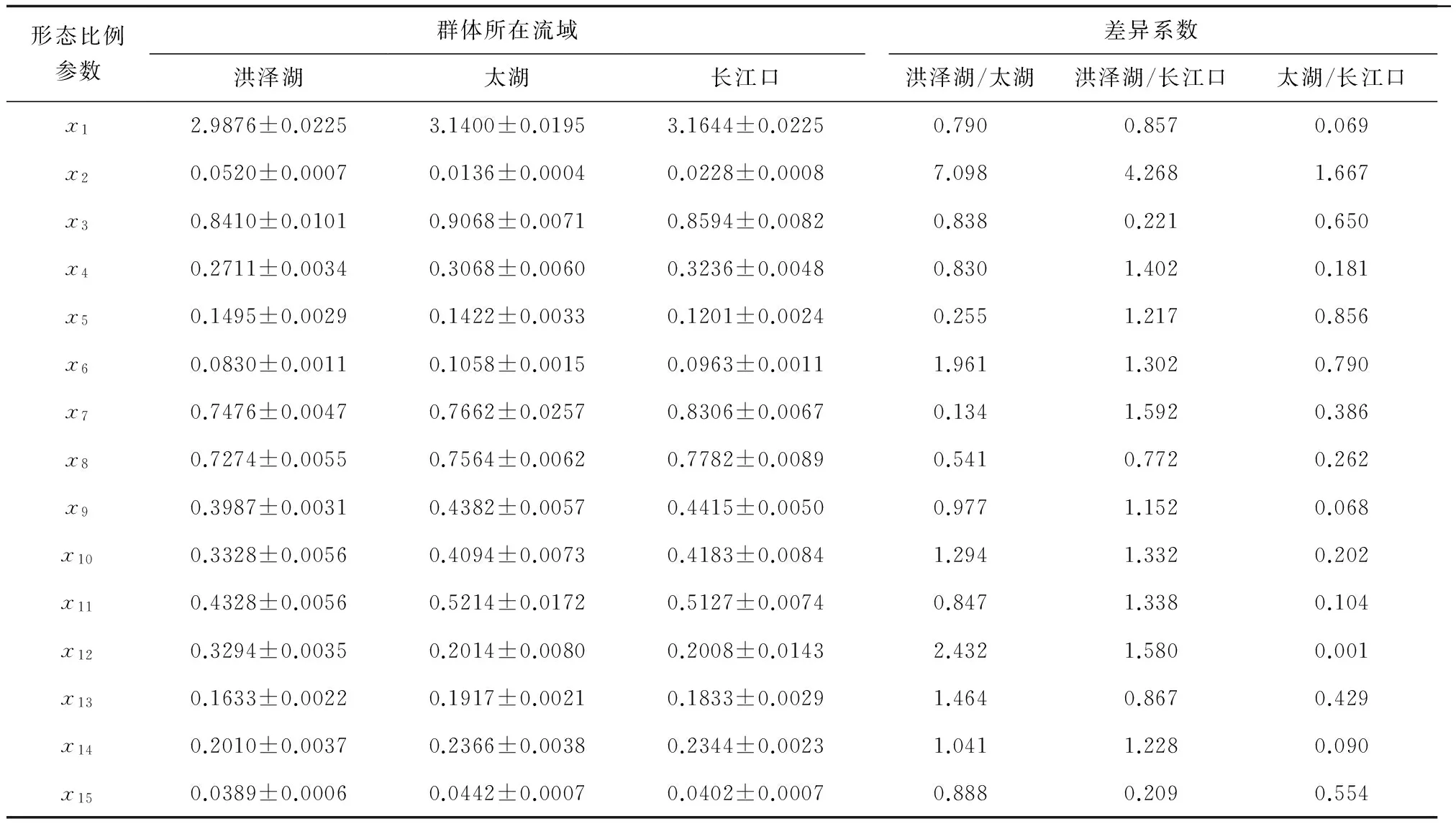
表5 3個不同中華鱉群體間差異比較分析
聚類分析能夠對不同群體進行初步歸類,量化群體間的差異水平,聚類先后的順序反映群體間親緣關系的遠近[12]。本研究中,3個不同地區中華鱉群體的長江口群體和太湖群體首先聚類,再與洪澤湖群體聚合,表明長江口群體和太湖群體之間的親緣關系較洪澤湖群體近,這與其群體分布的地理流域存在顯著的相關性,即長江口和太湖存在一定程度上的流域重疊,且水系也有交織和貫通,而洪澤湖則屬于淮河流域,相對處于隔離狀態。同時也反映出3個不同地區中華鱉群體之間,特別是長江口和太湖群體已經與洪澤湖群體產生了一定程度的形態差異。
主成分分析是將大量參數綜合成為少數因子來說明不同群體的差異大小,并可根據不同群體的主成分數值找出各個群體在各主成分值上差異較大的參數[9]。本研究中,3個主成分分別為56.208%、10.894%和7.165%,其累積貢獻率達到了74.267%,囊括了其總變異的大部分,說明可以通過以上幾個相互獨立因子概括不同種間的形態差異。由主成分1可見,中華鱉頭部的形態差異相比較于其他部位的差異較大,因此頭寬、頭長、眼間距和鼻寬可作為形態判別的主要指標進行群體間判別。
從判定生物種群分化程度的標準以及所測得的差異系數來看,洪澤湖群體和長江口群體之間分類水平已達到亞種以上,而太湖和長江口群體之間分類水平還未達到亞種。同樣,通過散點圖的疏密程度來看,洪澤湖群體與其他兩個群體的差異程度比較大,而長江口和太湖群體的差異程度則較小,這兩種方法得到的結論基本一致。這也說明了形態學方法在中華鱉種間遺傳多樣性的比較和劃分研究中的可行性,具有基礎和直觀的研究特點。
形態差異是生物物種多樣性的一種非常直觀的體現,而通過研究形態差異對生物物種遺傳多樣性進行種質資源評估、保護以及品種改良的研究也有很多[13-17]。于本研究而言,中華鱉3個不同群體形態差異較為明顯,可為中華鱉種質開發和利用提供基礎理論資料。
[1] 王培潮.中國的龜鱉[M].上海:華東師范大學出版社,2000:50-56.
[2] Chu B. The technology of using ground heat for soft-shelled turtle overwintering culture [J]. Proceedings of the International Symposium of Agricultural Engineering, 1989, 89(2):993-994.
[3] Eisenhour D J. Systematics ofMacrhybopsistetranema(Cypriniforms:Cyprinidae) [J]. Copoia, 1999(4):969-980.
[4] Golubtsov A S, Berendzen P B. Morphological evidence for the occurrence of two electric catfish (Matapterurus) species in the White Nile and Omo-Turkana systems (East Africa) [J]. Journal of Fish Biology, 1999, 55(3):492-505.
[5] Gibson A R, Baker A J, Moeed P. Morphometric variation in introduced populations of the common myna (Acridotherestristis):an application of the jackknife to principal component analysis [J]. Systematic Biology, 1984, 33(4):408-421.
[6] Voss R S, Marcus L F, Esealante P. Morphological evolution in muroid rodents I. Conservative patterns of craniometric covariance and their ontogenetic basis in the Neotropical genusZygodontomys[J]. Evolution, 1990, 44(6):1568-1587.
[7] Somers K M. Multivariate allometry and removal of size with principal components analysis [J]. Systematic Zoology, 1986, 35(3):359-368.
[8] 中華人民共和國農業部. GB 21044—2007,中華鱉國家標準[S].北京:中國標準出版社,2007.
[9] 張堯庭,方開泰.多元統計分析引論[M].北京:科學出版社,1982.
[10] 李思發,李晨虹,李家樂.尼羅羅非魚品系間形態差異分析[J].動物學報,1998,44(4):450-457.
[11] Mayr E, Linsley E G, Usinger R L. Methods and principles of systematic zoology [M]. New York and London:Mc Graw Hill, 1953.
[12] 李勤生,蔡慶華,華俐,等.東湖異養細菌群落的分類結構和聚類分析[J].水生生物學報,1991,15(3):242-253.
[13] 潘德博,陳昆慈,朱新平,等.黃喉擬水龜(♀)與三線閉殼龜(♂)雜交后代的形態特征及其與父母本的比較研究[J].水生生物學報,2009,33(4):621-626.
[14] 王偉,陳立僑,顧志敏,等.7個不同翹嘴紅鲌群體的形態差異分析[J].淡水漁業,2007,37(3):41-44.

[16] 明俊超,董在杰,梁政遠,等.6個不同鯉群體的形態差異分析[J].廣東海洋大學學報,2009,29(6):2-6.
[17] 李斌,何俊鋒,區小玲,等.廣西和廣東地區施氏獺蛤3個自然群體的形態差異和遺傳多樣性分析[J].大連海洋大學學報,2011,26(5):414-421.
MorphologicalVariationsAnalysisofDifferentGeographicPopulationsofChineseSoftShelledTurtleTrionyxsinensis
LU Wenhao1, HAN Xiaolei2, CHEN Liang2, XU Jianrong2, HAN Yaoping2
( 1. Fisheries Techniques Guidance Station of Taicang, Taicang 215400,China; 2. Department of Biology and Food Engineering, Changshu Institute of Technology, Changshu 215500,China )
Morphological data of three wild geographic populations of Chinese soft shelled turtleTrionyxsinensiscollected from Yangtze estuary, Tai Lake and Hongze Lake in China were analyzed. Sixteen proportions of morphometric traits were measured from Chinese soft shelled turtle and the degree of morphological variation among populations was calculated using four multivariate analysis. The cluster analysis revealed that the morphometric characters of the Tai Lake population were more similar to those of the populations of Yangtze estuary than to those of the populations of Hongze Lake. In the principal component analysis of the populations, three principal components were constructed, with contributory rate of 56.208% in first principal component, 10.894% in second and 7.165% in the third, and cumulative contributory rate of 74.267%. The accuracy rate of discrimination from Lake was 100% from Yangtze estuary,100% from Tai Lake and from Hongze 93.3%, with mean discrimination accuracy rate of 97.8% from the three populations. All results support that there are a little differences in populations of Yangtze estuary and Tai Lake on morphological characters and that the morphological variations in different populations were mostly exhibited by differences in carapace and abdomen, reaching the level of subspecies between Yangtze estuary Population and Hongze Lake population. This finding provides more reliable theoretical basis for better germplasm resources protection and genetic breeding in artificial breeding.
Trionyxsinensis; multivariation analysis; morphological variation
10.16378/j.cnki.1003-1111.2017.06.016
S966.5
A
1003-1111(2017)06-0784-05
2016-09-27;
2017-01-06.
江蘇省科技支撐計劃(農業)項目(BE2013349).
陸文浩(1983-),男,工程師,碩士;研究方向:水生動物疫病及水產技術示范推廣.E-mail:lwh.nyn@163.com.通訊作者:韓曉磊(1981-),男,實驗師,碩士;研究方向:淡水水生生物學. E-mail:hanxiaolei0724@163.com.
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
