西藏尼木縣綿羊背帶屬線蟲形態(tài)及分子分類鑒定
2018-01-08 03:47:01夏晨陽劉建枝唐文強
西南農業(yè)學報 2017年12期
關鍵詞:研究
夏晨陽,劉建枝*,胡 敏,楊 新,石 斌,唐文強
(1.西藏自治區(qū)農牧科學院畜牧獸醫(yī)研究所,西藏 拉薩 850009;2.華中農業(yè)大學動物醫(yī)學院,湖北 武漢 430070)
西藏尼木縣綿羊背帶屬線蟲形態(tài)及分子分類鑒定
夏晨陽1,劉建枝1*,胡 敏2,楊 新2,石 斌1,唐文強1
(1.西藏自治區(qū)農牧科學院畜牧獸醫(yī)研究所,西藏 拉薩 850009;2.華中農業(yè)大學動物醫(yī)學院,湖北 武漢 430070)
【目的】2014年在西藏尼木縣綿羊皺胃收集到紅褐色線蟲830余條,對其進行形態(tài)及分子分類鑒定。【方法】首先進行了形態(tài)學鑒定,之后提取DNA,用特異性引物擴增ITS-2基因,并進行測序,與GenBank中的相關序列進行了同源性比較,建立了系統(tǒng)發(fā)育樹。【結果】西藏尼木縣綿羊體內感染的寄生蟲為背帶屬普通背帶線蟲Teladorsagiacircumcincta。【結論】首次在西藏發(fā)現(xiàn)背帶屬普通背帶線蟲Teladorsagiacircumcincta。家畜寄生蟲分子分類學研究今后將成為西藏寄生蟲病研究重要組成。
背帶屬;核糖體DNA;進化樹分析;西藏
【研究意義】寄生蟲命名、診斷、治療與控制對于寄生蟲基礎生物學與流行病學研究至關重要[1-2]。傳統(tǒng)的線蟲種類鑒定以形態(tài)學鑒定為主。分子生物學診斷技術是形態(tài)學鑒定的重要補充,寄生蟲蟲種鑒定已成功利用基因標記物(線粒體DNA、內部轉錄間隔區(qū)ITS、核糖體DNA等)開展線蟲分子分類學研究[3-6]。核糖體第二內轉錄間隔區(qū)(rDNA ITS-2)基因在核糖體DNA(rDNA)中是介于18S和28S之間的內轉錄間隔區(qū),在生物進化過程中,其進化速度快,種內具有高度保守性,而在不同種間又有不同程度的變異,因此為理想的種間鑒定遺傳標記[7]。【前人研究進展】腸道線蟲引起的家畜及人緩慢消瘦在全世界均有發(fā)生,是造成綿羊養(yǎng)殖業(yè)經(jīng)濟損失的主要寄生蟲[8]。腸道線蟲對綿羊養(yǎng)殖主要危害有食欲減退,體重減輕,毛、肉及乳產(chǎn)量減少,甚至導致死亡[9-12]。在寒溫帶地區(qū)綿羊主要寄生線蟲蟲種是圓線科背帶屬(Teladorsagia)線蟲[8,13],易感動物以瘦弱的羔羊為主[14]。奧斯特屬線蟲(Ostertagia)與背帶屬線蟲很難從形態(tài)上加以區(qū)分,背帶屬線蟲主要寄生于綿羊和山羊,而奧斯特屬線蟲主要寄生于牛[15]。蟲體長小于14 mm,褐色,口腔短小寬大,交合刺末端有2個或3個短小分支,常見于反芻動物的真胃內。雌蟲尾部末端具環(huán)紋,排卵器內的蟲卵是圓線科蟲卵代表,陰門具陰門蓋。背帶屬線蟲是溫帶[16-17]和許多國家[18-21]山羊、綿羊[22]重要的寄生線蟲,嚴重影響羊生產(chǎn)性能[16-17]。【本研究切入點】本文主要目的是研究西藏尼木縣綿羊體內采集到的背帶屬線蟲形態(tài)學及其分子分類學特性。
1 材料與方法
1.1 蟲體標本
線蟲標本2013年12月采自西藏尼木縣綿羊真胃,生理鹽水洗滌干凈后保存于盛有75 %酒精的20 mL EP管內,室溫保存。
1.2 主要試劑
DNA提取試劑盒WizardTMDNA Clean-Up System 為Promega公司產(chǎn)品;ExTaq、PCR緩沖液、MgCl2、dNTPs均為上海生工生物工程技術服務有限公司產(chǎn)品;其余試劑均由華中農業(yè)大學實驗室提供。
1.3 引物合成
參照國外文獻報道的序列引物[3-5,20],設計ITS-2上游引物NC1:5’-ACGTCTGGTTCAGGGTTGTT-3’;ITS-2下游引物NC2:5’-TTAGTTTCTTTTCCTCCGCT-3’,引物由生工生物工程(上海)股份有限公司合成。
1.4 蟲株DNA的提取
按照蟲株DNA提取試劑盒的操作步驟,將已鑒定的蟲株樣品用無菌水清洗干凈,除去表面黏液,取15條線蟲,在無菌條件下解剖取出內容物,剪碎,提取蟲株DNA,用瓊脂糖凝膠電泳鑒定后-20 ℃保存?zhèn)溆谩?/p>
1.5 目的基因片段的PCR擴增
分別以各分離株的DNA為模板,用上述引物進行PCR擴增。50 μl PCR反應體系:DNA模板1 μl,10×緩沖液(含Mg2+)5 μl, 2.5 mmol/L dNTPs 4 μl,引物各1 μl,TaqDNA聚合酶(5 U/μl)0.3 μl,最后加滅菌雙蒸水至總體積為50 μl。
擴增條件:94 ℃預變性5 min, 94 ℃變性30 s,54.3 ℃退火30 s,72 ℃延伸1 min,進行35個循環(huán),然后72 ℃再延伸7 min,4 ℃保存?zhèn)溆谩H∩鲜鯬CR產(chǎn)物5 μl在1.0 %的瓊脂糖凝膠(含0.5 μg/mL溴化乙錠)中,進行電泳分析,觀察擴增結果并拍照。
1.6 鑒定
擴增產(chǎn)物送上海生工生物工程技術服務有限公司測序,然后進行序列比較和分析。將測序所得的正確序列和從GenBank下載的ITS-2基因的相關序列(表1)進行分析,利用MEGA 5.05采用極大似然法(ML)和UPGMA 法構建進化樹。
2 結果與分析
2.1 形態(tài)學鑒定
蟲體線蟲形態(tài)學鑒定主要參考吳淑卿[23],張路平[24],黃兵[25]等人的文獻,逐條鑒定。如無特定說明長度單位為微米。
總體描述:普通背帶線蟲T.circumcincta活蟲卷曲,紅褐色。頸乳突顯著,食道-腸聯(lián)合顯著(圖1)。雄蟲:小,交合傘明顯。蟲體長(n=33)8578~11 373(9754.22±1011.21),蟲體最寬處(n=33)131~223 (163.28±26.69),食道長(n=31) 512~821 (680.03±93.03),食道膨大部(n=31) 48~67 (59.06±8.91),頸乳突距頭端長(n= 31) 271~456 (349.51±55.65)。交合傘對稱,分葉不明顯,具橫紋,背肋明顯,內含背輻肋,背肋60~72處分支。交合刺,細長,末端分三支,右支長(n=32) 273~349 (320.20±22.25),右交合刺分支處至末端長67~95 (86.06±9.61);左支長(n=32) 273~349 (324.00±21.93),右交合刺分支處至末端長67-95 (85.86±9.47);引器球拍狀,前部寬大,頭部尖細,后部絲狀,長(n= 23) 95~116 (100.23±9.14)。
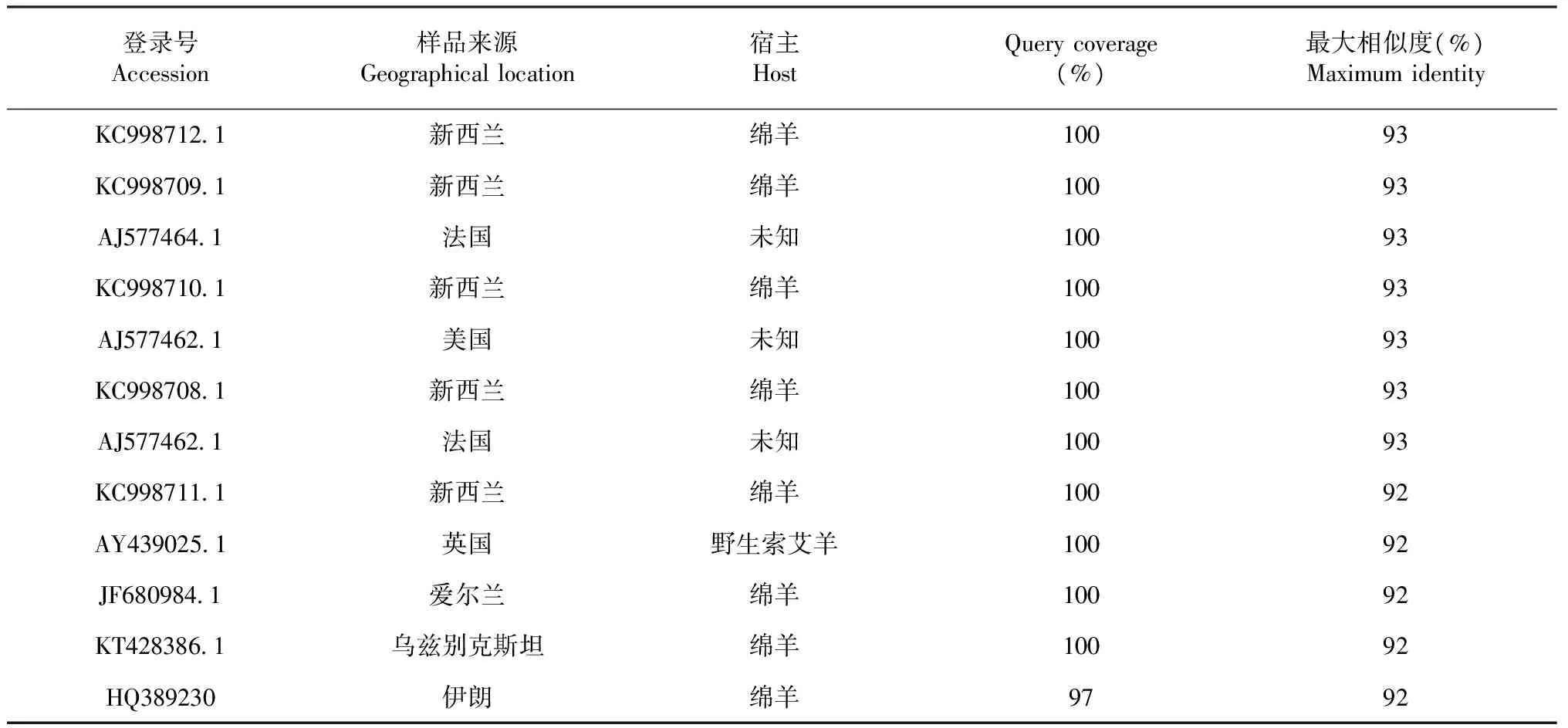
表1 普通背帶線蟲T. circumcincta同源性比較

1.頭部;2.頸乳突;3.肛門;4.陰門蓋;5.交合刺;6.引起;7.交合傘;8.背肋;9.交合傘橫紋1.Anterior and Cervical;2.Cervical papillae;3.Anus;4.Vulva;5.Spicule;6.Gubernaculum;7.Copulatory bursa; 8.Dorsal ray; 9.Copulatory bursa with cross striation圖1 普通背帶線蟲T. circumcincta形態(tài)特征Fig.1 Morphological character of T. circumcincta
雌蟲:小,線狀。蟲長:(n=31) 11518~15783 (13204.82±1791.82),蟲體最寬部位于陰門蓋附件;食道長(n=30) 640~916 (763.15±138.90),最寬部48~77 (66.02±12.66);頸乳突距頭端長(n=30) 349~475 (398.06±47.06)。
2.2 序列同源性比對
參照Genbank中已公布的普通背帶線蟲T.circumcincta序列及其他蟲株序列進行比對分析顯示,西藏普通背帶線蟲T.circumcincta樣本與其他地域樣本同源性最高達93 %(表1)。
2.3 進化樹的構建
以長刺后圓線蟲作為系統(tǒng)發(fā)育分析的外類群, 構建ML樹,各分支均進行1000次重復檢驗,其他對比線蟲包括艾克(氏)毛圓線蟲Trichostrongylusaxei(KF880746.1,KJ755059.1,伊朗), 細刺奧斯特線蟲Ostertagialeptospicularis(DQ354334.1,法國), 格氏奧斯特線蟲Ostertagiagruehneri(AJ400716.1, 挪威), 北方奧斯特線蟲Ostertagiaarctica(AJ250657.1, 挪威),捻轉血矛線蟲Haemonchuscontortus(KU558759.1,中國; FN432336.1, 意大利), 蛇形毛圓線蟲Trichostrongyluscolubriformis(HF 678107.1, 印度; KC337068.1, 老撾), 香柏馬歇爾線蟲Marshallagiaoccidentalis(AY013244.1, AJ400715.1,挪威),指形長刺線蟲Mecistocirrusdigitatus(AB114420.1, 日本; AJ577468.1, 法國),Teladorsagiacircumcincta(KC998712.1, 新西蘭), 長刺后圓線蟲Metastrongyluselongates(AJ305378.1,愛沙尼亞)。
進化樹結果顯示,西藏綿羊體內采集樣本與新西蘭發(fā)現(xiàn)的綿羊株(KC998712.1)同源性最高(圖2)。
3 討 論
3.1 背帶線蟲同源性
背帶屬線蟲世界上共有2種,我國寄生于綿羊和山羊體內主要是達氏背帶線蟲(Teladorsagiadavtiani),該寄生蟲呈世界性分布,澳大利亞、加拿大、美國、智利、挪威、英國、荷蘭,我國分布于內蒙古、新疆、甘肅[23]。2016年我國首次報道利用COI發(fā)現(xiàn)普通背帶線蟲Teladorsagiacircumcincta[26]。本研究首次利用ITS-2基因在青藏高原重要區(qū)域西藏發(fā)現(xiàn)普通背帶線蟲Teladorsagiacircumcincta。本研究以ITS-2作為遺傳標記,在分析序列變異的基礎上建立了區(qū)分背帶屬線蟲的PCR方法,從而為背帶屬線蟲分子生物學的進一步研究奠定了基礎,并為建立背帶屬線蟲的分子鑒別和快速診斷的PCR檢測方法提供了遺傳標記。

圖2 基于ITS-2基因序列的ML樹Fig.2 ML tree based on ITS2 gene sequences
3.2 分子技術在西藏寄生蟲研究中的應用前景
傳統(tǒng)形態(tài)學鑒定寄生蟲方法已無法適應現(xiàn)代寄生蟲分類研究,應采取形態(tài)與分子結合的方式對所在區(qū)域內的寄生蟲進行精確分類,這對于寄生蟲進化史研究具有重要的生物學意義,西藏家畜寄生蟲的分子分類研究今后必將成為家畜寄生蟲病研究領域重要組成。
以皮蠅蛆為例,通過分子技術手段證明中華皮蠅為一個獨立蟲種,不應歸類到紋皮蠅高山亞種,完全將其與紋皮蠅、牛皮蠅區(qū)分開來[27-31],筆者等人[32]也通過分子分類研究首次對西藏當雄牦牛體內采集的皮蠅樣本進行了分類研究,最終確定西藏當雄牦牛體內寄生的皮蠅蛆為牛皮蠅(Hypodermabovis)和中華皮蠅(Hypodermasinence),并且首次在西藏發(fā)現(xiàn)中華皮蠅。
4 結 論
通過分子生物學技術手段,首次在西藏發(fā)現(xiàn)背帶屬普通背帶線蟲Teladorsagiacircumcincta。家畜寄生蟲分子分類學研究今后將成為西藏寄生蟲病研究重要組成。通過不斷深入的調查研究,分子生物技術今后必將對西藏寄生蟲的分類研究產(chǎn)生深遠影響,取得重要的進展,寄生蟲蟲種的分類將更科學、更系統(tǒng)。
[1]Criscione C D, Poulin R, Blouin M S. Molecular ecology of parasites: elucidating ecological and microevolutionary processes[J]. Mol Ecol, 2005, 14: 2247-2257.
[2]Lichtenfels J R, Hoberg E P. The systematics of nematodes that cause ostertagiasis in domestic and wild ruminants in North America: an update and a key to species[J]. Vet Parasitol, 1993, 46:33-53.
[3]Stevenson L A, Gasser R B, Chilton N B. The ITS-2 rDNA ofTeladorsagiacircumcincta,T.trifurcataandT.davtiani(Nematoda: Trichostrongylidae) indicates that these taxa are one species[J]. Int J Parasitol, 1996, 26: 1123-1126.
[4]Hasheminasab S S. Molecular characterization of the first internal transcribed spacer of rDNA ofParabronemaskrjabinifor the first time in sheep[J]. Ann Parasitol, 2015, 61: 241-246.
[5]Holguin C M, Baeza J A, Mueller J D, et al. High genetic diversity and geographic subdivision of three lance nematode species (Hoplolaimusspp.) in the United States[J]. Ecol.Evol., 2015, 5: 2929-2944.
[6]Lott M J, Hose G C, Power M L. Parasitic nematode communities of the red kangaroo,Macropusrufus: richness and structuring in captive systems[J]. Parasitol Res., 2015, 114: 2925-2932.
[7]林瑞慶, 蔡天城, 吳桂英,等. 雞蛔蟲ITS rDNA的PCR擴增克隆及序列分析[J]. 中國獸醫(yī)雜志, 2008, 44(3):24-25.
[8]RV B. The economic impact of nematode infection in sheep: implications for future research and control[A]. Edited by Heath ACG. In The economic importance of parasites of livestock in New Zealand[C]. New Zealand: New Zealand Society for Parasitology, 1988: 4-16.
[9]Alberti E Z S, Ferrari N, Bruni G, et al. Effects of gastrointestinal nematodes on milk productivity in three dairy goat breeds[J]. Small Rumin Res, 2012, 106: S12-S17.
[10]Coop R L, Kyriazakis I. Influence of host nutrition on the development and consequences of nematode parasitism in ruminants[J]. Trends Parasitol, 2001, 17: 325-330.
[11]Roeber F, Jex A R, Gasser R B. Impact of gastrointestinal parasitic nematodes of sheep, and the role of advanced molecular tools for exploring epidemiology and drug resistance-an Australian perspective[J]. Parasites & Vectors, 2013(6):153.
[12]Zajac A M. Gastrointestinal nematodes of small ruminants: life cycle, anthelmintics, and diagnosis[J]. Vet Clin North Am Food Anim Pract, 2006, 22: 529-541.
[13]Urquhart G M, Armour J, Duncan J L, et al. Veterinary parasitology Second Edition[M]. Blackwell Science, Oxford, 2007:1-307.
[14]Wilkie H, Gossner A, Bishop S, et al. Variations in T Cell Transcription Factor Sequence and Expression Associated with Resistance to the Sheep NematodeTeladorsagiacircumcincta[J]. PloS one, 2016, 11: e0149644.
[15]Bowman D D. Georgis′ Parasitology for Veterinarians[M]. W.B. Saunders Company, 2014: 162-164.
[16]Morgan E R, Van D J. Climate and the epidemiology of gastrointestinal nematode infections of sheep in Europe[J]. Vet Parasitol, 2012, 189: 8-14.
[17]Stear M J, Bairden K, Bishop S C, et al. The processes influencing the distribution of parasitic nematodes among naturally infected lambs[J]. Parasitology, 1998, 117: 165-171.
[18]Barrett M, Jackson F, Huntley J F. Pathogenicity and immunogenicity of different isolates ofTeladorsagiacircumcincta[J]. Vet Parasitol, 1998,76: 95-104.
[19]Cabaret J, Morales G, Gruner L.Characterization ofTeladorsagiacircumcinctaandT.trifurcata. I. Epidemiological and biological aspects][J]. Ann Parasitol Hum Comp, 1984, 59: 607-617.
[20]Leignel V, Humbert J F. Mitochondrial DNA variation in benzimidazole-resistant and-susceptible populations of the small ruminant parasiteTeladorsagiacircumcincta[J]. J Hered, 2001, 92: 503-506.
[21]Stear M J, Henderson N G, Kerr A, et al. Eosinophilia as a marker of resistance toTeladorsagiacircumcinctain Scottish Blackface lambs[J]. Parasitology, 2002, 124: 553-560.
[22]Gasnier N, Cabaret J. Evidence for the existence of a sheep and a goat line ofTeladorsagiacircumcincta(Nematoda)[J]. Parasitol Res, 1996, 82: 546-550.
[23]吳淑卿. 中國動物志 線蟲綱 桿形目 圓線亞目(一)[M]. 北京:科學出版社,2001.
[24]張路平,孔繁瑤. 中國動物志 線蟲綱 桿形目 圓線亞目(二)[M]. 北京:科學出版社,2014.
[25]黃 兵,沈 杰. 中國畜禽寄生蟲形態(tài)分類圖譜[M]. 北京:中國農業(yè)科學技術出版社,2006:265-504.
[26]Lv J, Zhang Y, Feng C, et al. Species discrimination in the subfamily Ostertagiinae of Northern China: assessment of DNA barcode in a taxonomically challenging group[J]. Parasitol Res, 2016, 115: 987-996.
[27]Otranto D, Stevens J R. Molecular approaches to the study of myiasis-causing larvae[J]. Int J Parasitol, 2002, 32(11): 1345-1360.
[28]Otranto D, Colwell D D, Traversa D, et al. Species identification ofHypodermaaffecting domestic and wild ruminants by morphological and molecular characterization[J]. Med Vet Entomol, 2003, 17(3): 316-325.
[29]Otranto D, Traversa D, Guida B, et al. Molecular characterization of the mitochondrial cytochrome oxidase I gene of Oestridae larvae causing obligate myiasis[J]. Med Vet Entomol, 2003, 17(3): 307-315.
[30]Otranto D, Traversa D, Colwell D D, et al. A third species ofHypoderma(Diptera: Oestridae) affecting cattle and yaks in China: molecular and morphological evidence[J]. J Parasitol, 2004, 90(5): 958-965.
[31]Otranto D, Colwell D D, Pape T.Hypodermasinense: solving a century-old enigma[J]. Med Vet Entomol, 2005, 19 (3): 315-321.
[32]劉建枝,色 珠,關貴全,等. 西藏當雄牦牛皮蠅蛆病病原的分子分類鑒定[J]. 中國獸醫(yī)科學,2012,42(3):238-242.
MolecularandMorphologicallyClassificationofTeladorsagiafromSheepinNimuCountyofTibetAutonomousRegion
XIA Chen-yang1, LIU Jian-zhi1*, HU Min2, YANG Xin2, SHI Bin1, TANG Wen-qiang1
(1. Institute of Animal Science,Tibet Academy of Agricultural and Animal Husbandry Science,Tibet Lhasa 850009, China; 2. College of Veterinary Medicine, Huazhong Agricultural University, Hubei Wuhan 430070, China)
【Objective】830 reddish-brown nematodes were collected at Nimu county of Tibet in 2014 and morphologically characterized, and analysis molecular and morphologically classification ofTeladorsagia. 【Method】The genomic DNA of the nematode was extracted. Internal transcribed spacer 2(ITS-2)gene was amplified by specific primers sequenced and compared with that in GenBank. The phylogenetic tree based on the ITS-2 gene was constructed. 【Result】The specimens from the county wereTeladorsagiacircumcincta, for the first time founded in Tibet. 【Conclusion】The molecular classification of parasites from livestock in Tibet will be important part of study of Tibet parasitic diseases in future.
Teladorsagia; Nuclear ribosomal DNA; Phylogenetic analysis; Tibet
1001-4829(2017)12-2838-05
10.16213/j.cnki.scjas.2017.12.038
2017-02-12
公益性行業(yè)(農業(yè))科研專項(201303037)
夏晨陽(1980-),男,滿族,黑龍江明水人,副研究員,研究方向為西藏家畜寄生蟲病防控,E-mail:13518978860@163.com。*為通訊作者:劉建枝,研究員,主要從事家畜寄生蟲病防控研究,Tel:13659526886,E-mail:13659526886@163.com。
S432.4+5
A
(責任編輯李 潔)
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
