冬蟲夏草寄主昆蟲及其飼養技術研究進展
2018-01-03 07:30:52李文佳張宗耀李全平呂延華蔣成吉魏再鴻
世界中醫藥 2017年12期
關鍵詞:研究
李文佳 張宗耀 李全平 呂延華 蔣成吉 魏再鴻
(1 宜昌山城水都冬蟲夏草有限公司,宜昌,443000; 2 廣東東陽光藥業有限公司國家中醫藥管理局重點研究室,東莞,523850)
冬蟲夏草寄主昆蟲及其飼養技術研究進展
李文佳1,2張宗耀1李全平1呂延華1蔣成吉1魏再鴻1
(1 宜昌山城水都冬蟲夏草有限公司,宜昌,443000; 2 廣東東陽光藥業有限公司國家中醫藥管理局重點研究室,東莞,523850)
冬蟲夏草(Ophiocordycepssinensis(Berk.)Sacc.)是一味傳統的名貴中藥材。蟲草蝠蛾(Hepialus)作為冬蟲夏草的寄主昆蟲,受到廣泛的關注和研究。本文從冬蟲夏草的寄主昆蟲種類、鑒定方法及地理分布,生物學特性和人工飼養等方面對蟲草蝠蛾的研究進展做了綜述,旨在為今后冬蟲夏草寄主昆蟲的研究和飼養提供借鑒和參考。
冬蟲夏草;蟲草蝠蛾;寄主昆蟲;飼養技術
冬蟲夏草(Ophiocordycepssinensis(Berk.)Sacc.)是一味傳統的名貴中藥材。蝙蝠蛾幼蟲被冬蟲夏草菌侵染后,冬蟲夏草菌吸收蟲體營養進行生長繁殖致使蟲體內長滿菌絲繼而形成子座,在適宜的環境下,子座生長出棒狀子實體。冬蟲夏草是傳統的滋補強壯藥,始載于《本草從新》,本品味甘性平,有益腎壯陽、補肺、止血化痰之效。人工培育冬蟲夏草的研究始于20世紀70年代末,其寄主昆蟲飼養是人工培養冬蟲夏草的關鍵環節[1]。
冬蟲夏草的寄主昆蟲是蟲草鉤蝠蛾ThitarodesarmoricanusOberthur[2]。蟲草鉤蝠蛾T.armoricanus,隸屬昆蟲綱,鱗翅目Lepidoptera,蝙蝠蛾科Hepialidae,蝠蛾屬Hepialus,國外學者則把蟲草鉤蝠蛾歸為鉤蝠蛾屬Thitarodes。蟲草鉤蝠蛾主要分布于我國甘肅、青海、新疆、四川、云南、西藏等地,幼蟲為多食性昆蟲,喜食蓼屬、黃芪屬等植物地下根莖,幼蟲期長,一般2~3年,最長的4~5年,營地下生活,耐寒性強,其余蟲態期較短,成蟲個體壽命僅幾天。
1 冬蟲夏草寄主昆蟲種類、鑒定及地理分布
1.1 寄主種類 中華人民共和國藥典(2010)[3]規定“冬蟲夏草為麥角菌科真菌冬蟲夏草菌OphiocordycepsSinensis(Berk.)Sacc.寄生在蝙蝠蛾科昆蟲幼蟲上的子座和幼蟲尸體的干燥復合體”。據此規定凡是蝙蝠蛾科昆蟲,只要和冬蟲夏草菌形成復合體,那么這一復合體就是冬蟲夏草。顯然冬蟲夏草的寄主昆蟲不止蟲草鉤蝠蛾T.armoricanus,目前普遍認為冬蟲夏草寄主昆蟲主要集中于蝠蛾屬Hepialus。
冬蟲夏草寄主種類一直在不斷豐富,最早朱弘復和王林瑤[4]歸納整理了9種冬蟲夏草的寄主昆蟲,楊大榮等[5]統計增至33種,隨后楊大榮等[6]再次統計冬蟲夏草寄主昆蟲至44種,Wang&Yao[7]進一步統計得出57種冬蟲夏草寄主昆蟲,邱乙等[8]重新歸納整理了61種寄主昆蟲但其中阿爾泰蝠蛾應不屬于冬蟲夏草寄主昆蟲。筆者整理了國內外發表的冬蟲夏草寄主昆蟲,發現國內分布的寄主昆蟲63種,國外11種其中不丹2種,尼泊爾8種,緬甸1種。見表1。
1.2 寄主鑒定 冬蟲夏草寄主的形態學分類鑒定主要采用雄性生殖器和成蟲翅脈特征,幼蟲分類作為種類鑒定的輔助在冬蟲夏草寄主鑒定中也有研究。余虹等[13]用斜脈蝠蛾幼蟲為研究對象通過各體節形態特征、毛序、顏色及頭寬體長等進行種類鑒別。嚴林[14]用拉脊蝠蛾幼蟲通過顏色、毛色、毛序及各齡幼蟲的頭寬和體長等進行分類研究。
分子生物學手段應用于蝙蝠蛾種類鑒定日益增多,線粒體DNA是種屬鑒定的常用手段,利用線粒體Cytb基因建立冬蟲夏草寄主昆蟲種屬間分子進化關系被證明具有可行性[11,15-16]。結合線粒體Cytb基因與線粒體COI,COII基因不僅能用于探索蟲草屬物種寄主昆蟲鑒定也用于區分蝠蛾屬昆蟲地理種群之間的差異[17-18]。

表1 冬蟲夏草寄主昆蟲分布概況

續表1 冬蟲夏草寄主昆蟲分布概況
注:寄主狀態:P代表潛在寄主;I代表不確定寄主
1.3 寄主地理分布格局 冬蟲夏草寄主主要分布于我國青藏高原及其周邊區域,包括我國的西藏、青海、甘肅、四川、云南等省,以及喜馬拉雅山南麓邊緣區的不丹、印度、尼泊爾等地,海拔在2 500~5 100 m之間[6]。最南分布在N25°45′、E99°34′;最北分布在N38°49′、E102°90′;最西在N29°53′,E89°32′;最東分布在N32°31′、E104°24′[19]。
冬蟲夏草的寄主昆蟲分布具有區域性。少數種群分布地域較廣,如蟲草鉤蝠蛾T.armoricanus廣泛分布于甘肅、青海、四川、云南、西藏等,其余種群多地域分布狹窄,如貢嘎鉤蝠蛾T.kangdingroides僅分布于貢嘎山系海拔3 000~5 000 m區域,小金鉤蝠蛾T.xiaojincusis主要分布于阿壩藏族自治州小金縣及接壤的金川縣和丹巴縣,蒲氏鉤蝠蛾T.pui分布于西藏色季拉山海拔4 100~4 650 m的高寒灌叢草甸和高寒草甸[20-23]。有些蝠蛾種類更是僅分布于一座山如德欽鉤蝠蛾T.deqingensis分布于德欽甲午雪山,白馬鉤蝠蛾T.baimaensis、銹色鉤蝠蛾T.ferrugineus分布于德欽白馬雪山,梅里鉤蝠蛾T.esmeiliensis分布于德欽梅里雪山,等等[24-25]。蟲草蝠蛾的分布不僅具有區域分布特點還具有垂直分布的規律,即同一種在不同緯度其垂直分布的范圍不一致,同一緯度不同的垂直高度可能分布著不同的種群。如蟲草鉤蝠蛾在四川、甘肅、西藏等地分布在海拔3 000~4 600 m之間,在青海為4 200~5 000 m,而在云南的分布幅度則在海拔3 850~5 080 m之間,異翅鉤蝠蛾T.anomopterus分布在劍川老君山西北坡海拔2 800~3 100 m之間,云南鉤蝠蛾T.yunnanensis分布在劍川老君山西北坡海拔3 600~4 100 m之間[8,26-28]。同一物種不同的海拔高度也可能產生不同的生態型,例如貢嘎蝠蛾在貢嘎山系海拔3 000~5 000 m范圍內分布著3種生態型[20],A型分布于海拔3 200~4 100 m,B型4 100~4 500 m,C型4 500 m以上分布,3種生態型在個體大小、繁殖活動時間、生活周期等方面均存在差異;小金蝠蛾也存在2種生態型[29],A型生活在海拔3 400~4 800 m,B型生活在海拔2 800~3 600 m,兩者在外部形態存在明顯差異且COI序列分析表明兩者相似度為99.2%以上,將兩類生態型相互交配發現后代可育且孵化率正常。
蟲草蝠蛾的分布易受地形、海拔、氣候、植被、土壤結構等因素的影響,尤其與土壤小氣候、食物、土壤特性和植物分布最為密切[19,26,30]。冬蟲夏草寄主幼蟲種群在自然環境條件下呈聚集分布狀態,同一塊蟲草分布塊中不存在2種以上的蟲草蝠蛾的現象,幼蟲的空間分布型符合二項式理論分布型[27,31]。
2 冬蟲夏草寄主昆蟲生物學研究
2.1 生物學特性 冬蟲夏草寄主蝠蛾為完全變態昆蟲,蝙蝠蛾屬不同種類昆蟲生物學特性雖然有微小差異,但它們完成一個世代所經歷的蟲態是基本一致的,在自然條件下,生活周期大概3~5年時間,分為卵、幼蟲、蛹、成蟲4個蟲態[32]。
卵:以小金蝠蛾為例,卵呈長圓形,大小0.8 mm×0.6 mm,卵的孵化有變色現象,由開始的白色慢慢變為黑色,全程約7 h,但其中也有部分發育不全不能孵化的卵不會發生變色過程,卵孵化為幼蟲約需要45~60 d[33];西藏那曲比如蝠蛾卵歷期則長達70 d,卵的孵化也有變色現象,由開始的乳白色慢慢變為黑色;云南劍川蝠蛾卵比小金蝠蛾和比如蝠蛾要輕,自然條件下,卵的孵化率都可以達到80%,高峰期為8月下旬和9月中旬[34]。
幼蟲:孵化時,幼蟲利用口器將卵殼咬破,然后爬出四處活動,各地幼蟲齡期分為6~8齡,初孵幼蟲乳白色,體長2 mm左右,老熟幼蟲頭殼淡紅色,蟲體姜黃色,體長3~5 cm左右,幼蟲孵化后陸續鉆入土壤縫隙,主要集中在15 cm左右深度的土層中。幼蟲期一般為2~3年時間,故在自然界,幼蟲存在世代重疊現象,幼蟲耐澇、耐寒、耐饑能力強[35]。
蛹:小金蝠蛾雌蛹肥大,尾部鈍圓,體長22.5~28.5 mm,寬5.2~5.8 mm。雄蛹瘦小,尾部尖削,體長16.4~18.8 mm,寬4.4~4.9 mm,自然條件下,5月上旬~7月上旬為化蛹高峰期;西藏比如蝠蛾雌蛹體長24.0~27.0 mm,寬5.5~6.0 mm。雄蛹體長19.6~22.0 mm,寬4.3~5.0 mm,化蛹高峰期為5月上旬至7月中旬;四川小金蝠蛾、西藏比如蝠蛾、云南劍川蝠蛾的蛹期都是40 d左右,而玉樹蝠蛾的蛹期能達到60 d[33,36-37]。
成蟲:成蟲不飲水、不取食,壽命較短。僅以羽化、交尾、產卵等生殖活動為中心,交尾高峰和產卵高峰都與具體生長海拔高度和當年氣候條件相關。四川小金蝠蛾成蟲平均成活105 h左右,最長為172 h,最短為60 h,成蟲在6月上旬開始羽化,6月中下旬為高峰期,8月中旬為末期,每只雌蛾最多產卵能達836粒,最少147粒,平均能產卵428粒;西藏比如蝠蛾成蟲每年從7月上旬開始羽化,高峰期在7月中下旬,末期在8月上旬,每只雌蛾平均產卵600粒;玉樹蝠蛾成蟲產卵最多為718粒,最少281粒,平均499.5粒[33,36-37]。
2.2 寄主食性與天然飼料
2.2.1 寄主食性 蝙蝠蛾幼蟲是多食性昆蟲[38]。陳泰魯等[39]最早發現蝙蝠蛾幼蟲長期取食珠芽蓼Polygonumviviparum能正常生長發育。沈發榮等[40]報道了白馬蝠蛾幼蟲能取食黃芪屬Astragalus、食蓼屬Polygonum等29種高原植物的根莖。冬蟲夏草寄主幼蟲平時以植物根莖為主,但在饑餓狀態下也會取食土中的腐殖質[32,41]。
2.2.2 天然飼料 冬蟲夏草寄主幼蟲可食用19科35屬的植物根莖部分(表2),喜食蓼屬、黃芪屬、大黃屬Rheum的植物,如蓼屬的圓穗蓼Polygonummacrophyllum、珠芽蓼、大黃屬的小大黃Rheumpumilum,這些植物多生長在西藏、四川、云南、青海等高原地帶,這與寄主昆蟲的分布是一致的,此外冬蟲夏草寄主幼蟲還喜食蘋果、胡蘿卜、馬鈴薯等[1,42]。
2.3 寄主昆蟲遺傳多樣性 地理隔離、寄主植物差異和環境脅迫等是造成其遺傳多樣性的重要因素,種群間無法實現基因交流,逐漸累積的不同的遺傳變異,達到一定程度會導致生殖隔離而形成不同物種[43]。國內已鑒定的冬蟲夏草寄主昆蟲種類達到60多種,造成如此之多的種群的原因在于冬蟲夏草寄主成蟲僅能生存3~8 d,且雌蛾懷卵量大、體態笨重難以長距離遷移,其自身擴散能力較弱;加之青藏高原地形地貌復雜,峰巒重疊、峽谷深有效地限制了寄主昆蟲種群間的擴散交流,因而造就了其豐富的遺傳多樣性[44]。這些因素在促成冬蟲夏草寄主昆蟲形成諸多種群的同時也造就出不同的生態型,這些生態型在形態外觀上存在極大差異但仍不能算作不同種群,例如小金蝠蛾有兩類生態型,一類為體表有褐色絨毛,翅脈上斑紋被褐色絨毛覆蓋,另一類為黑白斑紋,但經雄性生殖器鑒定和DNA序列鑒定為同一物種[29];斜脈蝠蛾在翅面斑紋和顏色上也可區分為“黑白斑”和“黃毛斑”兩大類[45]。棲息地地形、地貌、海拔、氣候、植被、土壤等眾多因素的差異,為寄主昆蟲提供其特有的種群形成和繁衍生存環境,由于冬蟲夏草寄主昆蟲諸多分布地人們尚未涉足,或將有更多的寄主昆蟲種類被發現[6]。
3 冬蟲夏草寄主昆蟲人工飼養進展

表2 蝙蝠蛾幼蟲喜食植物名錄
3.1 研究歷史 20世紀50年代末,國外多個國家進行過冬蟲夏草的人工栽培實驗研究,但均未獲得成功,從70年代末開始,國內有數十家科研院所開始對冬蟲夏草的人工培育進行研究。20世紀80年代末期重慶中藥研究所在四川康定海拔3 500 m的蟲草基地,成功在室內飼養出冬蟲夏草寄主斜脈蝠蛾,對斜脈蝠蛾整個生活史進行了詳細研究[46]。高祖訓等人在1991年將海拔3 500 m以上高原的斜脈蝠蛾引種到杭州低海拔實驗室,用人工飼料獲得良好效果[47]。進入21世紀以來,中山大學以及太極集團(與重慶中藥研究所合作)的半野生培植冬蟲夏草均取得一定進展,但都還僅僅停留在研發階段,無法突破產業化的瓶頸,直到2012年,東陽光集團生態撫育冬蟲夏草的量產才標志著冬蟲夏草的栽培獲得了長足的進步。
3.2 飼養種類 國內發現并鑒定與冬蟲夏草寄主有關的蝠蛾屬昆蟲60多種,其中進行過飼養研究的種類文獻統計有8種。見表3。
3.3 人工飼料 昆蟲人工飼料分為實用飼料、半純飼料和實全純飼料3大類[54]。蝙蝠蛾幼蟲的人工飼料是根據其營養要求和食性特點,采用合適材料經人為配制而成,主要是半純飼料和實用飼料2種。高祖紃等[47]在杭州低海拔實驗室,對當地22種植物進行了篩選試驗,選出3種植物作為斜脈蝠蛾的飼料,通過搭配飼料,選出了一組適宜斜脈蝠蛾取食的配制飼料。有文章指出,胡蘿卜、蘋果、馬鈴薯等是低海拔較好的實用飼料[48]。王宏生[55]將天然實用詞料胡蘿卜、小大黃與其他成分混合,輔以防腐劑,得出一組適合蝙蝠蛾生長的半純飼料,隨后四川農業大學和福建安發生物沿著這一思路繼續進行了一些探索[56-57]。
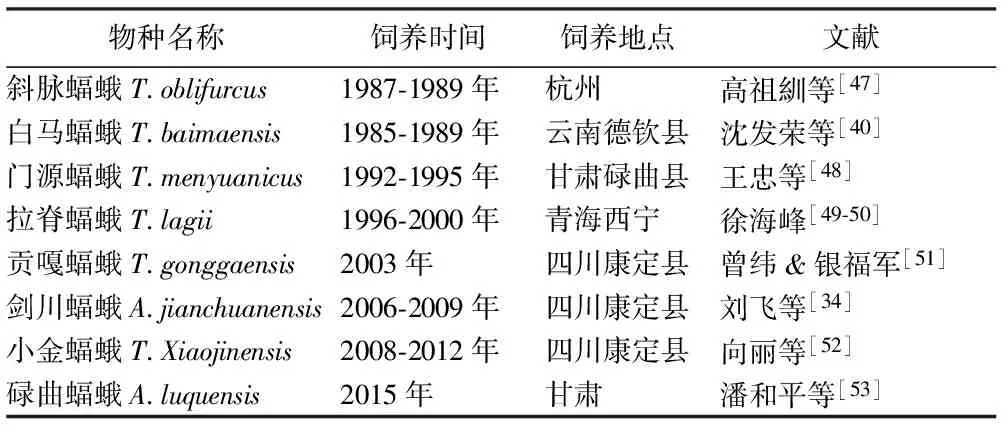
表3 蝠蛾屬昆蟲飼養種類
重慶中藥所[58-60]對冬蟲夏草寄主幼蟲飼料的營養成分進行了全面分析,發現在常規營養成分、維生素、礦質元素、氨基酸含量上均較高。四川農業大學對幼蟲的取食選擇性進行了詳細研究,得出幼蟲不僅對植物的種類取食存在偏好性,而且對同種植物不同部位也有一定的順序,同時對3種喜好植物塊根進行了營養成分分析,發現均含有較高的可溶性蛋白、粗脂肪和水分,同時礦質元素的含量也比較豐富[42]。而李文佳等[61]的研究證實,蝙蝠蛾幼蟲的取食選擇性與植物塊根中所含可溶性糖和粗蛋白存在著顯著的正相關。
3.4 飼養技術 蝙蝠蛾飼養包括卵、幼蟲、蛹、成蟲以及飼料和基質等,多年來科學家對蝙蝠蛾生長發育過程中所需要的光照、溫度、濕度以及飼養基質和飼養投放量進行了大量研究。
寄主卵的孵化方面,劉飛等[62]認為冬蟲夏草寄主卵孵化發育速率在濕度適宜的情況下主要受溫度制約,王忠等[48]研究表明卵孵化適宜溫度為12~16 ℃,濕度為85%~95%;陳仕江等[41]研究表明在10 ℃恒溫處理時孵化率最高,達91.4%,顯著高于野外變溫條件下86.8%的孵化率。寄主幼蟲飼養方面,王忠等[48]表明幼蟲飼養的溫度范圍應該控制在為6~12 ℃,相對濕度70%以上,該條件下幼蟲取食活躍,死亡率低;王宏生[55]則認為,幼蟲飼養的溫度范圍應該控制在為10~15 ℃,相對濕度在70%~80%內。蛹期羽化方面,王忠等[48]研究表明,蛹羽化的適宜條件與卵孵化一致,即最適溫度為12~16 ℃,濕度為85%~95%;劉飛等[62]也認為蛹期溫度應控制為12~16 ℃,但前25 d內的空氣相對濕度應控制在80%~90%內,后3~5 d應控制在90%~95%內,提供自然光照射。成蟲期,王忠等[48]提出交配箱置于溫度7~12 ℃,濕度范圍70%~85%的環境,黃昏時提供必需的散射光。
飼養基質和飼養投放量對幼蟲的成長有直接影響,基質對于初孵幼蟲成活率并無顯著影響,但飼料和卵的投放量對于初孵幼蟲的成活率有影響顯著[35]。
3.5 飼養裝置及集約化飼養 幼蟲期在生活史中所占周期最長,能否大規模飼養幼蟲是決定冬蟲夏草人工栽培是成功的關鍵。其中高祖訓等[47]將海拔3 500 m以上高原的斜脈蝠蛾引種到杭州低海拔實驗室,標志著人們探索飼養裝置進行集約化養蟲的開始,目前實驗室小規模一般是采用培養皿飼養幼蟲,雖然死亡率低,生長發育正常,由于耗時耗力無法做到大規模量產。實驗室中等規模飼養幼蟲涉及到養蟲盒的應用,李黎等[63]設計出了特制的蝙蝠蛾幼蟲養蟲盒,盒底鋪放40 mm滅菌基質,基質上擺放滅菌的天然飼料拌基質一層,然后投放50頭初孵幼蟲并蓋上與養蟲盒口大小相同的草皮,該方法相比培養皿在養蟲規模上有了重大突破,但養蟲盒養蟲容易造成幼蟲爭奪食物和地盤相互撕咬而死亡,并且容易發生交叉污染,使幼蟲大面積死亡,該方法用于擴大化生產存在系統性風險。另一種飼養方式是室外籠養,可在土壤疏松的野外挖深40 cm、長1 m、寬2 m的深槽、每平方米飼養密度為30~50頭[64],由于野外飼養幼蟲容易受到環境的影響,該方法至上世紀開展以來,一直沒有重大突破。現階段加緊研究集約化飼養幼蟲,解決蝙蝠蛾幼蟲的飼養工具,是該領域研究的方向,更是冬蟲夏草商品化成功的關鍵。
4 病蟲害
4.1 常見病蟲害 蟲草蝠蛾在野外生存率很低,造成死亡的因素很多[6],真菌、細菌、線蟲、寄生昆蟲、天敵捕食以及機械損傷都是造成幼蟲大批死亡的原因,其中病菌致死占幼蟲死亡率的50%,寄生昆蟲占15%,線蟲占20%,天敵捕食在10%,在人工飼養過程中幼蟲外逃也是造成大批死亡的誘因。造成蟲草蝠蛾死亡的常見真菌病害主要包括擬青霉和綠僵菌等。
4.1.1 細菌 染菌的蝠蛾幼蟲發病初期活動自如,約10 d左右,幼蟲蟲體體表顏色逐漸加深由黃白色至暗褐色、最后形成暗黑色。病害特點:蟲體不僵化,蟲體內部組織腐爛并發臭,對蟲體進行解剖學觀察可檢出細菌菌體[51,65]。
4.1.2 擬青霉 擬青霉曾被認作冬蟲夏草的無性型,隨著近些年的研究發現擬青霉是冬蟲夏草寄主蝠蛾的一種致病菌,陳建等[66]最早通過斜脈蝠蛾發現并鑒定該擬青霉病害為粉質擬青霉,也叫“黑點病”或“黑斑病”。蝙蝠蛾幼蟲18 ℃下接受侵染后3 d出現死亡癥狀,7 d死亡率達到90%。蟲體死后24 h內即可完全僵化,死后的幼蟲幾天后從尸體上長出白色菌絲,并逐漸生長形成分生孢梗束,同時產生大量白粉狀的分生孢子。孢梗束和白粉狀分生孢子是其典型病癥。該病為蝠蛾幼蟲的主要病害,長期引起幼蟲大面積死亡[51,65,67]。
4.1.3 綠僵菌 曾緯[68]研究表明,18 ℃下綠僵菌侵入幼蟲體內后3~4 d可引起80%的幼蟲死亡。發病的幼蟲尸體僵化,蟲體表面布滿白色氣生菌絲,菌絲上形成鏈生的卵圓形分生孢子,僵蟲體表顏色逐漸加深變綠,可產生綠孢子層,感染綠僵菌的幼蟲最典型的病癥是血淋巴里含有綠僵菌蟲菌體。
4.1.4 線蟲 被線蟲危害的幼蟲特點是死蟲不僵化,體表有大量線蟲,且尸體解剖可見有大量活的線蟲[51,65]。在野外線蟲感染致死率較低,但作者在室內飼養過程中發現由于幼蟲活動空間有限蝙蝠蛾幼蟲極易感染線蟲。
4.1.5 寄生昆蟲 寄生蜂和寄生蠅是野外常見的能夠寄生蝙蝠蛾的昆蟲[65]。寄生蠅寄生于幼蟲體內以幼蟲身體為營養基質,而寄生蜂可寄生于幼蟲體內直至蛹期。
4.1.6 機械損傷 蝠蛾幼蟲機械損傷較為常見,雖然不能造成直接死亡,但傷口增加了感病幾率導致死亡率升高且幼蟲受創后越冬成活的幾率遠低于健康幼蟲[69]。
4.2 病蟲害防治 最早在斜脈蝠蛾擬青霉病害中使用百菌清煙熏粉質擬青霉,能夠有效抑制孢子萌發[66]。紫外線直接照射擬青霉和綠僵菌也能有效抑制和殺滅孢子和菌絲體的生長,但非直射時效果較差且距離受限[70]。有人利用病蟲害綜合防治技術發現可以有效抑制主要病害和蟲害的發生[71]。
5 存在問題與展望
5.1 寄主種類與冬蟲夏草 寄主侵染研究是冬蟲夏草產業化的難題之一,在侵染時期、侵染方式、侵染機理、發育動態等方面取得眾多的成果[72-75]。冬蟲夏草寄主種類較多且在不斷豐富,眾多的種質資源為我們提供了數量繁多的研究樣本。不同產地的冬蟲夏草菌導致侵染力存在差異,不同產地冬蟲夏草菌與種類繁多的冬蟲夏草寄主昆蟲之間相互結合的時候,侵染力也是千差萬別[76]。因此篩選出優質的蟲草菌與優質的寄主,以及寄主與菌種間最優化的配置將是研究的熱點。
5.2 寄主昆蟲選育 李全平等[77]通過種質資源篩選獲得了一新種群,該新種對冬蟲夏草繁育環境的適應力強,發育整齊度高,種群擴繁倍數高,對中國被毛孢的敏感性較好,侵染率高,是目前少見的冬蟲夏草寄主昆蟲蟲種選育方面的報道。隨著人工培植冬蟲夏草的產業化,寄主昆蟲的良種選育工作也需全面開展。冬蟲夏草寄主昆蟲具有豐富的種質資源,育種工作的開展首先要系統評價各種蝠蛾種質大規模人工飼養的可行性、飼養難度及產出的冬蟲夏草的品質和質量,選出適宜冬蟲夏草產業化栽培的寄主種質資源;然后根據遺傳育種學的理論和技術,對表現優異的蟲種進行系統分離育種、雜交育種、抗病育種、誘變育種等,最終獲得寄主昆蟲優良品種,實現冬蟲夏草的寄主昆蟲標準化、規范化生產,滿足冬蟲夏草栽培高產、優質、高效的要求。
5.3 昆蟲飼養代時(生活史時間) 冬蟲夏草寄主昆蟲人工飼養條件下會出現世代周期縮短的現象,生活史由3~5年縮短為1~2年,斜脈蝠蛾、貢嘎蝠蛾、小金蝠蛾、拉脊蝠蛾均表現出這一特征[33,47,49,78-79],這很可能是人工飼養條件下食物充足、溫度穩定且適宜寄主昆蟲生長發育帶來有益效果。人工飼養條件下能更快達到其發育的有效積溫,從而完成生活史。寄主昆蟲的生活史長短對其大規模養殖和冬蟲夏草的產業化栽培成效有非常大的影響,應加強對冬蟲夏草各種寄主昆蟲人工飼養條件下的生長周期研究,從中選出更有利于生產的寄主昆蟲種類。
5.4 人工飼料及裝置 蝙蝠蛾幼蟲是一種多食性昆蟲,隨著冬蟲夏草培植產業化進行,蝙蝠蛾幼蟲取食的高原飼料需求將會大幅增加,對青藏高原的環境破壞將會更加嚴重。因此,采用人工飼料完全或部分取代天然飼料具有非常重要的意義,也是冬蟲夏草產業能否做大的關鍵。目前在人工飼料的研究上已經取得一定成果和經驗,但對長期取食人工飼料的蝙蝠蛾幼蟲是否存在退化風險、人工飼料保質期如何進一步延長等方面的研究還有待進行。另外為了滿足冬蟲夏草人工培植規模化、產業化生產,就必須使幼蟲飼養達到一定規模,而發明新的裝置,提高養蟲效率,是集約化養蟲需要攻克的難題。
5.5 病蟲害綠色防治 對于蟲草蝠蛾的病害防治目前仍無有效安全的防治藥劑,作為名貴中草藥冬蟲夏草的寄主昆蟲,蟲草蝠蛾在飼養過程中提倡綜合防治的方法無公害飼養。注重前期處理,即在室內飼養前對涉及的卵、水、飼料、土壤、飼養環境等做到全面消毒處理,盡量減少病原菌及寄生蟲帶入飼養環境。保證飼料供給、增強蟲體抗病性,蟲體感病后及時處理掉病死蟲。積極篩選抗病性高的種群,提倡雜交防止種群退化。
[1]張澤錦,葉萌,周祖基,等.冬蟲夏草寄主昆蟲飼養研究進展[J].中草藥,2009,39(40):85-87.
[2]朱弘復.冬蟲夏草的寄主昆蟲是蟲草蝙蝠蛾[J].昆蟲學報,1965,14(6):620-621.
[3]國家藥典委員會.中華人民共和國藥典[M].北京:中國醫藥科技出版社,2010:224-225.
[4]朱弘復,王林瑤.冬蟲夏草與蝙蝠蛾[J].動物學集刊,1985,2(3):121-134.
[5]楊大榮,李朝達,舒暢,等.中國蝠蛾屬昆蟲的種類和地理分布研究[J].昆蟲學報,1996,39(4):413-422.
[6]楊大榮,彭艷瓊,陳吉岳,等.中國冬蟲夏草寄主-蝠蛾屬昆蟲研究進展[C].云南省昆蟲學會2009年年會論文集,科學出版社,2009:291-301.
[7]Wang X L,Yao Y J.Host insect species of Ophiocordyceps sinensis:a review[J].ZooKeys,2011,(127):43-59.
[8]邱乙,程元柳,彭成,等.中國冬蟲夏草寄主昆蟲研究[J].時珍國藥醫藥,2015,26(3):720-722.
[9]Nielson E S,Robinson G S,Wagner A L.Ghost-moths of the world:a global inventory and bibliography of the Exoporia(Mnesarchaeoidea and Hepialoidea)(Lepidoptera)[J].Journal of Natural History,2000,34:823-878.
[10]Zou Z W,Liu X,Zang G R.Two new species of Thitarodes(Lepidoptera:Hepialidae)from Tibet in China[J].Pan-Pacific Entomologist,2011,87(2):106-113.
[11]鄒志文,劉昕,張古忍.中國蝠蛾屬(鱗翅目,蝙蝠蛾科)現行分類系統的修訂[J].湖南科技大學學報:自然科學版,2010,25(1):114-120.
[12]Maczey N,Dhendup K,Cannon P,etal.Thitarodes namnai sp.nov.and T.caligophilus sp.nov.(Lepidoptera:Hepialidae),hosts of the conomically important entomopathogenic fungus Ophiocordyceps sinensis in Bhutan[J].Zootaxa,2010,2412:42-52.
[13]余虹,高祖紃,陳健,等.斜脈蝠蛾幼蟲分類特征[J].昆蟲學報,1993,36(4):465-468.
[14]嚴林.青海拉脊蝠蛾幼蟲分類特征研究[J].青海大學學報:自然科學版,2001,19(1):5-9.
[15]陳永久,楊大榮,楊躍雄,等.青臧高原七個地區冬蟲夏草的分子進化研究[C].中國科協第三屆青年學術年會《材料科學與工程技術論文集》,北京,1998:87-89.
[16]程舟,耿楊,梁洪卉,等.用線粒體Cytb基因序列探討冬蟲夏草寄主蝠蛾的系統進化關系[J].自然科學進展,2007,17(8):1045-1052.
[17]Li S S,Chen H,Zhong X,etal.Study on CI I,Co II and Cytb Genes in the Identification of Hepialus Species[C].International Conference on Bioinformatics & Biomedical Engineering,2011,1-6.
[18]陳抒云,曹樹萍,袁航,等.線粒體COI和CYTB基因在蟲草屬物種寄主昆蟲鑒定中的適用性分析[J].世界科學技術-中醫藥現代化,2015,17(1):182-188.
[19]楊大榮,彭艷瓊,陳吉岳,等.中國冬蟲夏草分布格局與環境變化對其分布的影響[J].中國草地學報,2010,32(增刊):22-27.
[20]尹定華,李泉森,李黎,等.貢嘎蝠蛾生態分布的研究[J].特產研究,1994,44(1):6-9.
[21]朱弘復,王林瑤,韓紅香.中國動物志,昆蟲綱,第38卷,鱗翅目:蝙蝠蛾科、蛺蛾科[M].北京:科學出版社,2004:1-190.
[22]李峻鋒,鄒志文,劉昕,等.冬蟲夏草寄主蒲氏鉤蝠蛾的生物學[J].環境昆蟲學報,2011,33(2):195-202.
[23]涂永勤,朱華李,張德利,等.冬蟲夏草寄主-小金蝠蛾分布規律研究[J].西南農業學報,2013,26(5):2167-2169.
[24]梁醒財,楊大榮,沈發榮,等.云南蝠蛾屬Hepialus四新種(鱗翅目:蝙蝠蛾科)[J].動物學研究,1988,9(4):419-425.
[25]李朝達,楊大榮,沈發榮.蝠蛾屬一新種(鱗翅目:蝙蝠蛾科)[J].昆蟲學報,1993,36(4):495-496.
[26]楊大榮,龍勇誠,沈發榮,等.云南蟲草蝠蛾生態學的研究-Ⅰ.區域分布和生態地理分布[J].動物學研究,1987,7(1):1-11.
[27]劉禮平,龍綮新,周昌清.冬蟲夏草的寄主昆蟲-蟲草蝠蛾[J].昆蟲天敵,1995,17(4):184-190.
[28]劉飛,李兵,伍曉麗,等.從文獻計量分析國內外對冬蟲夏草的研究狀況[J].中國草地學報,2010,32(增刊):14-18.
[29]張德利,涂永勤.小金蝠蛾的生態型研究[J].環境昆蟲學報,2015,37(5):1055-1059.
[30]劉飛,伍曉麗,尹定華,等.冬蟲夏草寄主昆蟲的生物學研究概況[J].重慶中藥研究,2005,24(1):45-52.
[31]龍勇誠,楊大榮,沈發榮.蟲草蝠蛾幼蟲空間分布型的研究[J].動物學研究,1987,8(1):47-54.
[32]陸龍喜,劉淑梅,時連根.冬蟲夏草研究的新進展[J].生物學通報,2002,37(6):4-6.
[33]涂永勤,張德利,曾緯,等.四川小金蝠蛾生物學特性研究[J].應用昆蟲學報,2011,48(4):990-996.
[34]劉飛,伍曉麗,張德利,等.引種云南產冬蟲夏草寄主蝠蛾的生物學特性研究[J].中國中藥雜志,2009,34(4):379-381.
[35]劉飛,伍曉麗,羅慶明,等.冬蟲夏草寄主昆蟲的生物學特性研究[J].時珍國醫國藥,2008,19(12):2851-2852.
[36]王忠,馬啟龍,馬福全,等.玉樹蝠蛾生物學特性研究[J].甘肅農業科技,1995,32(12):38-40.
[37]尹定華,陳仕江,李黎,等.西藏冬蟲夏草寄主比如蝠蛾生物學特性的研究[J].特產研究,2004,54(2):1-5.
[38]黃天福,傅善全,羅慶明.康定蟲草蝙蝠蛾幼蟲食性試驗[J].四川動物,1989,8(3):8-10.
[39]陳泰魯,唐家駿,毛金龍.蟲草蝙蝠蛾Hepialus armoricanus Oberthür生物學的初步研究[J].昆蟲學報,1973,16(2):198-202.
[40]沈發榮,楊大榮,楊越雄,等.白馬蝙蝠幼蟲食性的觀察[J].昆蟲知識,1990,27(1):36-37.
[41]陳仕江,尹定華,丹增,等.中國西藏那曲冬蟲夏草的生態調查[J].西南農業大學學報,2001,23(4):289-292,296.
[42]陳一凱.蝙蝠蛾(Hepialus sp.)幼蟲對植物的取食選擇及營養成分測定[D].成都:四川農業大學,2011.
[43]劉全超,趙勇強,劉艷莊,等.昆蟲遺傳多樣性與種群分化的研究進展[J].河北林果研究,2011,26(2):184-187.
[44]張古忍,吳光國,古德祥,等.冬蟲夏草資源可持續利用面臨的問題及其對策[J].中國草地學報,2010,32(增刊):51-57.
[45]高祖紃,陳健,余虹,等.康定蟲草主要寄主斜脈蝠蛾的研究[J].昆蟲學報,1992,35(3):317-321.
[46]尹定華,唐雪梅.冬蟲夏草人工培殖研究的進展[J].中國中藥雜志,1995,20(12):707-710.
[47]高祖紃,陳健,余虹,等.全人工培養冬蟲夏草研究--康定蟲草寄主蝠蛾研究初報[J].浙江農業大學學報,1991,17(1):1-5.
[48]王忠,馬啟龍,喬正強,等.甘肅蟲草寄主昆蟲蝠蛾的全人工飼養技術[J].甘肅農業科技,2001,38(7):42-43.
[49]徐海峰.青海拉脊蝠蛾的生物學特性觀察研究[J].四川草原,2004,24(106):4-5.
[50]徐海峰.蝙蝠蛾多毛孢感染拉脊蝠蛾幼蟲的研究[J].四川草原,2005,25(114):15-16.
[51]曾緯,銀福軍.貢嘎蝠蛾幼蟲死蟲狀況調查[J].重慶中草藥研究,2003,45(2):5-6.
[52]向麗,陳士林,代勇,等.冬蟲夏草菌寄主小金蝠蛾人工飼養成蟲生物學特性[J].世界科學技術-中醫藥現代化,2012,14(1):1172-1176.
[53]潘和平,馬福全,王三喜,等.碌曲蝠蛾幼蟲的人工飼養方法[P].CN 104285902.2015.
[54]王延年.昆蟲人工飼料的發展、應用和前途[J].應用昆蟲學報,1990,27(5):310-312.
[55]王宏生.冬蟲夏草蝙蝠蛾人工飼養技術的初步研究[J].昆蟲知識,2002,39(2):144-146.
[56]李明超.冬蟲夏草寄主昆蟲小金蝙蝠蛾(Hepialus xiaojinensis Tu.)幼蟲天然飼料配方研究[D].成都:四川農業大學,2012.
[57]黃雪峰,王忠,鄭方毅,等.5種食料對蝙蝠蛾幼蟲生長的影響[J].武夷科學,2016,32(1):65-68.
[58]陳仕江,曾緯,尹定華.珠芽蓼根莖中的微量元素分析[J].特產研究,1994,44(3):61-62.
[59]劉飛,伍曉麗,錢敏,等.冬蟲夏草寄主幼蟲不同產地主要食料的營養成分比較分析[J].特產研究,2007,57(4):52-55.
[60]魯增輝,伍曉麗,劉飛.冬蟲夏草寄主幼蟲飼料營養成分比較分析[J].西南農業學報,2013,26(5):2048-2051.
[61]李文佳,李全平,魏再鴻,等.土壤和植物對冬蟲夏草寄主昆蟲規模化飼養的影響[J].菌物學報,2016,35(4):467-475.
[62]劉飛,伍曉麗,陳仕江,等.冬蟲夏草人工培殖的研究概況[J].中草藥,2007,38(2):302-305.
[63]李黎,陳仕江,小扎西,等.西藏那曲冬蟲夏草寄主昆蟲幼蟲飼養研究[J].重慶中草藥研究,2011,53(2):2-4.
[64]饒璐.冬蟲夏草寄主昆蟲的飼養[J].甘肅科技情報,1994,23(1):36.
[65]賀宗毅,劉飛,陳仕江,等.青藏高原特色資源冬蟲夏草培育過程中的病蟲害研究進展[J].重慶中草藥研究,2012,54(1):57-58.
[66]陳健,高祖紃,余虹,等.斜脈蝠蛾“黑點病”的病原及防治的初步研究[J].微生物學通報,1991,18(6):323-325.
[67]曾緯,陳仕江.冬蟲夏草寄主昆蟲擬青霉病研究[J].中國中藥雜志,2001,26(7):455-456.
[68]曾緯.貢嘎蝠蛾綠僵病菌的侵染、發生、傳播情況研究[J].重慶中草藥研究,2008,50(1):4-6.
[69]劉飛,張德利,曾緯,等.創傷對冬蟲夏草菌寄主幼蟲后續生長發育的影響[J].中國中藥雜志,2015,40(2):210-212.
[70]曾緯,錢敏,劉飛.紫外線照射對綠僵菌、擬青霉菌抑制或殺滅作用的研究[J].重慶中草藥研究,2006,48(2):1-3.
[71]劉飛,伍曉麗,曾緯,等.病蟲害綜合防治技術對冬蟲夏草寄主幼蟲生長發育的影響[J].中國中藥雜志,2008,33(23):2741-2744.
[72]曾緯,尹定華,李泉森,等.冬蟲夏草菌侵染及寄生階段的生長發育研究[J].菌物學報,2006,25(4):646-650.
[73]涂永勤,張德利,曾緯,等.蝠蛾屬幼蟲感染冬蟲夏草菌的實驗研究[J].食用菌,2010,29(3):16-17.
[74]馬少麗,張宗豪,徐海峰,等.拉脊蝠蛾幼蟲腸道菌群分析及侵染試驗[J].食用菌,2014,33(3):28-30.
[75]雷桅,彭青云,張古忍,等.冬蟲夏草菌在寄主鉤蝠蛾幼蟲中的潛伏侵染過程研究[J].環境昆蟲學報,2015,37(2):387-392.
[76]涂永勤,朱華李,張德利,等.不同產地冬蟲夏草菌對寄主幼蟲侵染力研究[J].中國食用菌,2012,31(5):32-34.
[77]李全平,賀媛,劉杰明,等.冬蟲夏草寄主昆蟲選育及生殖退化研究[J].菌物學報,2016,35(4):456-466.
[78]尹定華,李黎,黃天福,等.貢嘎蝠蛾各蟲態生態學的研究[J].特產研究,1994,44(4):5-7.
[79]陳仕江,黃天福,付善全.貢嘎蝠蛾生長發育的初步研究[J].中藥材,1995,18(4):171-173.
ResearchProgressofHostInsectofCordycepsanditsRaisingTechnique
Li Wenjia1,2, Zhang Zongyao1, Li Quanping1, Lyu Yanhua1, Jiang Chengji1, Wei Zaihong1
(1YichangShanchengShuiduCordycepsCo.,LTD,Yichang443000,China; 2SunShineLakePharmCo.,LTD,Dongguan523850,China)
Cordyceps is considered as a traditional rare Chinese materia medica. Hepialus, as the host insect of Cordyceps, is extensively focused on the scientific research. This paper summarized progress in research on Hepialus, including host insect species, identification method and geographical distribution for Cordyceps, their biological characteristics and manual feeding and so on, to provide references for further research and raising on the host insect of Cordyceps.
Cordyceps, Hepialus, Host insect, Raising technique
R282
A
10.3969/j.issn.1673-7202.2017.12.071
國家工信部中藥材提升和保障領域項目——冬蟲夏草規模化生產基地建設
李文佳(1982.11—),女,碩士,工程師,研究方向:中藥資源與開發,E-mail:LiWenjia@hec.cn
(2017-08-31收稿 責任編輯:徐穎)
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
