沙棘果肉發育期油脂合成積累的源匯基因協同表達
2018-01-03 00:51:52阮成江管文柯單金友吳雨蹊吳天忠
林業科學研究 2017年6期
關鍵詞:油脂
丁 健,阮成江*,關 瑩,管文柯,單金友,吳雨蹊,吳天忠
(1.大連民族大學 資源植物研究所,遼寧 大連 116600; 2.黑龍江省農業科學院 漿果研究所,黑龍江 綏棱 152200;3.新疆林業科學院,新疆 烏魯木齊 830064)
沙棘果肉發育期油脂合成積累的源匯基因協同表達
丁 健1,阮成江1*,關 瑩2,管文柯3,單金友2,吳雨蹊2,吳天忠3
(1.大連民族大學 資源植物研究所,遼寧 大連 116600; 2.黑龍江省農業科學院 漿果研究所,黑龍江 綏棱 152200;3.新疆林業科學院,新疆 烏魯木齊 830064)
目的探討沙棘果肉油脂合成積累與源匯基因表達的關系。方法以8個不同發育時期的近緣高油品系‘TF2-36’和低油品系‘雜56’果肉為材料,利用氯仿甲醇法測定含油率,采用qRT-PCR技術分析油脂合成源基因(GPD1)和匯基因(DGAT1和DGAT2)在近緣高低油果肉間的表達差異及其對油脂合成積累的影響。結果研究表明:(1)沙棘果肉含油率呈先上升后穩定趨勢,‘TF2-36’的果肉含油率一直高于‘雜56’;(2)GPD1、DGAT1和DGAT2基因在‘TF2-36’果肉發育期間中均有明顯高于‘雜56’的表達量峰值,但GPD1表達量峰值出現在油脂快速合成期,DGAT1和DGAT2表達量峰值出現在油脂穩定積累期。GPD1在發育前期高表達,促進合成更多的TAG前體G3P,而DGAT1和DGAT2在發育后期高表達,則促進了TAG的高積累。結論沙棘果肉高油脂積累源于源基因“GPD1”和匯基因“DGAT1和DGAT2”的協同高表達,研究結果為理解沙棘非種子組織(果肉)油脂合成機理提供了理論依據。
沙棘;油脂合成;源基因;匯基因;基因表達

阮成江等[18]研究表明TAG的生物合成與源基因“GPD1”和匯基因“DGAT”相關。GPD1基因是甘油酯合成通路中的關鍵限速酶基因,是油脂合成的“源基因”,具有調控TAG合成底物3-磷酸甘油含量的作用[9,18];DGAT為甘油二酯酰基轉移酶,是催化合成TAG的關鍵限速酶,也是植物油脂合成的“匯基因”[12,18]。同時調控源匯基因的表達可提高植物種子含油率,解析二者的相互作用機制對提高油脂產量和品質具有重要作用。隨后有學者將脂肪酸的生物合成形容為“推”,將脂肪酸組裝合成TAG形容為“拉”[19],共同表達“推”和“拉”基因可明顯提高油脂含量。因此,分析沙棘果肉油脂合成積累的源基因“GPD1”和匯基因“DGAT1和DGAT2”的表達與含油率的關系,對提高沙棘果肉油脂具有重要意義。
本研究以近緣高油品系‘TF2-36’和低油品系‘雜56’果肉為材料,分析不同時期的果肉含油率,利用qRT-PCR研究源基因“GPD1”和匯基因“DGAT1和DGAT2”的表達模式及其在高低油果肉中的表達差異,揭示高低油果肉的油脂合成積累與源匯基因表達的關系,為深入理解沙棘非種子組織(果肉)油脂合成提供科學依據。
1 材料與方法
1.1 實驗材料
以‘TF2-36’(蒙古沙棘亞種)和‘雜56’(蒙古沙棘和中國沙棘雜交種)為試驗材料,二者間基于ISSR標記分析的遺傳相似系數為0.752[20],果實分別于2015年6月25日、7月6日、7月17日、7月28日、8月8日、8月19日、8月30日和9月10日采自黑龍江省農業科學院漿果研究所。各品系樣品采自3株無性繁殖植株的多個部位,同植株果實混合,用錫紙包裹后置于液氮中速凍。樣品運抵大連民族大學資源植物研究所,保存于-80℃冰箱備用。
1.2 方法
1.2.1 果肉含油率測定 采用氯仿甲醇法[2,21-22]測定不同時期沙棘果肉含油率:冷凍干燥的果肉粉末轉移至玻璃試管中,加入甲醇和氯仿(均為色譜純,Honeywell公司)漩渦混勻后超聲30 min,上清液轉移到新試管中,殘渣用氯仿甲醇(體積濃度百分比2∶1)再次提取,合并的上清液加入其1/4體積的氯化鉀溶液(質量濃度0.88%),收集下層液至玻璃樣品瓶中,揮發至恒質量。含油率(%)=(m1-m2)/m×100;m1為油脂和玻璃樣品瓶的質量/g;m2為玻璃樣品瓶的質量/g;m為干燥樣品粉末的質量/g,實驗設3次生物學重復。
1.2.2 qRT-PCR檢測 參照柱式植物總RNA提取試劑盒(上海生物工程有限公司)方法提取沙棘果肉總RNA,根據PrimeScriptTMRT reagent Kit with gDNA Eraser試劑盒(大連寶生物公司)方法合成第一鏈cDNA[23]。本研究前期構建了沙棘種子、果肉、葉、莖和根轉錄組,獲得了大量的功能基因注釋以及差異表達基因信息。利用篩選獲得的目的基因片段和PrimerQuest在線軟件設計特異引物(表1)。參照SYBR Premix Ex TaqTMII(Tli RNaseH Plus)試劑盒(大連寶生物公司)方法和ABI7500 Real time PCR儀(美國Applied Biosystems公司)推薦程序進行qRT-PCR[11],以沙棘UBQ5為內參基因[4],采用2-ΔΔCt方法分析目的基因相對表達量[24]。實驗設3次生物學重復。
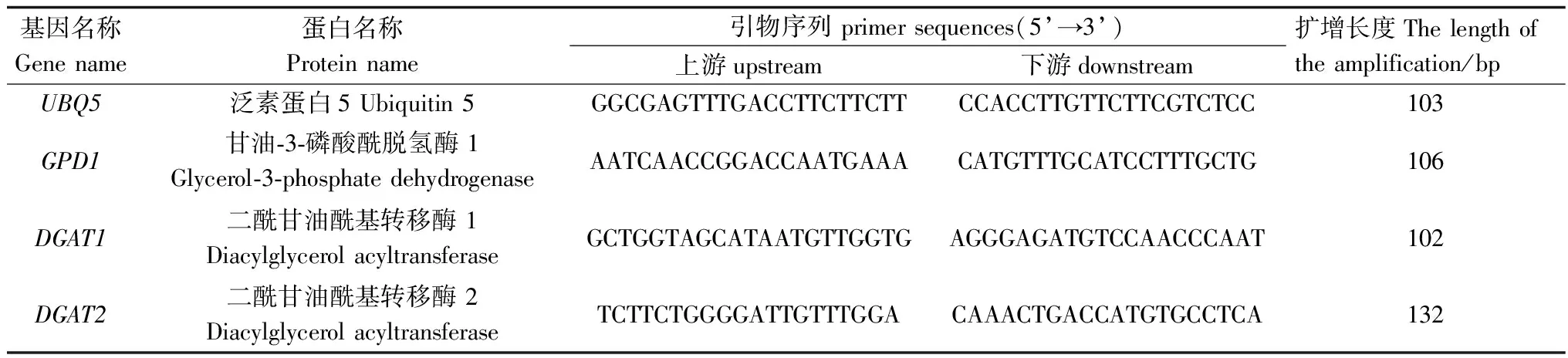
表1 基因名稱及qRT-PCR引物
1.2.3 數據統計分析 利用SPSS 20.0軟件進行單因素方差分析和LSD法進行差異性檢驗,采用EXCEL2010進行作圖。
2 結果與分析
2.1 沙棘果肉發育過程中含油率的動態變化
隨著果實的增大,果肉顏色由綠色轉為黃綠色,再轉為黃色(‘雜56’)或桔黃色(‘TF2-36’,圖1)。果肉含油率總體呈上升趨勢,‘TF2-36’干果肉含油率一直高于‘雜56’(圖2),最大增幅均發生在7月28日至8月8日期間,但‘TF2-36’干果肉含油率增速明顯大于‘雜56’。8月30日‘TF2-36’干果肉含油率達高峰(37.19%),約為‘雜56’的3.9倍(圖2)。

圖1 品系‘TF2-36’和‘雜56’果實形態和顏色變化Fig.1 Morphology and color of fruits of lines ‘TF2-36’ and ‘Za56’
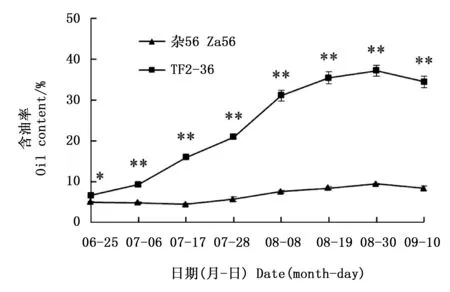
注:*和**分別表示同一時期兩個品系間含油率在0.05和0.01水平上的顯著差異Note: * and ** indicated significant differences of oil content between two lines at the same harvest time at the level of 0.05 and 0.01, respectively圖2 品系‘TF2-36’和‘雜56’果肉發育期間的含油率變化Fig.2 Changes of oil contents in fruit pulp of lines ‘TF2-36’ and ‘Za56’
2.2 GPD1、DGAT1和DGAT2基因表達分析
GPD1基因在‘TF2-36’果肉中先上調表達,在7月28日到達峰值;隨后,表達下調,并逐漸穩定。GPD1基因在‘雜56’果肉中呈總體下調表達模式。‘TF2-36’果肉的GPD1基因表達量一直顯著高于‘雜56’,且在7月28日達峰值(圖3A),兩個品系的GPD1基因表達量峰值約相差3.4倍,與含油率峰值相差倍數相似。
DGAT1基因在‘TF2-36’果肉中先下調表達后上調再微量下調,而在‘雜56’果肉中呈先下調再上調表達模式。‘TF2-36’果肉的DGAT1基因表達量一直顯著高于‘雜56’(圖3B),且與‘TF2-36’干果肉含油率顯著高于‘雜56’相一致。DGAT2基因在‘TF2-36’果肉中先穩定表達,在8月30日達峰值且顯著高于‘雜56’。‘TF2-36’果肉的DGAT2基因表達量和干果肉含油率達峰值的時間相同(圖2)。
與低果肉油品系‘雜56’相比,源基因GPD1在高果肉油品系‘TF2-36’發育初期的高表達(圖3A),為TAG合成提供了更多的前體G3P;且匯基因DGAT1在GPD1高表達后,仍維持高表達水平(圖3B),加速TAG組裝。可見,源匯基因的協同高表達促進了TAG合成積累。
3 討論
沙棘非種子組織(果肉)高積累棕櫚油酸的特性在自然界中非常罕見,它是我國唯一高積累棕櫚油酸的植物,但果肉較低的含油率嚴重限制了其有效開發利用。
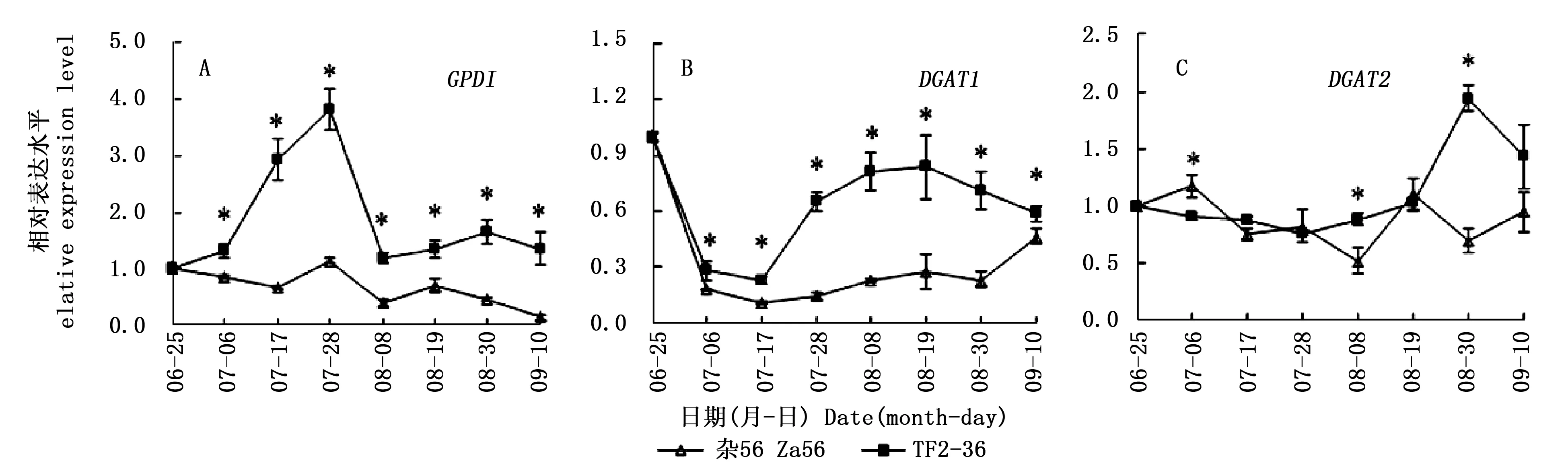
注:*表示同一采收期兩個品系間的基因相對表達量在0.05水平上的顯著差異Note: * indicated significant differences of gene relative expression level between two lines at the same harvest time at the level of 0.05圖3 GPD1、DGAT1和DGAT2基因在‘TF2-36’和‘雜56’果肉中的表達差異Fig.3 Differences of GPD1, DGAT1 and DGAT2 genes expression in fruit pulp between the lines ‘TF2-36’ and ‘Za56’
‘TF2-36’果肉的GPD1基因表達量一直顯著高于‘雜56’,且于7月28日達峰值,促使‘TF2-36’的干果肉含油率在8月30日達峰值且顯著高于‘雜56’,含油率的峰值滯后于GPD1基因表達量的峰值,而且兩個品系的干果肉含油率峰值相差倍數(3.9倍)與GPD1基因表達量峰值相差倍數(3.4倍)相近。可見,GPD1基因在‘TF2-36’果肉發育前期的高表達,為油脂合成積累了更多的G3P[7],進而提升其果肉含油率。近年來,在麻瘋樹(JatrophacurcasL.)和印加果(PlukenetiavolubilisL.)種子中也發現GPD基因表達量與TAG含量變化相關[23,25],且注射外源甘油的油菜種子油脂含量高峰也滯后于G3P高峰[10]。
高油品系‘TF2-36’果肉的匯基因DGAT1和DGAT2在源基因GPD1的前期高表達后出現表達量高峰,且顯著高于低果肉油品系‘雜56’,表明DGAT1和DGAT2基因在果肉發育期間的高表達也促進了油脂高積累。發育期的麻瘋樹和蓖麻(RicinuscommunisL.)DGAT1基因以及印加果DGAT2基因表達量呈明顯的單峰曲線變化規律,且與油脂合成相關[23,25-26]。發育期文冠果胚的油脂積累與XsDGAT1和XsDGAT2基因表達模式相關,與非轉基因植株相比,異源表達文冠果XsDGAT1和XsDGAT2基因的擬南芥種子油脂含量分別提高71.6 μg·mg-1(20.3%)和30.9 μg·mg-1(8.8%)[11]。不同物種和組織對DGAT1和DGAT2基因的響應程度存在一定差異,而它們對沙棘果肉含油率的影響程度還需要進一步驗證。吳永美等[27]將貓爪草(Doxanthaunguis-cati)的Δ9D(delta-9 desaturase)基因(催化棕櫚酸去飽和為棕櫚油酸)和對棕櫚油酸有特異選擇性的DGAT酶基因在大豆中共同表達發現,大豆體細胞胚的棕櫚油酸含量上升到19%,比單獨轉化Δ9D基因高7%。Li等[16]發現油桐中有催化桐油酸(十八碳三烯酸)與TAG特異結合的DGAT2基因。因此在后續研究中獲得對棕櫚油酸有特異選擇性的DGAT基因對進一步解析沙棘果肉油脂合成積累機理具有重要意義。
與低果肉油品系‘雜56’相比,高果肉油品系‘TF2-36’的源基因GPD1在發育前期高表達,而匯基因DGAT1和DGAT2在發育后期出現高表達,源和匯基因的協同高表達促進了沙棘果肉油脂的高積累。在煙草中共表達擬南芥的WRI1和DGAT1基因可產生協同作用,轉基因煙草的干葉TAG含量可提高約100倍(2.48%),是轉化單一基因的5倍左右[19]。Chen等[17]將油桐的VfFAD2和VfDGAT2基因融合導入紅酵母和擬南芥中,發現它們具有協同促進不飽和脂肪酸積累的作用,且可獲得更高含量的C18:3(紅酵母中提高174%,擬南芥中提高14.6%)。非種子組織(果肉)的油脂生物合成途徑復雜,同時涉及到種子油脂的合成,這些結構基因和調節基因對沙棘果肉油脂積累的影響正在進一步的研究驗證之中。
4 結論
(1)品系‘TF2-36’的果肉含油率一直顯著高于‘雜56’,兩個品系的果肉油脂積累模式相似;(2)GPD1、DGAT1和DGAT2基因在‘TF2-36’果肉中均有明顯高于‘雜56’的表達量峰值,但GPD1表達量峰值出現在油脂快速合成期,DGAT1和DGAT2表達量峰值出現在油脂穩定積累期,源基因“GPD1”在發育前期高表達,促進合成更多的TAG前體G3P,而匯基因“DGAT1和DGAT2”在發育后期高表達,則促進了TAG的高積累。(3)源基因“GPD1”和匯基因“DGAT1和DGAT2”的協同高表達在促進TAG合成底物G3P積累的同時加速acyl-CoA脂肪酸與DAG組裝成TAG。這不僅首次揭示了非種子組織(果肉)的油脂合成積累的源匯基因協同表達機制,而且為提高沙棘果肉含油量提供理論依據。
[1] Cenkowski S, Yakimishen R, Przybylski R,etal. Quality of extracted sea buckthorn seed and pulp oil[J]. Canadian Biosystems Engineering, 2006, 48(3): 9-16.
[2] Yang B R, Kallio H. Fatty acid composition of lipids in sea buckthorn (HippophaerhamnoidesL.) berries of different origins[J]. Journal of Agricultural and Food Chemistry, 2001, 49(4): 1939-1947.
[3] Ruan C J, Rumpunen K, Nybom H. Advances in improvement of quality and resistance in a multipurpose crop: sea buckthorn[J]. Critical Reviews in Biotechnology, 2013, 33(2): 126-144.
[4] Fatima T, Snyder C L, Schroeder W R,etal. Fatty acid composition of developing sea buckthorn (HippophaerhamnoidesL.) berry and the transcriptome of the mature seed[J]. Plos One, 2012, 7(4): e34099.
[5] Yang B R, Kallio H. Analysis of Triacylglycerols of seeds and berries of sea buckthorn (Hippophaerhamnoides) of different origins by mass spectrometry and tandem mass spectrometry[J]. Lipid, 2006, 41(4): 381-392.
[6] Ramli U S, Baker D S, Quant P A,etal. Use of control analysis to study the regulation of plant lipid biosynthesis[J]. Biochemical Society Transactions, 2002, 30(6): 1043-1046.
[7] Vigeolas H, Waldeck P, Zank T,etal. Increasing seed oil content in oil-seed rape (BrassicanapusL.) by over-expression of a yeast glycerol-3-phosphate dehydrogenase under the control of a seed-specific promoter[J]. Plant Biotechnology Journal, 2007, 5(3): 431-441.
[8] 張 霞, 張 樺, 張富春. 鹽脅迫下鹽穗木甘油醛-3-磷酸脫氫酶基因的表達與亞細胞定位分析[J]. 西北植物學報, 2015, 35(7): 1283-1288.
[9] Remize F, Barnavon L, Dequin S. Glycerol export and glycerol-3-phosphate dehydrogenase, but not glycerol phosphatase, are rate limiting for glycerol production inSaccharomycescerevisiae[J]. Metabolic Engineering, 2001, 3(4): 301-312.
[10] Vigeolas H, Geigenberger P. Increased levels of glycerol-3-phosphate lead to a stimulation of flux into triacylglycerol synthesis after supplying glycerol to developing seeds ofBrassicanapusL. in planta[J]. Planta, 2004, 219(5): 827-835.
[11] Guo H H, Wang T T, Li Q Q,etal. Two novel diacylglycerol acyltransferase genes fromXanthocerassorbifoliaare responsible for its seed oil content[J]. Gene, 2013, 527(1): 266-274.
[12] Zheng P, Allen W B, Roesler K,etal. A phenylalanine in DGAT is a key determinant of oil content and composition in maize[J]. Nature Genetics, 2008, 40(3): 367-372.
[13] Jako C, Kumar A, Wei Y D,etal. Seed-specific over-expression of anArabidopsiscDNA encoding a diacylglycerol acyltransferase enhances seed oil content and seed weight[J]. Plant Physiology, 2001, 126(2): 861-874.
[14] Cases S, Smith S J, Zheng Y,etal. Identification of a gene encoding an acyl CoA:diacylglycerol acyltransferase, a key enzyme in triacylglycerol synthesis[J]. Proceedings of National Academy of Sciences of the United States of America, 1998, 95(22): 13018-13023.
[15] Kroon J T M, Wei W X, Simon W J,etal. Identification and functional expression of a type 2 acyl-CoA: diacylglycerol acyltransferase (DGAT2) in developing castor bean seeds which has high homology to the major triglyceride biosynthetic enzyme of fungi and animals[J]. Phytochemistry, 2006, 67(23): 254-259.
[16] Li R, Yu K, Hildebrand D F.DGAT1,DGAT2 andPDATexpression in seeds and other tissues of epoxy and hydroxy fatty acid accumulating plants[J]. Lipids, 2010, 45(2): 145-157.
[17] Chen Y C, Cui Q Q, Xu Y J,etal. Effects of tung oilseed FAD2 and DGAT2 genes on unsaturated fatty acid accumulation inRhodotorulaglutinisandArabidopsisthaliana[J]. Molecular Genetics and Genomics, 2015, 290(4): 1605-1613.
[18] 阮成江, 李 群. 基因調控種子含油量研究進展及生物柴油用海濱錦葵遺傳改良策略[J]. 可再生能源, 2008, 26(4): 35-40.
[19] Vanhercke T, EI Tahchy A, Shrestha P,etal. Synergistic effect ofWRI1 andDGAT1 coexpression on triacylglycerol biosynthesis in plants[J]. FEBS Letters, 2013, 587(4): 364-369.
[20] Ding J, Ruan C J, Bao Y H,etal. Analysis of genetic relationships in sea buckthorn (Hippophaerhamnoides) germplasm from China and other countries using ISSR markers[J]. Journal of Horticultural Science & Biotechnology, 2015, 90(6): 599-606.
[21] Christie W W. Fatty acids and lipids: Structures, extraction and fractionation into classes[M]// Christie W W. Gas Chromatography and Lipids. Glasgow, UK: The Oily Press Ltd., 1989: 11-42.
[22] Vuorinen A L, Markkinen N, Kalpio M,etal. Effect of growth environment on the gene expression and lipids related to triacylglycerol biosynthesis in sea buckthorn (Hippophaerhamnoides) berries[J]. Food Research International, 2015, 77(3): 608-619.
[23] Wang X J, Liu A Z, Expression of genes controlling unsaturated fatty acids biosynthesis and oil deposition in developing seeds of Sacha Inchi (PlukenetiavolubilisL.)[J]. Lipids, 2014, 49(10): 1019-1031.
[24] Schmittgen T D, Livak K J. Analyzing real-time PCR data by the comparative CT method[J]. Nature Protocols, 2008, 3(6): 1101-1108.
[25] Xu R H, Wang R L, Liu A Z. Expression profiles of genes involved in fatty acid and triacylglycerol synthesis in developing seeds of Jatropha (JatrophacurcasL.)[J]. Biomass and Bioenergy, 2011, 35(5): 1683-1692.
[26] Chen G Q, Turner C, He X H,etal. Expression profiles of genes involved in fatty acid and triacylglycerol synthesis in castor bean (RicinuscommunisL.)[J]. Lipids, 2007, 42(3): 263-274.
[27] 吳永美, 毛 雪, 王書建, 等. 植物ω-7脂肪酸的系統代謝工程[J]. 植物學報, 2011, 46(5): 575-585.
CoordinatedExpressionofSourceandSinkGenesInvolvedinLipidBiosynthesisandAccumulationDuringSeabuckthornPulpDevelopment
DINGJian1,RUANCheng-jiang1,GUANYing2,GUANWen-ke3,SHANJin-you2,WUYu-xi2,WUTian-zhong3
(1.Institute of Plant Resources, Dalian Nationalities University, Dalian 116600, Liaoning, China; 2.Institute of Berries, Heilongjiang Academy of Agricultural Sciences, Suiling 152200, Heilongjiang, China; 3.Xinjiang Academy of Forestry Sciences, Urumqi 830064, Xinjiang, China)
ObjectiveThe objective of this study is to explore the relationship between lipid biosynthesis and source and sink genes’ expression in seabuckthorn (HippophaeL.) pulp.MethodTwo close-related trains ‘TF2-36’(with higher oil content) and ‘Za 56’ (with lower oil content) were selected as test samples. Their pulps were harvested in eight developmental stages. The oil content in pulp was tested by the method of chloroform methanol, and the differential expression of source gene ‘GPD1’ and sink genes ‘DGAT1 andDGAT2’ involved in lipid biosynthesis between high and low oil content lines were determined using qRT-PCR, and the effects of the three genes on lipid biosynthesis and accumulation were analyzed.Result(1) The oil contents in pulp of ‘TF2-36’ were higher than that of ‘Za 56’ at all stages, but it first increased, and then kept stable for two lines; (2) the peak values ofGPD1,DGAT1 andDGAT2 expression in pulp of TF2-36 were significantly higher than that in ‘Za 56’ during pulp development. The peaks ofGPD1 gene appeared in the period of rapid lipid biosynthesis, and the peaks ofDGAT1 andDGAT2 genes appeared in the period of stable lipid accumulation. The high expression of source gene (GPD1) contributed to synthesis more G3P of TAG precursor in early stages of pulp development, but the high expression of sink genes (DGAT1 andDGAT2) accelerated high TAG accumulation in later stages of pulp development.ConclusionThe high coordinated expression of source gene ‘GPD1’ and sink gene ‘DGAT1 andDGAT2’ resulted in the high lipid biosynthesis and accumulation in seabuckthorn pulp. These results provided basis for understanding lipid biosynthesis mechanism in seabuckthorn non-seed (pulp) tissue.
HippophaeL.; lipid biosynthesis; source gene; sink gene; gene expression
10.13275/j.cnki.lykxyj.2017.06.003
2016-09-02
國家自然科學基金(31570681)
丁 健(1983—),男,博士,講師,主要從事資源植物開發利用與遺傳育種.
* 通訊作者:阮成江(1972—),博士,教授,主要從事木本油料資源高效培育與利用研究。E-mail:ruan@dlnu.edu.cn
S793.6
A
1001-1498(2017)06-0902-06
張 研)
猜你喜歡
食品工業(2023年12期)2023-12-12 12:56:16
糧油食品科技(2022年6期)2022-11-25 12:18:34
糧油食品科技(2022年6期)2022-11-25 12:18:30
基層中醫藥(2020年8期)2020-11-16 00:55:20
環境衛生工程(2020年3期)2020-07-27 01:19:22
瘋狂英語·新讀寫(2020年3期)2020-06-06 09:06:14
塑料助劑(2019年3期)2019-07-24 08:51:22
中國糧油學報(2019年4期)2019-07-12 09:06:38
食品與機械(2018年11期)2019-01-03 06:15:34
中國糧油學報(2018年12期)2018-01-26 13:32:04
