黃瓜S期激酶相關(guān)蛋白Skp1的原核表達及其多克隆抗體的制備
2018-01-12 00:42:22孫新艷韓曉玉王振躍陳琳琳燕照玲艷1
華北農(nóng)學報 2017年6期
關(guān)鍵詞:分析
孫新艷,魏 瑩,韓曉玉,王振躍,陳琳琳,燕照玲,施 艷1,
(1.河南省果樹瓜類生物學重點實驗室,河南 鄭州 450002;2.河南農(nóng)業(yè)大學 植物保護學院,河南 鄭州 450002;3.河南省農(nóng)業(yè)科學院,河南 鄭州 450002)
S期激酶相關(guān)蛋白1 (S-phase kinase-associated protein 1,Skpl ) 是真核生物中普遍存在的一類蛋白,是SCF型E3泛素連接酶途徑的核心蛋白,具有特異性地連接Cullin1(Cul1) 和F-box蛋白的作用[1-3],通過與Cullin1、Rbx1和F-box蛋白形成SCF復合物來發(fā)揮功能,在SCF復合物中,Cullin1與Rbx1結(jié)合,F(xiàn)-box蛋白識別靶標蛋白,Skp1分別結(jié)合F-box蛋白和Cullin1,Skp1-Cul1-F-box (SCF) E3復合體在多個細胞過程中起重要作用[4]。研究表明,Skp1在真核生物的細胞周期、轉(zhuǎn)錄調(diào)控、信號傳導等很多細胞進程中發(fā)揮關(guān)鍵作用。Skp1相關(guān)基因被證實在植物生長素、赤霉素、乙烯、茉莉酸和光反應途徑中有十分重要的作用[5-11]。研究發(fā)現(xiàn),Skp1基因在小麥從可育轉(zhuǎn)變?yōu)椴挥^程中表達受到抑制,說明Skp1與小麥育性轉(zhuǎn)變密切相關(guān)[12]。本試驗構(gòu)建了黃瓜Skp1基因的原核表達載體,并在大腸桿菌中高效表達出該基因的融合蛋白,通過免疫新西蘭大耳白兔制備了特異性抗血清,為進一步研究該基因的功能奠定了基礎(chǔ)。
1 材料和方法
1.1 試驗材料、菌株、質(zhì)粒和試劑
供試黃瓜品種為新優(yōu)36,種植于溫室中。
大腸桿菌菌株DH5α、BL21、原核表達載體pET-28a由河南省果樹瓜類生物學重點實驗室保存。
限制性內(nèi)切酶、T4DNA 連接酶購自美國NEB公司。PrimeSTAR Max DNA Polymerase購自TaKaRa。IPTG、Kana、Tris、SDS、丙烯酰胺、甘氨酸為北京索萊寶科技有限公司產(chǎn)品。辣根過氧化物酶標記的羊抗兔IgG為Sigma 產(chǎn)品。SanPrep柱式質(zhì)粒DNA小量抽提試劑盒為生工生物工程上海股份有限公司產(chǎn)品。其他試劑為進口和國產(chǎn)分析純。
1.2 引物設(shè)計
根據(jù) NCBI 已經(jīng)發(fā)表的黃瓜Skp1基因序列,利用軟件DNAMAN設(shè)計擴增全長閱讀框的引物。正向引物為5′-GACGGATCCATGTCCTCCTCCAACAAA
AT-3′,反向引物為5′-GATCTCGAGTCATTCACAAGCC
CACTGAT-3′。引物由南京金斯瑞生物科技有限公司合成。為了便于克隆,分別在5′ 和3′ 端添加酶切位點BamH Ⅰ和XhoⅠ。
1.3 Skp1基因原核表達載體的構(gòu)建
以黃瓜葉片cDNA為模板擴增Skp1基因,PCR 50 μL擴增體系:1 μL cDNA,正向引物和反向引物各0.5 μL,PrimeSTAR Max DNA聚合酶25 μL,ddH2O 23 μL。PCR反應條件:94 ℃ 預變性5 min;94 ℃ 30 s,56 ℃ 30 s,72 ℃ 40 s,35個循環(huán);72 ℃ 延伸10 min。PCR產(chǎn)物通過試劑盒回收純化。將純化后的Skp1片段和pET-28a載體分別用BamHⅠ 和XhoⅠ雙酶切,用T4DNA 連接酶連接過夜后轉(zhuǎn)化大腸桿菌DH5α,挑取陽性克隆進行菌落PCR鑒定并測序,提取質(zhì)粒。
1.4 Skp1基因的誘導表達和SDS-PAGE 分析
將重組質(zhì)粒轉(zhuǎn)化大腸桿菌BL21菌株,挑取單菌落于37 ℃過夜培養(yǎng),用含Kana 50 μg/mL 的LB培養(yǎng)基按1∶100的比例擴大培養(yǎng),振蕩培養(yǎng)至OD600為0.8。加入IPTG 至終濃度為1 mmol/L,繼續(xù)培養(yǎng)3,6,9 h,分別離心收集菌體,每50 mL菌液加入5 mL ddH2O和1 mL 樣品緩沖液(40 mmol/L Tris-HCI(pH值6.8)、10% 甘油、5% SDS、5% 巰基乙醇、0.1%溴酚藍),煮沸10 min,離心后取上清,用12% 的分離膠進行SDS-PAGE分析。
1.5 抗血清制備及其效價測定
參照Hager 和Burgess的方法回收表達產(chǎn)物,表達產(chǎn)物經(jīng)12%的SDS-PAGE 電泳后,將凝膠用預冷的0.25 mmol/L KCl,1 mmol/L DTT 溶液浸泡5 min,重蒸水沖洗,切下含目的條帶的凝膠,按1∶1比例(m/V)加入PBS 緩沖液(0.14 mol/L NaCl、2.7 mmol/L KCl、1.5 mmol/L KH2PO4、8.1 mmol/L Na2HPO4),于冰浴中研磨[13]。用PBS緩沖液適當稀釋后,加入等體積的福氏不完全佐劑(1.5 g羊毛脂+8 mL石蠟油)進行乳化。采用肌肉注射的方法免疫新西蘭大耳白兔,共免疫5次。
以純化的蛋白(2.5 μg/mL) 作為抗原包被ELISA板,將抗血清進行一系列倍比稀釋后作為一抗,辣根過氧化物酶標記的羊抗兔IgG為二抗,用ACP-ELISA測定抗血清的效價。
1.6 蛋白免疫印跡(Western Blot)分析
利用植物總蛋白抽提緩沖液 (220 mmol/L Tris-HCl,pH值 7.4,250 mmol/L蔗糖,1 mmol/L MgCl2,50 mmol/L KCl,10 mmol/L β-巰基乙醇,100 μmol/L PMSF)提取黃瓜葉片總蛋白,取10 μL 上樣于5% 的濃縮膠和12% 的分離膠進行SDS-PAGE 電泳,電泳結(jié)束后轉(zhuǎn)移到PVDF膜上,加稀釋500倍和稀釋1 000倍的兔抗血清雜交,隨后加入辣根過氧化物酶標記的羊抗兔IgG(1∶2 000稀釋),在增強型HRP-DAB 底物顯色試劑盒中顯色至條帶清晰。
2 結(jié)果與分析
2.1 黃瓜Skp1基因原核表達載體的構(gòu)建
以黃瓜葉片cDNA為模板,經(jīng)PCR擴增得到1條特異性片段(圖1),與預期大小一致。擴增出的特異性條帶在T4DNA連接酶的作用下連接到pET-28a載體上,菌落PCR驗證陽性克隆并測序,獲得重組質(zhì)粒pETSkp1。Skp1基因全長閱讀框由468個核苷酸組成,共編碼155個氨基酸。
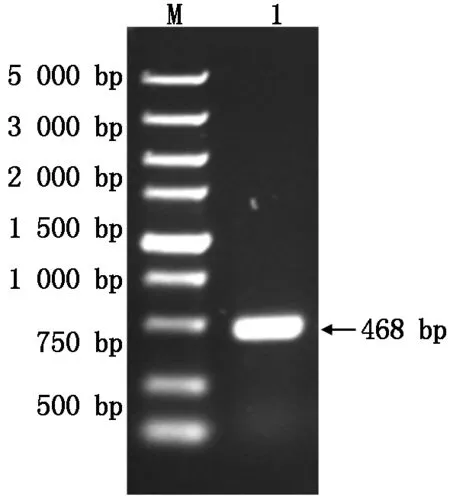
M.DNA 分子標準DL5000;1.PCR 產(chǎn)物。M.DNA Marker DL5000;1.PCR Product.
2.2 表達產(chǎn)物的SDS-PAGE 分析和蛋白純化
將誘導后的含有pETSkp1的重組質(zhì)粒的菌株裂解物進行SDS-PAGE分析,未誘導的菌株裂解物作為對照,從圖2可以看出,誘導后的菌株可產(chǎn)生分子質(zhì)量約20 kDa 的特異蛋白質(zhì)條帶,與預期大小相符。IPTG誘導后3 h,Skp1蛋白即獲得誘導表達,6,9 h誘導對表達量沒有影響,因此,本試驗用3 h進行誘導,切膠回收蛋白,研磨純化。
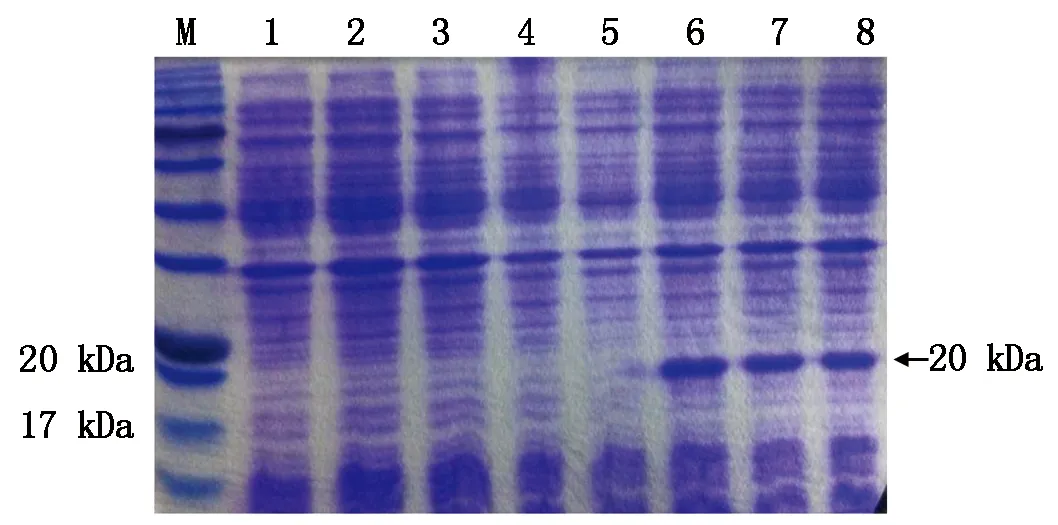
M.蛋白分子質(zhì)量標準;1.pET空載體未誘導;2~4.未加IPTG,pETSkp1 誘導3,6,9 h;5.pETSkp1未誘導;6~8.IPTG誘導pETSkp1后3,6,9 h。
M.Protein Marker;1.pET-28a without induction;2-4.Total proteins of pETSkp1 being induced for 3,6,9 h respectiveIy without IPTG;5.pETSkp1 without induction;6-8.Total proteins of pETSkplbeing induced for 3,6,9 h respectively.
圖2SDS-PAGE分析Skp1融合蛋白的誘導表達
Fig.2SDS-PAGEanalysisofexpressionproductsofpETSkp1
2.3 抗血清的制備及效價測定
用回收的特異性蛋白免疫新西蘭大白兔,5次免疫后取血獲得黃瓜Skp1的抗血清。將純化的蛋白作為抗原,分別通過2種方式進行抗原包被,ACP-ELISA 檢測結(jié)果表明:用膠條電洗脫包被,抗血清稀釋128×103倍后能明顯檢測出;用包涵體抗原包被,抗血清稀釋512 × 103倍后能明顯檢測出(表1)。

表1 抗血清效價測定Tab.1 Determination of the titer of antiserum
注:+.陽性;-.陰性。
Note:+.Positive;-.Negative.
2.4 Western Blot 檢測黃瓜樣品結(jié)果
提取黃瓜葉片的總蛋白,將制備的抗血清分別稀釋500,1 000倍作為一抗來檢測抗血清的特異性。結(jié)果表明,在這2種條件下均能明顯地檢測出Skp1編碼的蛋白,并與預期結(jié)果一致(圖3)。
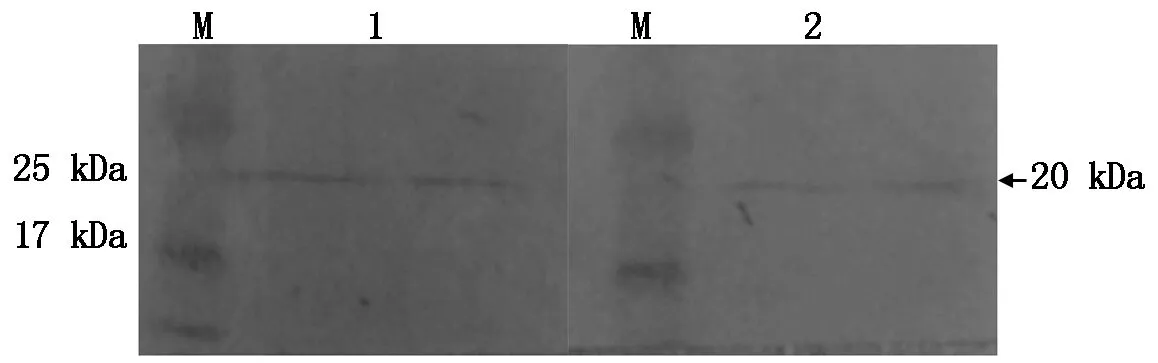
M.蛋白分子質(zhì)量標準;1.抗血清稀釋500倍;2.抗血清稀釋1 000倍。 M.Protein Marker;1.Antiserum diluted 500 times;2.Antiserum diluted 1 000 times.
3 結(jié)論與討論
本研究確定了Skp1蛋白在大腸桿菌BL21中高效表達的條件,利用純化的蛋白制備了抗血清,并通過ELISA測定了其效價為1∶128 000~512 000,Western Blot檢測結(jié)果發(fā)現(xiàn)對抗血清進行1∶500和1∶1 000稀釋都可以很好地檢測到Skp1蛋白,通過Western Blot進一步驗證了抗血清在植物中的應用,為通過免疫共沉淀篩選與Skp1互作蛋白奠定了試驗基礎(chǔ),同時為進一步研究黃瓜Skp1蛋白功能提供依據(jù)。
Skp1蛋白具有廣泛的生物學功能,最多的研究是對泛素蛋白酶的研究,泛素蛋白酶體系統(tǒng)(Ubiquitin-proteasome system,UPS)是目前已知的所有真核生物體內(nèi)具有高度選擇性的最為重要的蛋白質(zhì)降解途徑,參與細胞內(nèi)80%以上蛋白質(zhì)的降解,泛素蛋白酶體途徑廣泛調(diào)控植物生長發(fā)育,如激素信號轉(zhuǎn)導、胚胎發(fā)育、光信號調(diào)節(jié)、開花、干旱、自交不親和性以及植物與病原菌相互作用和免疫反應等過程。
在擬南芥的研究中Skp1蛋白參與了多個SCF復合體的形成,Skp1作為SCF復合物中的一個關(guān)鍵的骨架蛋白同時結(jié)合F-box蛋白和Cul1,調(diào)控著包括植物雄性減數(shù)分裂、生長素、赤霉素、茉莉酸和乙烯等生理和發(fā)育進程[14-17]。在許多研究中,Skp1通過與病毒互作在病毒侵染植物中發(fā)揮重要作用[18-19]。黃瓜作為多種植物病毒寄主,Skp1可能在病毒侵染黃瓜中發(fā)揮重要作用,另外研究發(fā)現(xiàn),Skp1參與糖基化調(diào)節(jié)[20]。黃瓜中Skp1功能研究鮮有報道,Skp1作為多種生物學過程的交叉點可能對黃瓜的生長發(fā)育及抗病性發(fā)揮重要作用。
[1] Cheng X, Liu Y Q, Wang G Z, et al. Proteomic identification of the oncoprotein STAT3 as a target of a novel Skp1 inhibitor[J]. Oncotarget, 2017, 8(2):2681-2693.
[2] Hao Q, Ren H X, Zhu J, et al. Overexpression ofPSK1, a SKP1-like gene homologue, fromPaeoniasuffruticosa, confers salinity tolerance inArabidopsis[J]. Plant Cell Reports, 2017, 36(1): 151-162.
[3] Li P, Miao H X, Ma Y E, et al. CrWSKP1, an SKP1-like Gene, Is Involved in the Self-Incompatibility reaction of Wuzishatangju(CitrusreticulataBlanco)[J]. International Journal of Molecular Sciences, 2015, 16(9): 21695-21710.
[4] Zhao L, Huang J, Zhao Z H, et al. The Skp1-like protein SSK1 is required for cross-pollen compatibility in S-RNase-based self-incompatibility[J]. Plant Journal, 2010, 62(1): 52-63.
[5] Takahashi N, Kuroda H, Kuromori T, et al. Expression and interaction analysis ofArabidopsisSkp1-related genes[J]. Plant & Cell Physiology, 2004, 45(1): 83-91.
[6] Sandoval D, Hill S, Ziemba A, et al. Ubiquitin-conjugating enzyme Cdc34 and ubiquitin ligase Skp1-Cullin-F-box ligase (SCF) interact through multiple conformations[J]. Journal of Biological Chemistry, 2015, 290(2): 1106-1118.
[7] Wilken F E, Ros B, Berger D K. Expression analysis of the defence gene SGT1 (suppressor of the G2 allele of skp1) in pearl millet (Pennisetumglaucum) during salicylic acid treatment[J]. South African Journal of Botany, 2009, 75(2): 427-428.
[8] Parry G, Estelle M. Auxin receptors: a new role for F-box proteins[J]. Current Opinion in Cell Biology, 2006, 18(2): 152-156.
[9] Tintignac L A, Lagirand J, Batonnet S, et al. Degradation of MyoD mediated by the SCF (MAFbx) ubiquitin ligase[J]. Journal of Biological Chemistry, 2005, 280(4): 2847-2856.
[10] 姚明月. 酵母科F-box基因家族的進化分析[D]. 楊凌: 西北農(nóng)林科技大學, 2015.
[11] Dimitrova Y N, Li J, Lee Y T, et al. Direct ubiquitination of beta-Catenin by siah-1 and regulation by the exchange factor TBL1[J]. Journal of Biological Chemistry, 2010, 285(18): 13507-13516.
[12] 宋瑜龍. 小麥雄性不育相關(guān)基因SKP1的克隆與表達分析[D]. 楊凌: 西北農(nóng)林科技大學, 2011.
[13] 馬瑞豐. 橡膠樹乳管細胞SCF-(COI1)復合物相關(guān)蛋白的基因克隆及表達分析[D]. 海口: 海南大學, 2010.
[14] Wang M Y, Song Y L, Zhang S X, et al. The analysis ofSKP1 gene expression in physiological male aterility induced by chemical hybridizing agent SQ-1 in wheat(TriticumaestivumL.) [J]. Cereal Research Communications, 2015, 43(2): 204-212.
[15] 鄭鴻平, 李逸平. 調(diào)控細胞活動不可或缺的重要分子-F-box蛋白[J]. 生命的化學, 2011, 5:619-624.
[16] Liu A, Yu Y, Duan X, et al. GsSKP21, a Glycine soja S-phase kinase-associated protein, mediates the regulation of plant alkaline tolerance and ABA sensitivity [J]. Plant Molecular Biology, 2015, 87: 111-124.
[17] Xu Y F, Sengupta T, Kukreja L, et al. MicroRNA-223 regulates cyclin E activity by modulating expression of f-box and WD-40 domain protein 7[J]. Journal of Biological Chemistry, 2010, 285(45): 34439-34446.
[18] Mudhasani R, Tran J P, Retterer C, et al. Protein kinase R degradation is essential for rift valley fever virus infection and is regulated by SKP1-CUL1-F-box (SCF)FBXW11-NSs E3 ligase[J]. PLOS Pathogens, 2016, 12(2): e1005437.
[19] 陶 濤, 王 倩, 陳相儒, 等. 水稻黑條矮縮病毒(RBSDV) P7-2與部分水稻SKP1蛋白家族成員的互作分析[C]//彭友良. 中國植物病理學會2015年學術(shù)年會論文集.北京:中國農(nóng)業(yè)科學技術(shù)出版社, 2015.
[20] 范錫麟. 小粒野生稻OmSKP1的點突變蛋白表達純化與體外互作的初步分析[D]. 長沙: 湖南農(nóng)業(yè)大學, 2013.
猜你喜歡
現(xiàn)代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設(shè)計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
當代經(jīng)濟研究(2016年5期)2016-12-01 03:12:05
現(xiàn)代農(nóng)業(yè)(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫(yī)藥現(xiàn)代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
