溫度對波紋龍蝦消化酶活力的影響
2017-12-25 18:19:56黃東科梁華芳溫崇慶廖永巖
漁業現代化 2017年6期
黃東科, 梁華芳, 溫崇慶, 廖永巖
(1 廣西北部灣海洋生物多樣性養護重點實驗室,欽州學院,廣西 欽州,5350091;2 廣東海洋大學水產學院,廣東 湛江 5240881)
溫度對波紋龍蝦消化酶活力的影響
黃東科1,2, 梁華芳2, 溫崇慶2, 廖永巖1
(1 廣西北部灣海洋生物多樣性養護重點實驗室,欽州學院,廣西 欽州,5350091;2 廣東海洋大學水產學院,廣東 湛江 5240881)
研究了波紋龍蝦溫度對不同消化器官中消化酶活力的影響,為人工飼料科學配制依據。用酶學分析方法,設計6個溫度梯度(20℃、25℃、30℃、35℃、40℃和45℃),分別測定波紋龍蝦胃、腸和肝胰臟的類胃蛋白酶、胰蛋白酶、淀粉酶和脂肪酶的活力。結果顯示:在反應溫度20℃~45℃范圍內,波紋龍蝦胃、腸、肝胰臟內的消化酶均隨著溫度的升高表現為先升后降趨勢;不同消化器官中胃蛋白酶活力值出現拐點的溫度不一樣,胃、腸和肝胰臟的胃蛋白酶活力最大的溫度分別是30℃、35℃和40℃;不同消化器官胃蛋白酶的活力有顯著差異(P<0.05),大小依次為胃>腸>肝胰臟;波紋龍蝦不同消化器官的類胰蛋白酶出現最大酶活力的溫度相同,為40℃,但胃的類胰蛋白酶活力明顯較腸和肝胰臟的低(P<0.05),差值最大可達40 U/mg;波紋龍蝦胃、腸和肝胰臟的淀粉酶活力在25℃均出現最大值;在消化器官中,腸道淀粉酶活力最大,與胃和肝胰臟的酶活力有顯著性差異(P<0.05);波紋龍蝦胃、腸和肝胰臟內的脂肪酶活力最大的溫度為30℃,活力最高的是肝胰臟,胃內的脂肪酶活力明顯的比腸和肝胰臟的要低(P<0.05)。
波紋龍蝦;溫度;消化酶;活力
環境溫度對變溫動物代謝的影響貫穿于其整個生命過程。水溫影響蝦蟹類的新陳代謝、攝食和飼料轉化率等,從而影響其生長發育和存活[1-2]。在蝦蟹對食物的消化、吸收過程中,消化酶起著關鍵作用,其中的蛋白酶、淀粉酶、脂肪酶等活力受到溫度、pH等因素影響,在十多種蝦蟹類中都得到證明[3-5]。研究不同消化酶的反應條件十分必要。波紋龍蝦(Panulirushomarus)隸屬十足目(Decapoda)、龍蝦科(Palinuridae),龍蝦屬(Panulirus),以其經濟價值高、生長速度快、抗病力強、耐低氧、種苗易獲得等優點而成為主要龍蝦養殖品種之一[6-7]。波紋龍蝦養殖投喂的都是動物性餌料,如蝦蟹類、魚類和貝類等[8-9],尚無人工配合飼料供應,養殖成本高,發展受到制約,急需開發人工配合飼料。目前對龍蝦的消化生理研究較少,國內外關于溫度對龍蝦消化酶影響的研究極少,而溫度對中國龍蝦(Panulirusstimpsoni)、眼斑龍蝦(Panulirusargus)和日本龍蝦(Panulirusjaponicus)消化酶影響報道較多[10-12]。本文就溫度對波紋龍蝦消化酶活力影響開展研究,旨在為波紋龍蝦消化生理研究以及研制人工配合飼料配伍提供參考。
1 材料與方法
1.1 材料
試驗在廣東海洋大學東海島生物研究基地進行。使用經沉淀、砂濾的自然海區海水,鹽度28~30,pH 8.0~8.2。波紋龍蝦(50~60 g)來自海南省瓊海青葛海灣,在20 m3的水泥池中養殖50 d左右,投喂褶牡蠣和蝦等優質餌料。養殖達到試驗規格(95~100 g/尾)后,解剖取出消化器官進行試驗。福林一酚試劑、酪蛋白、酪氨酸、麥芽糖、橄欖油等均由南京建成生物工程研究所提供。
1.2 方法
1.2.1 消化器官解剖處理
波紋龍蝦平均體質量(97.28±8.26) g,以5尾為1組,解剖后取出胃、腸和肝胰臟,提取消化酶,放在6個溫度梯度(20℃、25℃、30℃、35℃、40℃和45℃)中反應,分別測定胃蛋白酶、類胰蛋白酶、淀粉酶活力和脂肪酶的活力。用恒溫水浴鍋調節反應溫度。每個溫度梯度設3個平行組,結果取平均值。
具體操作過程:在冰盤中活體解剖,取出胃、腸和肝胰臟,剝離附著物,剖開,用4℃雙蒸水將消化道內容物沖洗干凈,用濾紙吸干水分,分別稱重后剪碎,加入10倍體積的預冷雙蒸水(4℃),用勻漿器在冰面上進行勻漿,取部分勻漿液直接用于測定脂肪酶活力,其余用高速冷凍離心機離心30 min(4℃,4000 r/min),取上清液(即粗酶液)用于測定龍蝦的胃蛋白酶、類胰蛋白酶和淀粉酶活力。
1.2.2 酶活力測定
參照姜永華等[10]和劉玉梅等[13]的方法,用考馬斯亮藍法G-250,以0.1mg/mL牛血清蛋白溶液為標準物,測定蛋白質濃度。酶活力用比活力表示,定義為每毫克蛋白所含酶活力單位(U/mg)。
(1)胃蛋白酶活力測定。參考潘魯青等[14]的方法 采用以1%酸性酪蛋白(pH2.6)為底物的福林一酚試劑顯色法,以1 mg/mL酪氨酸溶液制作標準曲線。胃蛋白酶活力的定義:在各溫度梯度、pH2.0的條件下,單位粗酶液每分鐘水解干酪素產生1 mg酪氨酸為一個活力單位(U/mg)。
(2)類胰蛋白酶活力測定。參照姜永華等[10]方法,采用以1%堿性酪蛋白為底物的福林一酚試劑顯色法,標準曲線和酶活力定義同胃蛋白酶,pH9.0。
(3)淀粉酶活力測定。采用以1%可溶性淀粉為底物的3,5-二硝基水楊酸顯色法測定淀粉酶的活力[10],以1 mg/mL麥芽糖溶液制作標準曲線。淀粉酶活力的定義:在各溫度梯度及pH6.9的條件下,單位粗酶液每分鐘水解淀粉產生1 mg麥芽糖為一個活力單位U(mg/min)。
(4)脂肪酶活力測定。采用以25%聚乙烯醇橄欖油為底物的標準氫氧化鈉溶液滴定法[13]。三角燒瓶中加入0.025 M磷酸緩沖液(pH 7.5 ) 5毫升,25%聚乙烯醇橄欖油乳化液4 mL,置于40℃水浴中預熱5 min,然后加入酶液,保溫30 min,立即加入95 %乙醇15 mL,終止酶反應。用1%麝香草酚酞做指示劑,標準氫氧化鈉滴定。脂肪酶活力的定義:在各溫度梯度及pH7.5的條件下,單位粗酶液每分鐘水解橄欖油產生1mg脂肪酸為一個活力單位U(mg/min)。
1.2.3 數據處理及統計
數據整理使用Excel 2010。用統計軟件 SPSS17.0進行單因素方差分析及多重比較,各處理平均數之間用鄧肯(Duncan)方法進行差異顯著性比較,以P<0.05為差異顯著,以P<0.01為差異極顯著。
2 結果
2.1 溫度對胃蛋白酶活力的影響
溫度對波紋龍蝦胃、腸、肝胰臟內的胃蛋白酶活力均有顯著影響(P<0.05)。在反應溫度20℃~45℃范圍內,波紋龍蝦胃、腸和肝胰臟的胃蛋白酶均隨著溫度的升高表現為先升后降趨勢,但在不同的部位出現拐點的溫度不一樣。胃蛋白酶活力最大時的溫度分別為30℃、35℃和40℃。在最適溫度下,不同消化器官的胃蛋白酶活力也有顯著差異(P<0.05),胃的最高53.10 U/mg,其次是腸的,最低是肝胰臟42.63 U/mg(表1)。
表1反應溫度對蛋白酶活力的影響
Tab.1 Effects of different reaction temperatures on pepsin enzyme activities in the panulirus homarus

溫度胃/(U/mg)腸/(U/mg)肝胰臟/(U/mg)20℃24 41±1 27F18 93±0 71G23 79±0 52F25℃29 83±0 97E33 95±0 48DE27 73±0 36EF30℃53 10±0 29A42 67±0 68C30 64±1 22E35℃51 18±0 63A46 98±3 28B36 65±1 03D40℃31 36±0 14E41 81±2 58C42 63±0 49BC45℃16 82±2 02G28 81±1 13E33 57±0 78DE
注:同列字母不同,表示差異顯著(P<0.05);同列字母相同表示差異不顯著。
2.2 溫度對類胰蛋白酶活力的影響
溫度對波紋龍蝦胃、腸和肝胰臟的類胰蛋白酶活力均有顯著影響(P<0.05)。在20℃~45℃范圍內,波紋龍蝦胃、腸和肝胰臟的類胰蛋白酶均隨著溫度的升高表現為先升后降趨勢,不同的消化器官出現最大酶活力的溫度一致,均為40℃。其中,胃的類胰蛋白酶活力明顯較腸和肝胰臟的低(P<0.05),差值最大可達40 U/mg(表2)。
表2反應溫度對類胰蛋白酶活力的影響
Tab.2 Effects of different reaction temperatures on tryptase enzyme activities in the panulirus homarus
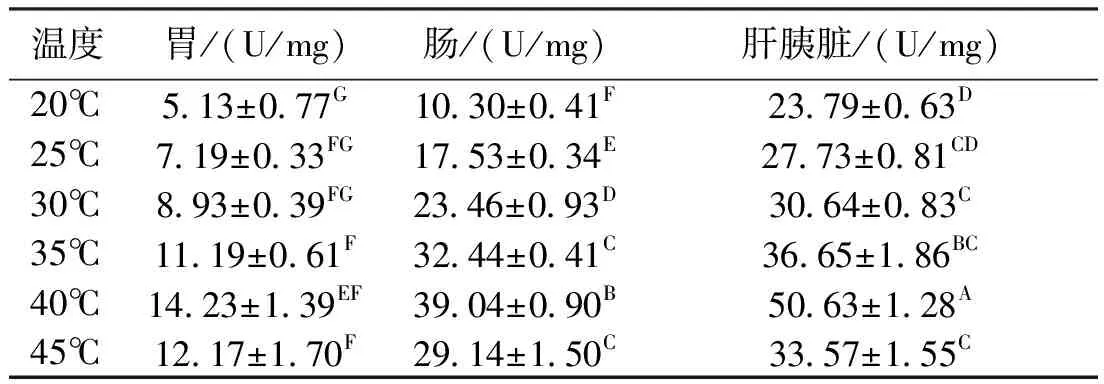
溫度胃/(U/mg)腸/(U/mg)肝胰臟/(U/mg)20℃5 13±0 77G10 30±0 41F23 79±0 63D25℃7 19±0 33FG17 53±0 34E27 73±0 81CD30℃8 93±0 39FG23 46±0 93D30 64±0 83C35℃11 19±0 61F32 44±0 41C36 65±1 86BC40℃14 23±1 39EF39 04±0 90B50 63±1 28A45℃12 17±1 70F29 14±1 50C33 57±1 55C
注:同列字母不同,表示差異顯著(P<0.05);同列字母相同表示差異不顯著。
2.3 溫度對淀粉酶活力的影響
溫度對波紋龍蝦胃、腸、肝胰臟內的淀粉酶活力均有顯著影響(P<0.05)(表3)。在反應溫度20℃~45℃范圍內,波紋龍蝦胃、腸、肝胰臟內的淀粉酶出現最大酶活力的溫度一致,為25℃,較蛋白酶的溫度低;其中20℃~35℃各組淀粉酶的活力沒有顯著性差異。腸的淀粉酶活力最大,與胃和肝胰臟的酶活有顯著性差異(P<0.05)。
表3反應溫度對淀粉酶活力的影響
Tab.3 Effects of different reaction temperatures on amylase activities in the panulirus homarus

溫度胃/(U/mg)腸/(U/mg)肝胰臟/(U/mg)20℃6 03±1 34BC16 70±0 83A7 33±1 05BC25℃7 52±0 83BC18 61±0 53A8 77±0 64B30℃7 10±1 02BC17 33±0 74A8 16±1 65B35℃6 05±1 04BC14 07±0 66A7 20±1 01BC40℃6 21±0 62BC9 80±0 64B6 50±1 12BC45℃4 78±0 73C8 62±0 55B4 45±1 17C
注:同列字母不同,表示差異顯著(P<0.05);同列字母相同表示差異不顯著。
2.4 溫度對脂肪酶活力的影響
溫度對波紋龍蝦胃、腸和肝胰臟內的脂肪酶活力均有顯著影響(P<0.05)(表4)。在反應溫度20℃~45℃范圍內,波紋龍蝦胃、腸和肝胰臟的脂肪酶出現最大酶活力的溫度都是30℃;胃的脂肪酶活力明顯比腸和肝胰臟的要低(P<0.05);脂肪酶活力最高的部位是肝胰臟。
表4反應溫度對脂肪酶活力的影響
Tab.4 Effects of different reaction temperatures on lipase activities in the panulirus homarus

溫度胃/(U/mg)腸/(U/mg)肝胰臟/(U/mg)20℃2 72±1 18C4 61±0 92C6 57±1 12B25℃4 44±0 52C6 81±0 80B11 02±0 65A30℃4 93±0 72C8 09±0 33B12 53±1 23A35℃4 80±0 63C7 69±0 92B10 88±1 02AB40℃3 15±1 02C4 90±1 10C8 17±0 92B45℃2 00±0 52C2 82±0 83C4 98±1 06C
注:同列字母不同,表示差異顯著(P<0.05);同列字母相同表示差異不顯著。
2.5 淀粉酶/蛋白酶比值
在最適溫度條件下對4種主要消化酶活力進行比較(表5)并以大小順序排列,胃蛋白酶活力:胃>腸>肝胰臟;類胰蛋白酶活力:肝胰臟>腸>胃;淀粉酶活力:肝胰臟>腸>胃;脂肪酶活力:肝胰臟>腸>胃。以淀粉酶/蛋白酶(A/P) 比值作為動物食性和營養狀況的一個指標。A/ P1比值和A/ P2比值均遠小于1,說明肝胰臟、腸道、胃中淀粉酶的比活力均小于蛋白酶的比活力,蛋白酶占主導地位。

表5 不同器官主要消化酶活性比較及A/P值分析
注:同列字母不同,表示差異顯著(P<0.05);同列字母相同表示差異不顯著。
3 討論
3.1 關于蛋白酶活力
胃蛋白酶和類胰蛋白酶是甲殼動物幼體的 2 種主要蛋白水解酶,在對蝦幼體發育過程中,后者是一種最占優勢的消化酶[14,16]。中國明對蝦、日本囊對蝦、三疣梭子蟹(Portunustrituberculatus)和中華絨螯蟹(Eriocheirsinensis)4種甲殼動物幼體的相關研究結果表明,類胰蛋白酶比胃蛋白酶活力大,約為1.2~2.8 倍[14,17],但在中國龍蝦[10]和本研究的波紋龍蝦中,類胰蛋白酶并不是優勢酶,在胃和腸中類胰蛋白酶比胃蛋白酶活力小。可見,在蝦的不同發育階段,其消化酶活力隨著食性轉變和營養需求不同而發生變化。
動物的消化能力決定了其對營養物質消化吸收的效率,消化酶活力的高低直接影響動物對營養物質的消化能力從而影響生長。酶促反應受到溫度的影響,本試驗結果表明,反應溫度對波紋龍蝦蛋白酶活力有顯著影響。波紋龍蝦胃蛋白酶的最適反應溫度為30℃~40℃,與中國龍蝦的最適反應溫度(31℃~47℃)[10]和克氏原螯蝦(Procambarusclarkia)的最適溫度(35℃)相近,但遠低于凡納濱對蝦的50℃~65℃和中國明對蝦的45℃~55℃[19-20],以及眼斑龍蝦[11]和日本龍蝦[12]的60℃。另外,本試驗結果表明,波紋龍蝦類胰蛋白酶的最適反應溫度均為40℃,與中國龍蝦類胰蛋白酶的最適反應溫度39℃的結果相似[10],而與眼斑龍蝦類胰蛋白酶的最適反應溫度60℃和中國明對蝦的47℃有較大差異[11,20]
3.2 關于脂肪酶活力
本試驗表明,溫度對脂肪酶活力有較大影響。波紋龍蝦各消化器官的脂肪酶活力在30℃時最高,但中國龍蝦[10]不同消化器官的脂肪酶最適反應溫度不同,腸和肝胰臟的均為31℃,與波紋龍蝦接近,但胃的脂肪酶活力在23℃時最高,低于波紋龍蝦的30℃,而這兩種龍蝦的脂肪酶最適反應溫度比眼斑龍蝦[11]的40℃低得多。不同種類的蝦,其最適反應溫度也不相同。如:克氏原螯蝦脂肪酶活力在肝胰臟、腸道、胃的最適溫度分別為 25℃、35℃和25℃[18],而凡納濱對蝦的最適溫度均為 37℃[19]。據報道,胃、肝胰臟和腸道脂肪酶的最適溫度分別為 50℃、30℃和 40℃[21];中國明對蝦與日本囊對蝦脂肪酶最適溫度均為 35℃~40℃[20]。它們間的差異除了種類不同外,還可能與其棲息環境密切相關,如波紋龍蝦和中國龍蝦分布區重疊,最適溫度相近,而與眼斑龍蝦地理分布(美國東海岸、加勒比海和墨西哥灣)相距甚遠,最適溫度相差較大。
3.3 關于淀粉酶活力
波紋龍蝦淀粉酶最適反應溫度為25℃,與中國龍蝦[10]的23℃較為接近,但與眼斑龍蝦的最適溫度50℃相差較大[11],進一步說明了地理分布不同可能是造成消化酶活力差異的原因。另外,波紋龍蝦淀粉酶最適反應溫度也與日本囊對蝦的25℃~30℃相近,但低于凡納濱對蝦[19]、中國明對蝦[20]的30℃~35℃,克氏原螯蝦的35℃~40℃[18]和三疣梭子蟹[22]的55℃~65℃,其棲息環境與淀粉酶反應溫度或多或少有點負相關。
3.4 關于龍蝦食性偏向性
波紋龍蝦的4種消化酶活性表現為胃蛋白酶活力最高,類胰蛋白酶次之,脂肪酶最小。Biesiot等[23]提出采用淀粉酶/胃蛋白酶(A/ P1) 比值和淀粉酶/ 胰蛋白酶(A/ P2) 比值作為食性和營養狀況的指標,當比值高于1時為植物食性或偏植物食性,比值低于1時為動物食性或偏動物食性。如對蝦類的幼體,溞狀幼體期的A/P>1,與其以單胞藻為食的植物食性相符,而幼體發育到糠蝦期時,其A/P<1,轉變為動物食性[3,14-15,24-25]。本試驗發現,波紋龍蝦的A/ P1和A/ P2比值在胃、腸和肝胰臟中都小于1。根據以上推論,波紋龍蝦的食性應為動物食性或偏動物食性,這與事實相符。可見,用A/P比值作為判斷甲殼動物食性的指標是可行的。
4 結論
波紋龍蝦不同的消化器官,其消化酶活力也不同。胃蛋白酶活力大小依次為胃>腸>肝胰臟,類胰蛋白酶活力依次為肝胰臟>腸>胃,淀粉酶活力依次為腸>肝胰臟≈胃,脂肪酶活力依次為肝胰臟>腸>胃。溫度對波紋龍蝦消化酶活力有較大影響,在反應溫度20℃~45℃范圍內,胃、腸和肝胰臟內的消化酶均隨著溫度的升高表現為先升后降趨勢。波紋龍蝦消化酶最適反應溫度在不同消化器官中有差異,胃、腸和肝胰臟胃蛋白酶的最適反應溫度分別為30℃、35℃和40℃,而胃、腸、肝胰臟3個消化器官的類胰蛋白酶活力最大的反應溫度均為40℃,在胃、腸、肝胰臟中淀粉酶最反應溫度都是25℃,脂肪酶最適反應溫度為30℃。通過淀粉酶和蛋白酶比值(A/P<1)為動物食性或偏動物食性分析,對比波紋龍蝦的偏動物食性習性和本研究結果,認為用A/P比值作為判別甲殼動物食性指標是可行的。
□
[1] 黃東科,梁華芳,張志.溫度對波紋龍蝦存活、攝食、蛻殼和生長的影響[J].生態學報,2017,37(18): 5973-5980.
[2] 張龍崗,鐘君偉,朱永安.溫度對克氏原螯蝦苗種生長和存活的影響[J].河北漁業,2015(1):4-5.
[3] 潘魯青,劉泓宇,肖國強.甲殼動物幼體消化酶研究進展[J].中國水產科學,2006,13(3):492-501.
[4] LUO W,ZHAO Y L,ZHOU Z L,et.al.Digestive enzyme activity and mRNA level of trypsin in embryonic red claw crayfish,Cherax quadricarnatus[J].Chinese Journal of Oceanology and Limnology,2008,26(1):62-68.
[5] 姜鶴,孔繁東,劉兆芳.溫度、pH及金屬離子對大連蝦蛄消化酶活性的影響[J].保鮮與加工,2016,16(4):84-88.
[6] 梁華芳,趙運添,黃東科.波紋龍蝦耗氧率和窒息點的研究[J].水產養殖.2012(4):1-4.
[7] KEMP J O,BRITZ P J.The effect of temperature on the growth,survival and food consumption of the east coast rock lobster Panulirus homarus rubellus[J].Aquaculture ,2008,280:227-231.
[8] 沈銘輝,符芳霞,范薇.不同餌料對波紋龍蝦生長影響的研究[J].熱帶農業科學,2011,31(7):55-59.
[9] 沈銘輝,范薇.波紋龍蝦工廠化養殖技術[J].中國水產,2008(5):52,59.
[10] 姜永華,顏素芬.反應溫度對中國龍蝦消化酶活力的影響[J].集美大學學報(自然科學版),2009(1):14-19.
[11] ERICK PERERA,MOYANO F J,DAZ M,et.al.Polymorphism and partial characterization of digestive enzymes in the spiny lobster Panulirus argus[J].Comparative Biochemistry and Physiology,Part B ,2008,150(3):247-254.
[12] GALGANI F,NAGAYAMA F.Digestive proteinases in the Japanese spiny lobster Panulirus japonicus[J].Comparative Biochemistry and Physiology Part B: Comparative Biochemistry,1987,87(4):889-893.
[13] 劉玉梅,朱謹釗.對蝦消化酶的研究[J].海洋科學,1984(5):46-50.
[14] 劉玉梅,朱謹釗.中國對蝦幼體和仔蝦消化酶活力及氨基酸組成的研究[J].海洋與湖沼,1991,22(6):571-575.
[15] 潘魯青,王克行.中國對蝦幼體消化酶活力的實驗研究[J].水產學報,1997,21(1):26-31.
[16] TSAI I H,CHUANG K L ,CHUANG J L.Chymotrypsins in digestive tracts of crustacean decapods(shrimps) [J].Comp Biochem Physiol,1986,85:235-240.
[17] 潘魯青.四種蝦蟹類幼體消化酶活力的比較研究[J].青島海洋大學學報,1997,27(3): 313-318.
[18] 祝堯榮,壽建昕,沈文英.溫度對克氏原螯蝦消化酶活性的影響[J].浙江農業學報,2009,21(3): 238-240.
[19] 沈文英,胡洪國,潘雅娟.溫度和pH值對南美白對蝦(Penaeusvannmei)消化酶活性的影響[J].海洋與湖沼,2004,35(6): 543-548.
[20] 吳垠,孫建明,周遵春.溫度對中國對蝦、日本對蝦主要消化酶活性的影響[J].大連水產學院學報,1997,12(2):15-22.
[21] 黃燕華,王國霞,劉襄河,等.溫度和pH對南美白對蝦主要消化酶活性的影響[J].華南農業大學學報,2008,29(4):87-90,94.
[22] 胡毅,潘魯青.三疣梭子蟹消化酶的初步研究[J].中國海洋大學學報(自然科學版),2006,36(4): 621-626.
[23] BIESIOT P M,CAPUZZO J M.Changes in digestive enzyme activities during early development of the American lobster Homarus americanus Milne Edwards [J].Journal of Experimental Marine Biology and Ecology,1990,136(2): 107-122.
[24] 朱春華,李廣麗,文海翔.南美白對蝦早期幼體消化酶活力的研究[J].海洋科學,2003,27(5):54-57.
[25] 潘魯青,王偉.日本對蝦幼體幾種消化酶活力的研究[J].海洋湖沼通報,1997(2):15-18.
Effectsofdifferenttemperaturesonthedigestiveenzymeactivitiesinthepanulirushomarus
HUANGDongke1,2,LIANGHuafang2,WENChongqing2,LIAOYongyan2
(1GuangxiKeyLaboratoryofBeibuGulfMarineBiodiversityConservation,QinzhouUniversity,Qinzhou535009,China;2FisheriescollegeofGuangdongOceanUniversity,Zhanjiang524088,China)
To provide an basis for scientific formula of artificial diets,the effects of different temperatures on the digestive enzyme activities in the panulirus homarus were investigated.Using enzymatic analysis method,the four enzymes,including pepsin,trypase,amylase and lipase,were determined in the digestive organs of stomach,intestines,hepatopancreas in the panulirus homarus at temperatures of 20℃,25℃,30℃,35℃,40℃ and 45℃.All activities of the four kinds of digestive enzymes presented a trend of first increasing and then decreasing with temperatures ranging from 20℃to 45℃.The highest reaction temperature differed for pepsin reaching the turning point in different digestive organs,with values of 30℃,35℃,40℃ corresponded to stomach,intestines and hepatopancreas,respectively.And there were significant differences(P<0.05)in pepsin among the three tissues,of which stomach presented the maximum level,while hepatopancreas had the lowest one.The temperature for the presentation of the maximal activity of trypsase in different digestive organs were same at 40℃,and level of trypsase in stomach was significantly lowerP<0.05)than those in intestines and hepatopancreas,with the interval was up to 40 U/mg.The amylase activity exhibited maximal in intestine,which differed significantly(P<0.05) from those in other two organs,but the highest reaction temperature for activity of amylase in all three digestive organs were 25℃.The activities of lipase in all digestive organs had the same optimal reaction temperature of 30℃,when the activity of lipase was highest in hepatopancreas,whereas stomach presented a lower lipase activity(P<0.05).
panulirus homarus;temperature;digestive enzyme;activity
10.3969/j.issn.1007-9580.2017.06.006
2017-09-06
廣西北部灣海洋生物多樣性養護重點實驗室(欽州學院)開放課題(2015KB03);廣東省科技廳農業攻關項目(2013B020308009)
黃東科(1988-),男,碩士,研究方向:水產養殖技術。E-mail: 522865834@qq.com
梁華芳(1965—),男,教授,研究方向:甲殼動物生物學及養殖技術。E-mail:hfliang@126.com
S968.22
A
1007-9580(2017)06-032-06
