不同放養(yǎng)規(guī)格和放養(yǎng)密度下吉富羅非魚體長生長模型研究
2018-01-06 07:59:02陳倩蘇勝彥劉思辰葉偉朱偉凡袁新華
江蘇農(nóng)業(yè)科學(xué) 2017年22期
陳倩+蘇勝彥+劉思辰+葉偉+朱偉凡+袁新華


摘要: 為研究不同放養(yǎng)規(guī)格和養(yǎng)殖密度下吉富羅非魚的體長生長規(guī)律,采用von Bertalanffy、Gompertz、Logistic、Brody 4種生長模型對吉富羅非魚體長的生長進(jìn)行擬合,使用R-Studio軟件自行編程求出模型中各生長參數(shù),以AIC統(tǒng)計(jì)量作為確定吉富羅非魚體長最優(yōu)生長模型的準(zhǔn)則。結(jié)果表明,4種模型均能很好地模擬吉富羅非魚生長曲線,其中平均體長為0.8 cm (SL1)的試驗(yàn)組以Gompertz模型為最佳,平均體長為(2.8±0.14)cm (SL2)的試驗(yàn)組以Logistic模型為最佳。體長生長瞬時(shí)生長速度曲線均呈先上升再下降的趨勢,SL2試驗(yàn)組比SL1試驗(yàn)組更具養(yǎng)殖優(yōu)勢,養(yǎng)殖密度為15尾/m2(D1)的個(gè)體生長狀況優(yōu)于養(yǎng)殖密度為30尾/m2(D2)和60尾/m2(D3)的個(gè)體生長狀況。研究結(jié)果可為吉富羅非魚的育種研究、規(guī)模化養(yǎng)殖及養(yǎng)殖管理模式優(yōu)化提供理論依據(jù)。
關(guān)鍵詞: 吉富品系尼羅羅非魚;生長模型;AIC準(zhǔn)則;體長;規(guī)格;密度
中圖分類號: S965.125 文獻(xiàn)標(biāo)志碼: A
文章編號:1002-1302(2017)22-0170-06
羅非魚(Tilapia)屬慈鯛科非鯽屬,外形類似鯽魚,鰭條多刺,原產(chǎn)于非洲的坦噶尼喀湖,俗稱非洲鯽魚、非洲仔等。聯(lián)合國糧農(nóng)組織(FAO)在1976年向世界推廣養(yǎng)殖羅非魚,其作為物美價(jià)廉的經(jīng)濟(jì)魚類,快速獲得世界各國的認(rèn)可與養(yǎng)殖,也是中國主要的養(yǎng)殖魚類之一。吉富羅非魚具有生長速度快、養(yǎng)殖周期短、食性雜、抗病力強(qiáng)等優(yōu)點(diǎn),在廣東省、海南省、福建省、云南省和廣西壯族自治區(qū)這5個(gè)主產(chǎn)區(qū)實(shí)現(xiàn)了大規(guī)模養(yǎng)殖。范立民等在研究養(yǎng)殖密度對吉富羅非魚的生長性狀的影響時(shí)發(fā)現(xiàn),放養(yǎng)密度為27 000尾/hm2時(shí),吉富羅非魚的體長和體質(zhì)量要小于放養(yǎng)密度為18 000尾/hm2和22 500尾/hm2時(shí),高密度會(huì)對羅非魚的生長產(chǎn)生負(fù)影響[1]。朱佳杰等研究發(fā)現(xiàn),吉富羅非魚放養(yǎng)規(guī)格為2.5~2.7 cm的試驗(yàn)組比0.8~1.0 cm 和1.2~1.5 cm的試驗(yàn)組的畝產(chǎn)量、成魚規(guī)格、成活率高[2]。因此,研究不同放養(yǎng)規(guī)格和養(yǎng)殖密度下吉富羅非魚的生長情況,以量化模型研究其生長發(fā)育規(guī)律,對吉富羅非魚的養(yǎng)殖發(fā)展具有重要意義。
生長模型可以反映生物個(gè)體生長發(fā)育的規(guī)律性變化[3-4],魚類的生長曲線研究有助于掌握魚類的生長規(guī)律[5]。國內(nèi)對吉富羅非魚的生長模型研究僅限于同種規(guī)格和密度的研究,如唐章生等用4種生長模型對網(wǎng)箱單養(yǎng)吉富羅非魚的生長進(jìn)行了擬合,得出VB-GF生長模型對體質(zhì)量生長的擬合效果最好,而Gompertz GF生長模型對吉富羅非魚體長生長的擬合效果最好[6];肖俊等對尼羅羅非魚生長相關(guān)參數(shù)進(jìn)行生長模型構(gòu)建,對尼羅羅非魚的生長發(fā)育規(guī)律進(jìn)行了研究,得出Logistic、Gompertz、Bertalanffy這3種模型均能很好地模擬尼羅羅非魚生長曲線,其中以Logistic模型的擬合度最高[7]。而對于羅非魚在不同密度和放養(yǎng)規(guī)格的情況下的生長模型研究尚未見報(bào)道。本研究采用4種常用的生長模型擬合吉富羅非魚在2種放養(yǎng)規(guī)格和3種放養(yǎng)密度下的生長,揭示其體長生長規(guī)律,旨在為吉富羅非魚育種研究、養(yǎng)殖模式選擇和養(yǎng)殖生產(chǎn)經(jīng)濟(jì)性狀預(yù)測提供理論依據(jù)。
1 材料與方法
1.1 試驗(yàn)材料
1.1.1 試驗(yàn)用魚 供試吉富羅非魚由中國水產(chǎn)科學(xué)研究院淡水漁業(yè)研究中心屺亭科研實(shí)驗(yàn)基地提供。設(shè)置2種放養(yǎng)規(guī)格,每種分別為4 200尾,試驗(yàn)魚體長均值為0.8、(2.8±014)cm,用SL1、SL2表示。
1.1.2 試驗(yàn)池塘 試驗(yàn)地點(diǎn)設(shè)于中國水產(chǎn)科學(xué)研究院淡水漁業(yè)研究中心屺亭科研實(shí)驗(yàn)基地,試驗(yàn)池塘為12個(gè)長方形水泥池 (長×寬×高為10.0 m×2.0 m×1.5 m)。養(yǎng)殖用水為 地下水,水質(zhì)清新,養(yǎng)殖水深1.0 m,水溫變化范圍為24~33 ℃。微孔增氧,晝夜增氧,以保證氧氣充足。每日清除池塘內(nèi)殘餌和糞便,視情況進(jìn)行換水處理,每次換1/3水。
1.2 試驗(yàn)方法
1.2.1 密度設(shè)置 SL1和SL2均設(shè)置15尾/m2(D1)、30尾/m2(D2)、60尾/m2(D3)3種不同放養(yǎng)密度,每種放養(yǎng)密度設(shè)置2個(gè)水泥池作為對照。
1.2.2 飼喂 試驗(yàn)選用浙江寧波天邦股份有限公司生產(chǎn)的羅非魚粉料和膨化配合飼料(其中有0號羅非魚料、1號羅非魚料和2號羅非魚料)。前期投喂加水混好的羅非魚粉料,根據(jù)魚體的不同生長階段依次投喂0號羅非魚料、1號羅非魚料以及1號羅非魚料和2號羅非魚料的混合料。試驗(yàn)時(shí)間為2016年7月4日至2016年10月26日。采取飽食喂法,每日投飼量根據(jù)水溫和魚群吃食等具體情況調(diào)整。
1.2.3 數(shù)據(jù)測量 每周測量魚的體質(zhì)量、體長各1次,使用精確度為0.001 g的電子天平測定魚體質(zhì)量,使用精確度為0.02 mm的游標(biāo)卡尺進(jìn)行體長測量。每次從每個(gè)試驗(yàn)池中分別隨機(jī)取30尾進(jìn)行測量,測完放回,共進(jìn)行15次測量。
1.3 數(shù)據(jù)分析
1.3.1 模型選擇 本研究選取4種常用的生長模型來模擬吉富羅非魚體長的生長,各模型和擬合度見表1。
1.3.2 模型選擇準(zhǔn)則 本研究采用統(tǒng)計(jì)模型選擇中廣泛應(yīng)用的赤池信息量準(zhǔn)則(Akaikes information criteria,常簡稱AIC)作為選擇最佳模型的準(zhǔn)則[9-10]。
1.3.3 生長參數(shù)估計(jì)方法 使用R-Studio軟件自行編程求解方程,用Excel 2007軟件作折線圖。
2 結(jié)果與分析
2.1 不同規(guī)格和密度下體長生長曲線的分析
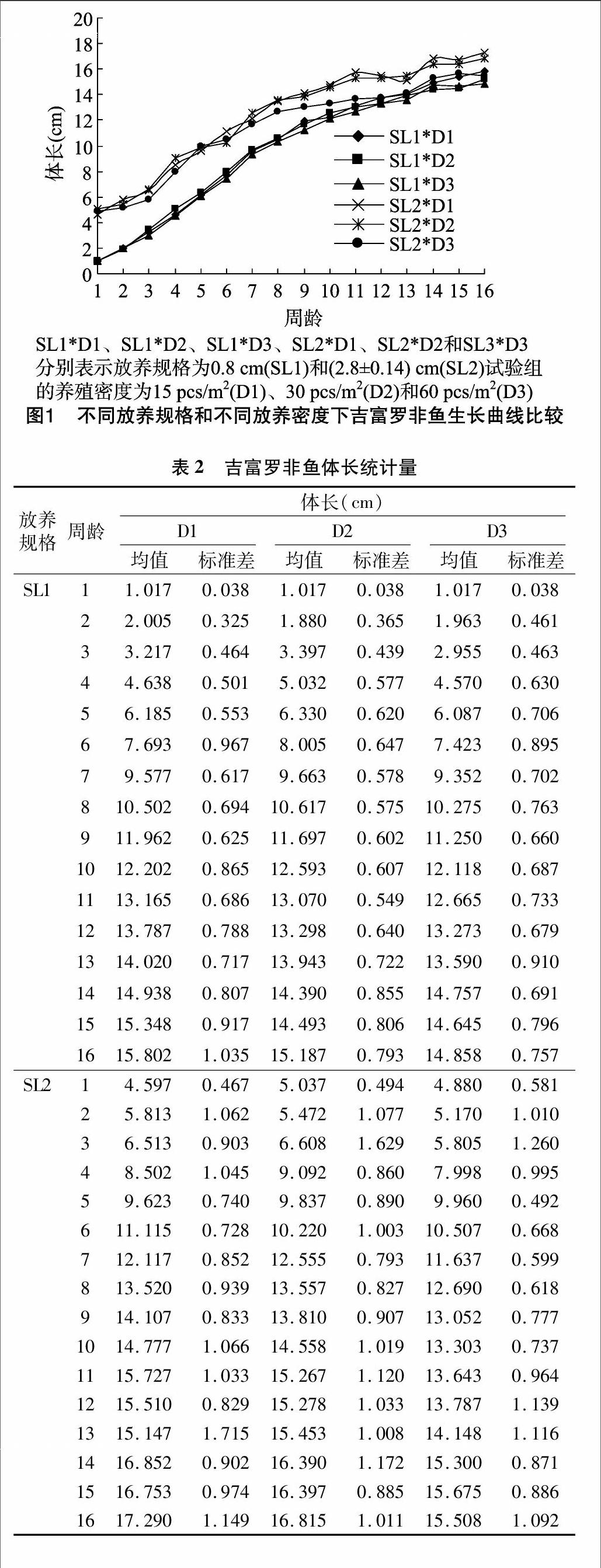
不同周齡吉富羅非魚SL1和SL2試驗(yàn)組的體長在D1、D2、D3這3個(gè)密度梯度下均呈不斷增長的趨勢(圖1)。SL1試驗(yàn)組養(yǎng)殖前期吉富羅非魚體長增長趨勢相同,從3周齡開始,D2的長勢明顯快于D3,但12周齡開始趨于相同,總體是D2快于D3。SL1試驗(yàn)組D1密度下的吉富羅非魚的體長增長速度介于D2與D3之間,但最后3周明顯快于D2和D3。SL1試驗(yàn)組3個(gè)養(yǎng)殖密度中,吉富羅非魚的總體體長增長速度是D1>D2>D3。SL2試驗(yàn)組中,從2周齡開始,D3密度下吉富羅非魚體長增長速度慢于D1和D2,從7周齡開始,明顯落后于D1和D2。從9周齡開始,D1密度下的吉富羅非魚體長增長速度快于D2密度下。SL2試驗(yàn)組中,吉富羅非魚的總體體長增長速度是D1>D2>D3。該研究中,SL2試驗(yàn)組是SL1試驗(yàn)組經(jīng)由約15 d養(yǎng)殖所得,后期吉富羅非魚SL2試驗(yàn)組的體長增長速度較SL1試驗(yàn)組明顯更快(圖1、表2),表明SL2試驗(yàn)組比SL1試驗(yàn)組更具養(yǎng)殖優(yōu)勢。endprint
2.2 體長性狀的模型擬合
由表3、表4可知,各模型的預(yù)測標(biāo)準(zhǔn)誤為0.003~0407,R2均大于0.922,方差分析(ANOVA)結(jié)果顯示各模型F值具有高度統(tǒng)計(jì)學(xué)意義(P<0.001),這表明各模型的擬合度都較好。
2.3 最優(yōu)模型的確定
由AIC準(zhǔn)則的統(tǒng)計(jì)值和R2可知,SL1試驗(yàn)組D1密度下von Bertalanffy模型和Gompertz模型的R2同為最大值 0.977,AIC值分別為 2 138.708 和2 142.999,相差甚微,但Gompertz模型擬合的體質(zhì)量值與觀察值更為符合,說明Gompertz模型擬合SL1試驗(yàn)組D1密度下的體長生長更合適。SL1試驗(yàn)組D2密度下Gompertz模型的AIC值最小,為 1 861.234,R2最大,為0.981,Gompertz模型擬合的體質(zhì)量值與觀察值比較符合,說明Gompertz模型擬合SL1試驗(yàn)組D2密度下的體長生長更合適;SL1試驗(yàn)組D3密度下Gompertz模型的AIC值最小,為 2 080.491,R2的值最大,為0.977,Gompertz模型擬合的體質(zhì)量值與觀察值比較符合,說明Gompertz模型擬合SL1試驗(yàn)組D3密度下的體長生長更合適(表3、圖2)。SL2試驗(yàn)組D1密度下,Logistic模型的AIC值最小,為2 812.770,R2的值最大,為0.938,Logistic模型擬合的體質(zhì)量值與觀察值比較符合,所以Logistic模型擬合SL2試驗(yàn)組D1密度下的體長生長更合適;SL2試驗(yàn)組D2密度下Logistic模型的AIC值最小,為 2 869.084,R2的值最大,為0929,Logistic模型擬合的體質(zhì)量值與觀察值比較符合,所以Logistic模型擬合SL2試驗(yàn)組D2密度下的體長生長更合適;SL2試驗(yàn)組D3密度下Gompertz模型和Logistic模型的AIC值分別為2 736.674和 2 742.390,相差較微小,Gompertz模型和Logistic模型的R2的值分別為0.926和0.925,相差也較微小,但Logistic模型擬合的體質(zhì)量值與觀察值更為符合,所以Logistic模型擬合SL2試驗(yàn)組D3密度下的體長生長更合適(表4、圖2)。
2.4 體長的絕對生長速度和生長拐點(diǎn)
為反映體長在其生長過程中某一時(shí)刻點(diǎn)的增長率,由模型的一階導(dǎo)數(shù)求得體長關(guān)于時(shí)間的瞬時(shí)生長速度[8,11-13]。由圖3可知,SL1試驗(yàn)組不同密度下的Gompertz模型和SL2試驗(yàn)組不同密度下的Logistic模型描述的吉富羅非魚體長瞬時(shí)增長速度變化趨勢大致相同,均為先上升后降低。SL1試驗(yàn)組不同密度下的Gompertz模型描述的曲線在4周齡同時(shí)達(dá)到峰值;SL2試驗(yàn)組不同密度下的Logistic模型描述的曲線變化趨勢不完全一致,其中D1和D2密度下模型描述的曲線在4周齡同時(shí)到達(dá)峰值,D3密度下模型描述的曲線在3周齡達(dá)到峰值。
整個(gè)生長過程中生長速率最大值點(diǎn)即是生長曲線的拐點(diǎn)。由表5可知,SL1試驗(yàn)組不同密度下的Gompertz模型的拐點(diǎn)分別出現(xiàn)在4.862、4.476、4.776周,拐點(diǎn)體長分別為6017、5.648、5.731 cm,還比較相近。SL2試驗(yàn)組不同密度下Logistic模型的拐點(diǎn)分別出現(xiàn)在4.220、4.026、3.800周,拐點(diǎn)體長分別為8.608、8.442、7.718 cm。SL2試驗(yàn)組D3密度下的拐點(diǎn)出現(xiàn)時(shí)間相對較早,拐點(diǎn)性狀值相對較小。
3 討論
3.1 不同放養(yǎng)規(guī)格和密度下的體長生長特征
體長是影響吉富羅非魚雄魚的重要性狀指標(biāo),并且相對體質(zhì)量指標(biāo)來說,體長指標(biāo)可以最大限度地減少環(huán)境的影響[13],所以本試驗(yàn)采用體長指標(biāo)討論吉富羅非魚的最優(yōu)生長模型。范立民等發(fā)現(xiàn)體長指標(biāo)在吉富羅非魚養(yǎng)殖中對體質(zhì)量的影響較大,尤其在低密度養(yǎng)殖中最顯著,因此可以利用吉富羅非魚的體長生長模型預(yù)測后期的體長和體質(zhì)量,達(dá)到對體質(zhì)量的早期選擇[1]。
該研究中SL2試驗(yàn)組是SL1試驗(yàn)組經(jīng)由約15 d養(yǎng)殖所得, 后期吉富羅非魚SL2試驗(yàn)組的體長增長速度較SL1試驗(yàn)
組明顯更快,且本試驗(yàn)結(jié)束時(shí)間為10月26日,此時(shí)氣溫開始下降,吉富羅非魚需轉(zhuǎn)移進(jìn)溫室進(jìn)行保溫養(yǎng)殖,若此時(shí)體長達(dá)到一定標(biāo)準(zhǔn),第2年溫度升高后,吉富羅非魚的體質(zhì)量較易增長,因此SL2試驗(yàn)組比SL1試驗(yàn)組更具養(yǎng)殖優(yōu)勢。吉富羅非魚總體體長增長速度是D1>D2>D3。
魚類在人工養(yǎng)殖過程中可能因?yàn)閾屖巢痪馐莛囸I脅迫[14-15],處于饑餓脅迫的魚類在恢復(fù)食物供給后會(huì)出現(xiàn)一段快速生長期,其生長速度快于同期非饑餓的魚類,這一現(xiàn)象稱為魚類的補(bǔ)償生長[16-17]。SL1試驗(yàn)組D3密度下的Gompertz模型描述的吉富羅非魚體長瞬時(shí)增長速度曲線后期高于D2密度下的曲線,可能是一種補(bǔ)償生長現(xiàn)象。
3.2 最適生長模型的選擇
一個(gè)完整的生物體生長過程可以用一條S形曲線表示,根據(jù)其生長速度的快慢,一般可以分為初始生長階段、指數(shù)生長階段和穩(wěn)定生長階段這3個(gè)階段,且具有“慢-快-慢”這樣的生長特征[8,11,18-22]。SL1和SL2試驗(yàn)組中,吉富羅非魚體長生長曲線經(jīng)過峰值后都是不斷下降,趨近于零,符合魚類生長的規(guī)律。雖然SL1和SL2試驗(yàn)組在生長過程中有一段生長是有重疊的,但我們發(fā)現(xiàn)不同規(guī)格適合不同的生長模型。
在動(dòng)物生長最優(yōu)模型的研究上,不同物種或同一物種的研究結(jié)果都不完全相同,可能受研究對象的生長環(huán)境和養(yǎng)殖模式影響[12,23-24],并且不同的生長發(fā)育階段也可能會(huì)呈現(xiàn)不同的生長規(guī)律[25-27]。同一魚類種苗不同,預(yù)測模型也可能不同[7]。Gompertz模型和Logistic模型在較多魚類體長研究中適用,唐章生等發(fā)現(xiàn)Gompertz GF生長模型對網(wǎng)箱養(yǎng)殖吉富羅非魚體長生長的擬合效果最好[6],何小燕等發(fā)現(xiàn)Logistic模型對大口黑鱸體長生長擬合效果最佳[24],郭媛等發(fā)現(xiàn)Gompertz生長模型對美洲紅點(diǎn)鮭(A)和白斑紅點(diǎn)鮭(B)及其雜交子代的體長生長曲線擬合效果最好[5]。Gompertz和Logistic模型不僅對吉富羅非魚體長擬合度高,并且曲線描述的體長瞬時(shí)增長速度變化趨勢和拐點(diǎn)時(shí)間也符合2種放養(yǎng)規(guī)格下的吉富羅非魚的體長生長規(guī)律,所以該研究認(rèn)為Gompertz和Logistic模型可分別作為SL1和SL2試驗(yàn)組下的最適體長生長模型。endprint
3.3 模型在吉富羅非魚養(yǎng)殖中的指導(dǎo)意義
在實(shí)際養(yǎng)殖生產(chǎn)中,可以根據(jù)建立的吉富羅非魚的生長模型對其生長情況進(jìn)行預(yù)測,根據(jù)不同生長階段的特點(diǎn)來確定吉富羅非魚的養(yǎng)殖生長潛力,優(yōu)化養(yǎng)殖管理模式,實(shí)現(xiàn)吉富羅非魚養(yǎng)殖經(jīng)濟(jì)效益最大化[12]。本研究對不同放養(yǎng)規(guī)格和養(yǎng)殖密度下的吉富羅非魚生長規(guī)律進(jìn)行探索,其生長模型的建立將有助于為養(yǎng)殖戶選擇放養(yǎng)規(guī)格和養(yǎng)殖密度提供重要參考;此外,也可以為不同規(guī)模的養(yǎng)殖戶在不同情況下選擇最佳的養(yǎng)殖模式提供理論指導(dǎo)。
參考文獻(xiàn):
[1] 范立民,Kamira B,宋 超,等. 不同養(yǎng)殖密度下吉富羅非魚生長性狀的通徑分析[J]. 中國農(nóng)學(xué)通報(bào),2015,31(11):83-87.
[2]朱佳杰,甘 西,謝爾登,等. 放養(yǎng)規(guī)格、養(yǎng)殖密度和水深對吉富羅非魚養(yǎng)殖效果的影響[J]. 水產(chǎn)科技情報(bào),2012,39(2):99-101.
[3]劉旭光,高 麗,趙益賢. 淮南麻黃雞生長曲線分析與擬合的研究[J]. 安徽農(nóng)業(yè)大學(xué)學(xué)報(bào),1997,24(4):362-365.
[4]李乃賓,杜炳旺,楊芬霞,等. 貴妃母雞生長模型擬合和分析的比較研究[J]. 家禽科學(xué),2015(2):9-11.
[5]郭 媛,戶 國,谷 偉,等. 美洲紅點(diǎn)鮭和白斑紅點(diǎn)鮭及其雜交子代幼魚的生長曲線擬合[J]. 中國水產(chǎn)科學(xué),2015,22(1):51-58.
[6]唐章生,林 勇,楊慧贊,等. 吉富羅非魚生長模型研究[J]. 廣東農(nóng)業(yè)科學(xué),2011(18):104-107.
[7]肖 俊,凌正寶,唐瞻楊,等. 尼羅羅非魚(Oreochromis niloticus)生長相關(guān)分析及生長模型構(gòu)建[J]. 海洋與湖沼,2012,43(6):1272-1278.
[8]符世偉,劉志剛,王 輝,等. 馬氏珠母貝殼長生長模型研究[J]. 廣東海洋大學(xué)學(xué)報(bào),2008,28(4):30-35.
[9]王 輝,郭 宏. AIC準(zhǔn)則應(yīng)用于動(dòng)物最佳生長模型的選擇[J]. 哲里木畜牧學(xué)院學(xué)報(bào),1998,8(1):34-38.
[10] 宋喜芳,李建平,胡希遠(yuǎn). 模型選擇信息量準(zhǔn)則AIC及其在方差分析中的應(yīng)用[J]. 西北農(nóng)林科技大學(xué)學(xué)報(bào),2009,37(2):88-92.
[11]王 輝,劉志剛,符世偉. 湛江北部灣海域養(yǎng)殖墨西哥灣扇貝重量性狀增長規(guī)律的研究[J]. 熱帶海洋學(xué)報(bào),2007,26(5):53-59.
[12]郭華陽,李有寧,張 楠,等. 企鵝珍珠貝早期養(yǎng)殖生長性狀的增長規(guī)律及生長曲線擬合研究[J]. 南方水產(chǎn)科學(xué),2016,12(5):71-80.
[13]唐瞻楊,林 勇,陳 忠,等. 尼羅羅非魚的形態(tài)性狀對體重影響效果的分析[J]. 大連海洋大學(xué)學(xué)報(bào),2010,25(5):428-433.
[14]Abolfathi M,Hajimoradloo A,Ghorbani R,et al. Compensatory growth in juvenile roach Rutilus caspicus:effect of starvation and re-feeding on growth and digestive surface area[J]. Journal of Fish Biology,2012,81(6):1880-1890.
[15]Velazquez M,Zamora S,Martinez F J. Influence of environmental conditions on demand-feeding behaviour of gilthead seabream (Sparus aurata)[J]. Journal of Applied Ichthyology,2004,20(6):536-541.
[16]Peres H,Santos S,Oliva-Teles A. Lack of compensatory growth response in gilthead seabream (Sparus aurata) juveniles following starvation and subsequent refeeding[J]. Aquaculture,2011,318(3/4):384-388.
[17]Ali M,Nicieza A,Wootton R J. Compensatory growth in fishes:a response to growth depression[J]. Fish and Fisheries,2003,4(2):147-190.[HJ1.66mm]
[18]劉志剛,王 輝,符世偉. 湛江北部灣養(yǎng)殖墨西哥灣扇貝的形態(tài)增長規(guī)律[J]. 水產(chǎn)學(xué)報(bào),2007,31(5):675-681.
[19]鄔思榮,劉志剛,王 輝. 湛江北部灣馬氏珠母貝紅殼色選育系生長模型研究[J]. 廣東海洋大學(xué)學(xué)報(bào),2011,31(3):16-24.
[20]Kuhi H D,Kebreab E,Lopez S,et al. An evaluation of different growth functions for describing the profile of live weight with time (age) in meat and egg strains of chicken[J]. Poultry Science,2003,82(10):1536-1543.
[21]Gamito S. Growth models and their use in ecological modelling:an application to a fish population[J]. Ecological Modelling,1998,113(1/2/3):83-94.
[22]邢黎峰,孫明高,王元軍. 生物生長的Richards模型[J]. 生物數(shù)學(xué)學(xué)報(bào),1998,13(3):348-353.
[23]頡曉勇,鐘金香,李思發(fā),等. 吉富品系尼羅羅非魚選育系F6、F7和F8當(dāng)年魚生長對比研究[J]. 南方水產(chǎn),2009,5(1):48-53.
[24]何小燕,白俊杰,樊佳佳,等. 大口黑鱸早期生長發(fā)育規(guī)律的研究[J]. 大連海洋大學(xué)學(xué)報(bào),2011,26(1):23-29.
[25]索效軍,張 年,熊 琪,等. 麻城黑山羊生長曲線的擬合與分析[J]. 江蘇農(nóng)業(yè)科學(xué),2015,43(11):291-293.
[26]Chen X B,Liu H B,Su Y P,et al. Morphological development and growth of the freshwater mussel Anodonta woodiana from early juvenile to adult[J]. Invertebrate Reproduction & Development,2015,59(3):131-140.
[27]Urban H J. Modeling growth of different developmental stages in bivalves[J]. Marine Ecology Progress Series,2002,238(1):109-114.endprint
