光裸星蟲體腔液中卵子發生的超微結構*
2018-01-22 05:28:59王慶恒張家煒郝瑞娟黃榮蓮杜曉東
海洋與湖沼 2017年1期
王慶恒 張家煒 郝瑞娟 黃榮蓮 杜曉東
(廣東海洋大學水產學院 湛江 524088)
光裸星蟲(Sipunculus nudus)又名裸體方格星蟲,為星蟲動物門習見種, 廣泛分布于熱帶、亞熱帶和溫帶的海洋潮間帶沿岸至 30m水深的海域, 是華南地區重要的海珍品之一, 具有很高的經濟價值。近年來,光裸星蟲已逐漸成為北部灣沿岸地區海水養殖新品種, 人工增養殖技術研究涉及繁殖周期(郭學武等,1993)、親體培育(鄒杰等, 2010)、苗種培育(蔣艷等,2010)、餌料開發(張琴等, 2012)、機械化采捕(陳振國等, 2015)等各個環節。目前, 光裸星蟲的基礎生物學研究相對薄弱。
配子發生是動物繁殖生物學研究的重要內容,對配子發生規律和結構特征的認識有助于提高生產上的親體促熟與催產技術水平。星蟲動物的配子發生過程比較特別, 發生早期就從生殖腺排放到體腔, 在體腔液中游離發育至成熟。星蟲動物部分物種的卵子發生已見研究報道。Gonse(1956)、Sawada(1975)分別用光鏡和電鏡觀察了普通戈芬星蟲(Golfingia vulgaris)和戈芬星蟲(lkedai)體腔中的卵細胞發育過程, 分為6個和5個不同發育時期。Rice(1974, 1988,1989); Rice等(2012)在多種星蟲動物配子形態、胚胎發育方面做了大量工作, 并報道了光裸星蟲成熟卵子的顯微和超微結構; 研究表明, 星蟲動物多為雌雄異體, 不同物種卵子的形態、大小、卵膜結構、胚胎發育過程等方面存在明顯差異。蘭國寶等(2002)報道光裸星蟲卵細胞分期成熟, 分批產卵, 在廣西海區的繁殖期為 4—9月, 繁殖盛期為 5—8月。王慶恒等(2005)利用石蠟切片觀察了光裸星蟲卵子發生的顯微結構, 將卵母細胞發育劃分為小生長期、大生長期、成熟前期和成熟期 4個階段。顧曉英等(2009)報道,可口革囊星蟲(Phascolosoma esculenta)卵子發生過程可劃分為6個階段, 卵母細胞無濾泡膜, 生發泡破裂是成熟的標志。竺俊全等(2012)研究發現可口革囊星蟲卵母細胞的卵黃發生途徑包括線粒體、內質網、高爾基體等細胞器演變成卵黃粒以及吞飲作用形成卵黃粒, 卵黃膜由卵母細胞自體形成, 由纖維層、致密層、粒狀突層及外膜構成。但是尚未有光裸星蟲卵子發生過程的超微結構觀察的詳盡報道。本研究利用透射和掃描電鏡技術觀察了光裸星蟲卵子發育過程中的超微結構的變化, 旨在積累光裸星蟲卵子發生的細胞學資料, 為探究光裸星蟲卵子發生機制,以及生殖調控與人工繁殖技術研究提供理論參考。
1 材料與方法
1.1 實驗材料
實驗用光裸星蟲采集自廣東海洋大學東海島海洋生物研究基地。活體帶回實驗室后置于30L底部鋪有 8cm細沙的塑料水槽中, 在鹽度為 30的海水中,不投餌、微充氣條件下暫養 24h, 選擇鉆沙有力、體表無損傷的健康個體用于實驗。
1.2 實驗方法
先用過濾海水清洗光裸星蟲體表沙粒。然后用5mL無菌注射器吸取 3mL2.5%戊二醛后, 抽取 2mL星蟲體腔液并轉移至離心管中, 加入 5mL2.5%戊二醛預固定; 光學顯微鏡下鏡檢體腔液中的生殖細胞進行雌雄鑒定; 將預固定20min后的雌性生殖細胞轉移至2.5%戊二醛中繼續固定40min; 0.1mol/L磷酸緩沖液洗滌3次; 1%鋨酸固定45min, 0.1mol/L磷酸緩沖液洗滌3次; 50%—100%乙醇逐級脫水后, 將沉淀細胞分為兩部分。一部分細胞以叔丁醇替代乙醇,JFD-310真空冷凍儀干燥5min, JFC-1600離子濺射儀中表面鍍膜 180s, JSM-6300 LV掃描電子顯微鏡觀察、拍照。另一部分細胞以環氧丙烷為過渡劑,Epon812包埋; Leica EM UC7超薄切片機切片; 2%醋酸雙氧鈾(15—20min)和 3.52%檸檬酸鉛(10min)雙染色; JEM-1400透射電子顯微鏡觀察、拍照。利用電鏡成像系統軟件進行細胞直徑、卵黃膜厚度等長度測量,測量5次后取平均值。
2 結果
光裸星蟲體腔液中可見到游離發育的卵原細胞和卵母細胞。根據卵母細胞細胞器及細胞核的形態、卵黃發生情況、卵膜結構變化等特征, 可將卵母細胞的發育分為卵黃形成前期、卵黃形成初期、卵黃旺盛合成期和成熟期4個階段。
2.1 卵原細胞
光裸星蟲卵原細胞呈卵圓形或近圓形, 長徑約9—11μm, 短徑為 7—9μm, 表面有皺褶(圖 1a); 核質比大, 核呈卵圓形或近圓形, 長徑約 5—6μm, 短徑約 4—5μm, 核膜平滑, 核孔很少; 核內染色質聚集為粗大顆粒沿核膜分布或在核內分散分布; 核仁小, 1個, 中位或偏位(圖 1b)。細胞質較少, 細胞器不發達, 可見少量線粒體、高爾基體和內質網。線粒體近圓形, 嵴少, 呈空泡狀; 高爾基體不發達, 膜囊數量少; 有少量囊泡狀光面內質網和粗面內質網(圖 1c)。
卵原細胞外附有1—2個濾泡細胞。濾泡細胞呈扁盤狀, 表面有大量皺褶(圖 1a); 核質比很大, 細胞核扁盤狀, 占據細胞大部分體積, 長徑約為短徑的 3倍, 核內染色質電子密度高, 沿核膜聚集分布; 細胞質薄而延展, 有少量線粒體、內質網等細胞器(圖1b、d)。濾泡細胞和卵原細胞的質膜緊密相貼(圖1d)。
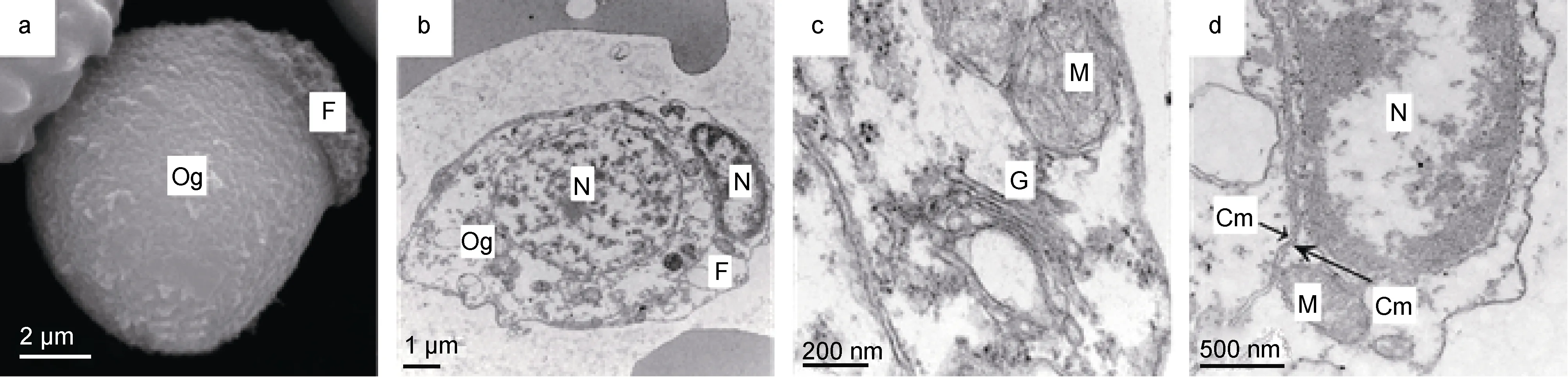
圖1 光裸星蟲卵原細胞及濾泡細胞Fig.1 The oogonium and follicular cells of S. nudus注: 子圖a: 卵原細胞, 長徑和短徑分別為9.7μm和8.6μm; 濾泡細胞, 長徑和短徑分別為5.9μm和1.3μm。b: 卵原細胞與濾泡細胞, 示細胞核。c: 卵原細胞局部, 示線粒體和高爾基體。d: 濾泡細胞局部, 示細胞核、線粒體、卵原細胞與濾泡細胞質膜緊密相貼。圖中字符Cm: 細胞膜; F: 濾泡細胞; G: 高爾基體; M: 線粒體; N: 細胞核; Og: 卵原細胞
2.2 卵黃形成前期卵母細胞
卵黃形成前期卵母細胞呈圓形或近圓形, 直徑約10—25μm(圖2a), 但核質比減小(圖2b), 細胞核圓,中位或稍偏位; 核膜平滑, 染色質為細小顆粒狀, 分散分布在細胞核中; 具1個明顯的核仁, 偏位或中位(圖 2b); 核孔增多, 高電子密度的核仁外排物通過核孔進入細胞質(圖 2c)。細胞質中尚未明顯積累卵黃,游離核糖體較多, 粗面內質網、線粒體和高爾基體數量有所增加。
卵母細胞質膜外開始形成卵黃膜, 分為兩層: 外層為顆粒層, 具有密集的中等電子密度的顆粒, 顆粒直徑約為 50nm, 顆粒層表面分布有電子密度很高的不連續的粒狀突; 內層為疏松層, 呈電子密度較低的均質結構(圖 2d)。初始時, 顆粒層先于均質層增厚,達到 600—700nm 時, 不再生長; 疏松層開始很薄,但在發育過程中可不斷增厚并發生結構變化。卵黃膜遍布微孔, 微孔為漏斗狀, 側壁在卵黃膜外表面一端形成樹枝狀分支連接粒狀突(圖2d)。
隨著卵母細胞的生長, 濾泡細胞分裂增殖, 數量逐漸增多, 包裹在卵母細胞外形成疏松的單層濾泡膜(圖2a)。濾泡細胞形態無明顯變化, 染色質沿核膜或在核內聚集分布(圖 2b、e)。濾泡細胞與卵黃膜間形成電子密度極低的膠質膜(圖2d、e), 可隨卵子發育而逐漸增厚。膠質膜有弱黏性。
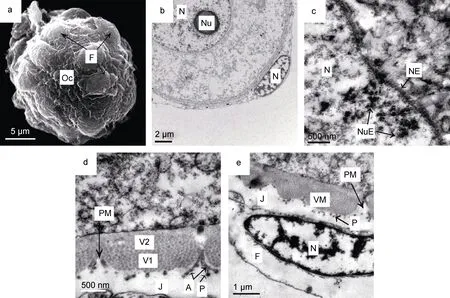
圖2 光裸星蟲卵黃形成前期卵母細胞Fig.2 The oocyte in the pre-vitellogenic stage of S. nudus注: 子圖a: 卵黃形成前期卵母細胞, 直徑為20.9μm, 示濾泡細胞。b: 卵黃形成前期卵母細胞和濾泡細胞, 示卵母細胞核、核仁。c: 卵黃形成前期卵母細胞核膜與核仁外排物。d: 卵黃膜由顆粒層和均質層構成; 顆粒層表面有電子密度很高的粒狀突, 卵黃膜孔側壁形成分支狀結構連接粒狀突。e: 濾泡細胞, 示細胞核, 濾泡細胞與卵黃膜間有膠質膜形成。圖中字符A: 分支狀連接結構; J: 膠質膜; Nu: 核仁;NE: 核膜; NuE: 核仁外排物; Oc: 卵母細胞; P: 粒狀突; PM: 卵黃膜微孔; VM: 卵黃膜; V1: 卵黃膜外層(顆粒層); V2: 卵黃膜中層(均質層)
2.3 卵黃形成初期卵母細胞
卵母細胞直徑增大至25μm左右開始進入卵黃形成初期(圖 3a), 由于膠質膜易于破裂, 掃描電鏡下, 卵黃膜上的微孔清晰可見(圖 3b), 卵質中出現少量卵黃粒(圖 3c)。卵黃膜增厚至 1.5—4μm, 其中,顆粒層厚度基本保持不變; 均勻層增厚明顯(圖3d)。微孔外側開口擴大, 使卵黃膜膜孔處的切面呈“山丘”狀, 粒狀突明顯, 膜孔側壁的分支連接結構增粗(圖3e)。濾泡細胞仍然為單層, 細胞質中可見少量線粒體、內質網、高爾基體等細胞器。由于膠質膜有所增厚, 濾泡細胞與卵黃膜間距離加大(圖 3d)。?

圖3 光裸星蟲卵黃形成初期卵母細胞及其卵膜Fig.3 The oocyte and envelope in the primary vitellogenic stage of S. nudus注: 子圖a: 卵黃形成初期卵母細胞, 直徑為35.5μm, 示濾泡細胞、膠質膜和卵黃膜。b: 卵黃膜微孔, 微孔開口平均直徑300 nm。c: 卵黃形成初期卵母細胞, 示濾泡細胞、膠質膜和卵黃膜, 胞質內有零星卵黃粒形成。d: 卵黃膜顆粒層厚650-700nm, 均質層厚1000—1100nm, 示膠質膜、濾泡細胞。e: 卵黃膜孔側壁的分支狀連接結構增粗。圖中字符Y: 卵黃粒
卵母細胞核增大, 位于細胞中部; 核仁大, 偏位;部分染色質形成高電子密度的團塊散布于核內; 核膜不平整, 形成多個突起伸入細胞質區域, 核周池明顯, 核仁外排物明顯增多(圖4a、b)。細胞質中游離核糖體豐富, 顆粒增大, 常形成多聚核糖體; 粗面內質網發達, 呈囊泡狀, 上面密布核糖體; 偶見光面內質網形成不規則的多片層結構; 高爾基體膜囊和分泌泡的數量均明顯增加; 線粒體數量多, 部分線粒體嵴短少,呈空泡狀(圖4b、c); 細胞質中常見髓樣小體(圖4e)。
根據電子密度的差異可將卵黃粒分為2種類型。其中, I型卵黃電子密度高, 有單層膜包裹, 始終保持圓形或橢圓形結構, 周圍常伴有大小不等的囊泡狀粗面內質網(圖4c), 甚至被粗面內質網包圍(圖4d)。II型卵黃粒電子密度中等, 形態為近圓形(圖 4f)。卵黃發生初期, 卵母細胞胞質中主要合成I型卵黃粒。
2.4 卵黃旺盛合成期的卵母細胞
卵母細胞直徑增大至40μm左右時開始大量積累卵黃, 進入卵黃旺盛合成期。細胞直徑迅速擴大, 在制樣過程中膠質膜因脫水等操作而發生顯著的收縮并易于破裂、脫落(圖5a、b), 卵黃膜膜孔的膜外開口明顯擴大(圖5c、d)。卵黃膜逐漸增厚至4—8μm, 超微結構也發生改變: 均質層密度發生變化, 形成很多基本平行于卵子表面的纖維(圖 5e), 并逐漸加粗(圖5f)。在纖維層與卵子質膜之間出現致密的卵黃膜內層,電子密度很高, 多在膜孔末端開始形成(圖 5e), 厚度可達2—2.5μm(圖5f)。此時, 卵黃膜由外向內分為外層(顆粒層)、中層(纖維層)和內層(致密層)。卵黃膜微孔的開口逐漸增大, 微孔通道橫切面顯示, 通道由多個低電子密度的近圓形區域圍成一圈, 構成環狀結構, 每個圓形區域中央有1個電子密度很高的核心區;在環狀結構內外兩側是卵黃膜外層顆粒(圖5g)。
卵母細胞核膜不平整, 有多個突起伸入胞質區域, 核內具1個粗大的核仁, 在核膜附近區域常見核仁外排物(圖6a)。高爾基體數量很多, 部分成對出現,扁平膜囊數量多, 附近有大量分泌泡(圖 6b); 線粒體顯著增加, 常聚集在一起, 部分線粒體呈桿狀(圖6c)。
II型卵黃粒數量迅速增加, 并不斷融合為大塊的無定形卵黃塊(圖 6d); I型卵黃粒逐漸被擠壓至卵質邊緣區域, 但不發生融合, 也不與 II型卵黃粒融合(圖 6e)。初期, 卵黃粒的分布不均勻, 近核區域卵黃粒較少(圖 6a); 隨著發育進行, 近核區域也逐漸被卵黃粒填滿。
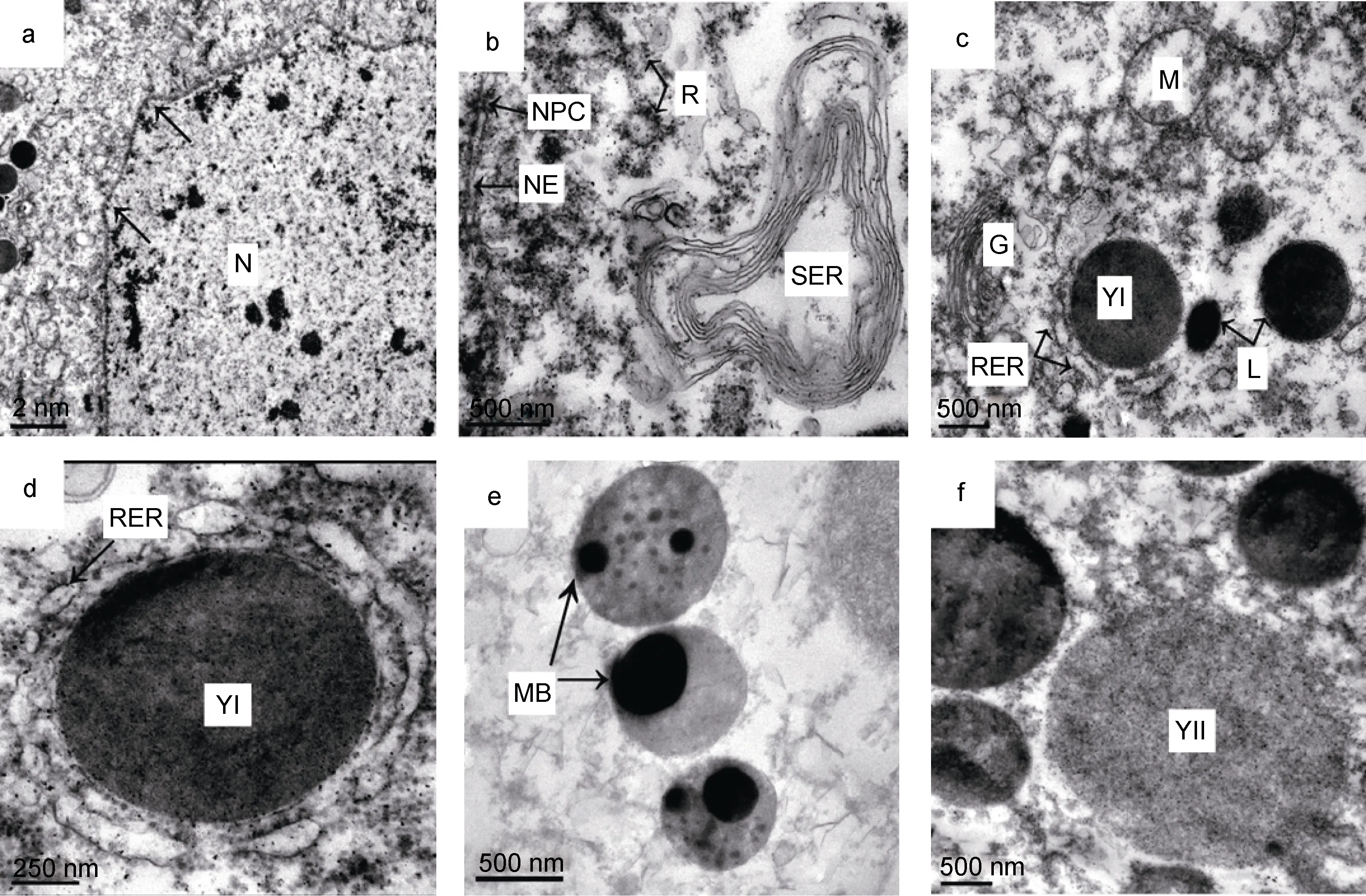
圖4 光裸星蟲卵黃形成初期卵母細胞的細胞器和卵黃Fig.4 The organelle and yolk of oocyte in the primary vitellogenic stage of S. nudus注: 子圖a: 卵黃形成初期卵母細胞局部, 示細胞核及核膜突起。b: 光面內質網形成的多片層結構、核膜和核孔復合體。c: 細胞質局部,示高爾基體、粗面內質網、溶酶體、線粒體和I型卵黃粒。d: 被粗面內質網包圍的I型卵黃粒。e: 髓樣小體。f: II型卵黃粒。圖中字符L: 溶酶體; MB: 髓樣小體; NPC: 核孔復合體; R: 核糖體; RER: 粗面內質網; SER: 光面內質網; YI: I型卵黃; YII: II型卵黃

圖5 光裸星蟲卵黃旺盛合成期卵母細胞及其卵膜Fig.5 The oocyte and envelope in the active vitellogenic stage of S. nudus注: 子圖a:卵黃旺盛合成期初期的卵母細胞, 直徑50μm, 膠質膜發生較多破裂。b: 卵黃旺盛合成期后期的卵母細胞, 直徑130μm, 膠質膜易于脫落。c: 圖a卵黃膜局部放大, 膜孔開口平均直徑375nm。d: 子圖b卵黃膜局部放大, 膜孔開口平均直徑1.7μm。e: 卵黃旺盛合成期中期卵母細胞, 卵黃膜中層增厚約6μm并出現纖維化, 開始形成高電子密度的卵黃膜內層。f: 卵黃旺盛合成期后期卵母細胞, 卵黃膜內層厚達2.8—4.4μm, 中層明顯纖維化, 示卵母細胞質膜。g: 微孔通道橫切面, 有多個低電子密度的近圓形區域圍成一圈, 構成環狀結構, 每個圓形區域中央有1個電子密度很高的核心區。圖中字符V3: 卵黃膜內層(致密層)

圖6 光裸星蟲卵黃形成初期卵母細胞的細胞器和卵黃Fig.6 The organelle and yolk of oocyte in the active vitellogenic stage of S. nudus注: 子圖a: 卵黃旺盛合成期卵母細胞局部, 示細胞核、核仁, 近核區域尚未被卵黃占據。b: 成對出現的高爾基體。c: 大量聚集出現的線粒體。d: II型卵黃粒融合為大塊的無定形卵黃塊。e: I型卵黃粒被擠壓至卵質邊緣區域, 但不發生融合。圖中字符YB: 卵黃塊
2.5 成熟期的卵母細胞
卵母細胞直徑約為 160μm左右時發育成熟。成熟卵母細胞的膠質膜可厚達 20—25μm, 但最終將和濾泡細胞一起脫落; 膠質膜已脫落的成熟的卵母細胞經后腎管收集, 排放到海水中可進行受精。電鏡制樣過程中, 膠質膜極易破裂脫落, 暴露出卵黃膜(圖7a), 卵黃膜膜孔開口可達 2—2.5μm(圖 7c)。細胞核仍然可見, 但核膜突起逐漸回縮, 核仁外排物少見(圖7b)。卵黃膜增厚至9—12μm, 中層的纖維數量增至20余條(圖7d、e), 每條纖維由大量高電子密度的短棒狀結構平行排列而構成(圖 7f); 內層厚度不均,甚至局部減小(圖7d)或消失(圖7e)。
濾泡細胞核逐漸發生縊縮, 染色質固縮并集聚在核膜下, 走向凋亡(圖7g)。
3 討論
3.1 卵子發生過程中細胞器的變化特點及其作用
無脊椎動物的卵黃可分為內源性卵黃與外源性卵黃。其中, 前者主要在卵黃發生早期, 由卵母細胞自身的細胞器合成。已有研究指出, 線粒體、內質網、溶酶體等細胞器均可以通過物質積累轉變為內源性卵黃粒。顧海龍等(2013)報道泥蚶(Tegillarca granosa)卵黃合成中線粒體和內質網最重要, 高爾基體也參與形成卵黃。朱冬發等(2006)報道內質網是東方扁蝦最重要的內源性卵黃合成胞器, 未觀察到高爾基體液泡參與卵黃粒或脂滴的形成。姜永華等(2005)認為囊泡狀粗面內質網是凡納濱對蝦(Litopenaeus vannamei)卵黃合成中最主要的細胞器, 高爾基體不直接形成卵黃粒。吳明燦等(2015)則認為高爾基體、線粒體和核糖體是糙海參(Holothuria scabra)主要的內源性卵黃合成細胞器。
本研究發現, 光裸星蟲卵母細胞發育過程中, 游離核糖體是最早大量增加的細胞器。從功能分析, 這些快速增加的核糖體的合成產物除了形成內源性卵黃以外, 也為細胞內膜系統的增加奠定基礎。進入卵黃形成初期后, 卵母細胞中溶酶體數量明顯增多; 而首先形成的I型卵黃粒具有單層膜包裹, 電子密度和形態均與溶酶體相似。因此, 推測I型卵黃粒由溶酶體演變形成, 是內源性卵黃。粗面內質網不直接轉變為卵黃粒, 而是圍繞在 I型卵黃粒周圍, 為其生長提供卵黃蛋白等營養物。在卵黃積累過程中, 線粒體數量明顯增加, 盡管有部分線粒體呈空泡狀, 但未觀察到線粒體基質中電子密度明顯增大、積累大量沉積物的現象, 因此, 光裸星蟲線粒體只是為細胞代謝提供能量, 間接參與卵黃形成, 而不直接轉變為卵黃粒。光裸星蟲卵母細胞的細胞質中沒有明顯的脂滴形成,但在卵黃旺盛合成期, 細胞中的高爾基體及其分泌囊泡數量明顯增加, 基于高爾基體功能的復雜性和多樣性, 推測高爾基體可能在多個方面參與了光裸星蟲卵母細胞發育, 包括內膜系統駐留蛋白合成、膜脂合成、膜蛋白糖基化等。
本研究發現, 光裸星蟲卵母細胞直徑達到 40μm以上時, II型卵黃快速增加, 未觀察到II型卵黃與細胞器之間的密切聯系, 推測 II型卵黃主要屬于外源性卵黃。光裸星蟲身體結構簡單, 臟器皆浸浴在體腔液中, 僅有消化道和后腎管, 無肝胰臟等器官; 另一方面, 如后文所述, 光裸星蟲濾泡細胞缺乏物質合成與分泌能力, 而是發揮選擇性吸收和轉運作用。因此,光裸星蟲外源性卵黃前體物質可能由腸道分泌到體腔液中。這與雙殼類的外源性卵黃來源于消化腺(曾志南等, 1991; 應雪萍, 2002; 顧海龍等, 2013)及甲殼類外源性卵黃可能來源于肝胰腺(堵南山等, 1999)或是濾泡細胞(洪水根, 1992)不同。操向文等(2009)利用石蠟切片觀察發現, 光裸星蟲腸壁薄, 漿膜層有大量圓形的顆粒細胞, 這些顆粒細胞在釋放了胞質顆粒后呈空泡狀。某些類型的顆粒細胞的分泌活動是否與外源性卵黃的合成有關值得深入研究。
與光裸星蟲不同, 可口革囊星蟲卵黃卵黃旺盛合成期卵母細胞中有大量脂滴(竺俊全等, 2012)。類似的近緣物種卵子物質積累的差異在甲殼動物中已有報道。楊筱珍等(2009)比較發現, 三疣梭子蟹(Portunus trituberculatus)和中華絨螯蟹(Eriocheir sinensis)卵母細胞中存在大量脂滴, 但日本新糠蝦(Neomysis japonica)卵子中卻沒有脂滴分布。研究表明, 甲殼動物卵母細胞的脂滴主要由中性脂構成, 胚胎發育過程較長的物種的卵母細胞含有較多中性脂(成永旭等, 1999)。在28—29°C條件下, 可口革囊星蟲胚胎發育時間為 45h30min(金春華等, 2011), 光裸星蟲胚胎發育時間為38—40h(蘭國寶等, 2003), 胚胎發育時間的差異可能與卵母細胞中脂滴(中性脂)積累存在相關性, 但需要更多研究證據。
3.2 卵子發生過程中卵膜結構變化特點及其作用
已知的星蟲動物卵子均具有厚而分層的卵黃膜,并有很多卵膜孔。本研究發現, 光裸星蟲卵母細胞的卵黃膜分為三層。其中, 中層為卵黃膜的主體, 初始時為低電子密度的均質結構, 很薄, 在卵母細胞的發育過程中不斷增厚, 可達 9—12μm, 并不斷發生纖維化, 形成 20余條基本平行于卵子質膜表面的粗大纖維。作者認為, 這種較厚而致密的卵黃膜有助于卵母細胞緩沖機械壓力, 是星蟲動物對卵子游離發育方式的一種保護性適應; 卵膜孔作為外源性物質進入卵母細胞的通道, 可解決較厚的卵黃膜與大量營養物質跨卵黃膜運輸的矛盾。竺俊全等(2012)認為可口革囊星蟲成熟卵母細胞的卵膜孔未關閉, 體腔液中的營養物質仍可通過微孔進入卵內。此外, 作者在胚胎發育觀察中發現, 光裸星蟲卵膜孔與原腸胚體表纖毛形成存在相關性(另文發表)。
與大多數星蟲卵母細胞卵黃膜裸露不同, 光裸星蟲卵黃膜外具有一層透明的膠質膜。蘭國寶等(2002)顯微觀察認為, 光裸星蟲卵細胞直徑在 70μm以上時, 膠質膜開始形成。本研究超微結構觀察發現,膠質膜在卵母細胞卵黃形成前(小于 25μm)已經開始形成, 研究手段分辨率的顯著差異是造成本研究與上述報道結果不一致的主要原因。正常卵母細胞的膠質膜可厚達 20—25μm, 電子密度極低, 呈完全的“透明”狀, 沒有可見的精細結構, 顯然這同樣在增強對卵子物理保護的同時利于物質的運輸。另一方面, 膠質膜具有黏性, 推測含有較多的粘多糖, 是否存在對卵母細胞的化學保護功能還有待深入研究。
水生生物卵母細胞外常見濾泡細胞, 這些濾泡細胞(或其分化細胞)大多具有分泌功能。例如, 哲羅魚(Hucho taimen)部分濾泡細胞在卵母細胞發育到IV時相分化為顆粒細胞, 胞內密集分布著大量的內質網、線粒體和分泌顆粒(張永泉等, 2013)。東方扁蝦的濾泡細胞富含游離核糖體、粗面內質網和線粒體,合成活性較強(朱冬發等, 2006)。本研究顯示, 光裸星蟲卵母細胞外包圍著疏松排列的單層濾泡細胞, 在卵子發育過程中, 濾泡細胞數量有所增加, 但形態結構變化小, 細胞質很薄, 核糖體、線粒體、內質網等細胞器少。顯然, 這些濾泡細胞缺乏物質合成與分泌的能力, 難以滿足卵母細胞發育對卵黃積累的需求。可口革囊星蟲和革囊星蟲(agassizii)的卵母細胞外均沒有濾泡細胞, 普通戈芬星蟲的濾泡細胞在卵母細胞發育早期階段即消失(Rice, 1989; 竺俊全等, 2012),說明這些星蟲動物的濾泡細胞不參與合成卵黃前體物質。成永旭等(2002)認為鋸緣青蟹(Scylla serrata)卵泡細胞沒有合成與分泌的作用, 而是類似于“過濾器”, 允許血淋巴中的卵黃前體物質或卵黃脂蛋白通過卵泡細胞而進入卵母細胞。作者認為, 光裸星蟲的濾泡細胞不是外源性卵黃的合成部位, 而是與鋸緣青蟹卵泡細胞類似, 起著選擇性吸收與轉運的作用。
本研究發現, 采集自湛江遂溪的光裸星蟲成熟卵的膠質膜厚約 20—25μm, 卵黃膜厚 9—12μm, 卵黃膜纖維多達20余條; Rice(1989)報道光裸星蟲波多黎各群體成熟卵的膠質膜為 12—13μm, 卵黃膜平均厚度為 5μm, 卵黃膜纖維僅 3—4條。兩個群體成熟卵的卵膜結構存在較大差異。光裸星蟲不同地理群體的成熟卵卵徑也存在明顯差異: 光裸星蟲坦帕灣群體(Rice, 1988)、波多黎各群體(Rice, 1989)、廈門群體(郭學武, 1993)成熟卵的平均直徑分別為120、178和170μm。地理隔離造成的群體遺傳結構差異可能是導致卵徑大小和卵膜結構差異的主要原因, 現有研究已經證實光裸星蟲群體間存在明顯的遺傳分化: 王慶恒等(2008)報道了光裸星蟲湛江群體與沖繩群體的染色體數目相同, 但染色體組型明顯不同; RAPD分析(王慶恒等, 2006)、16S rRNA(Du et al, 2008)和COI(Du et al, 2009)基因序列比較均顯示, 光裸星蟲多態位點比例和遺傳多樣性指數較高, 群體間分化明顯。
Rice(1989)將星蟲胚胎發育分為 3種類型(D型:無幼蟲直接發育; L型: 具有完全依賴卵黃營養的幼蟲期; P型: 具有以浮游生物為營養的幼蟲期), 并統計了23種星蟲動物的成體規格、成熟卵形狀、卵徑、卵色與胚胎發育類型之間的相關性, 結果顯示, 只有卵徑與發育類型間存在顯著相關(P<0.05), 即D型星蟲的卵徑最大(135—280μm), L型星蟲的卵徑次之(125—175μm), P 型星蟲卵徑最小(<125μm)。可口革囊星蟲和光裸星蟲均需要經過擔輪幼蟲、海球幼體及變態等發育階段才能發育為稚蟲, 為典型的 P型星蟲。其中, 可口革囊星蟲成熟卵卵徑約為115—140μm(顧曉英等, 2009), 與 Rice的分析結果基本一致; 但光裸星蟲波多黎各(Rice, 1989)、廈門(郭學武等,1993)、北海(蘭國寶等, 2002)、湛江群體(王慶恒等,2005)的成熟卵卵徑均達到 160μm 以上, 不符合上述規律。
4 結論
光裸星蟲卵子在體腔液中游離發育, 卵外有濾泡細胞。發育過程中, 細胞器數量、細胞核形態、卵黃發生情況、卵膜的形態結構等發生復雜變化, 可將卵母細胞的發育劃分為卵黃形成前期、卵黃形成初期、卵黃旺盛合成期和成熟期4個階段。
王慶恒, 杜宇宏, 林斯里等, 2008. 裸體方格星蟲染色體組型分析. 動物學雜志, 43(5): 95—98
王慶恒, 杜曉東, 李 康, 2006. 光裸星蟲遺傳多樣性的RAPD分析. 海洋水產研究, 27(3): 57—61
王慶恒, 杜曉東, 黃紅艷等, 2005. 湛江地區光裸星蟲的生殖細胞發育和生殖周期. 湛江海洋大學學報(自然科學版),25(1): 5—9
蘭國寶, 閻 冰, 2002. 方格星蟲繁殖生物學研究. 水產學報,26(6): 503—509
蘭國寶, 閻 冰, 廖思明, 2003. 方格星蟲胚胎與幼體發育的研究. 熱帶海洋學報, 22(6): 70—75
成永旭, 李少菁, 王桂忠等, 2002. 鋸緣青蟹卵黃發生期卵母細胞和卵泡細胞之間的結構變化. 動物學報, 48(1):80—92
成永旭, 堵南山, 賴 偉, 1999. 中華絨螯蟹成熟卵巢的脂類及脂肪酸組成. 中國水產科學, 6(1): 79—82
朱冬發, 王桂忠, 李少菁, 2006. 東方扁蝦卵子發生的超微結構. 水生生物學報, 30(4): 439—445
楊筱珍, 王金峰, 楊麗娜等, 2009. 日本新糠蝦(Neomysis japonica)卵巢發育與卵子發生. 海洋與湖沼, 40(3): 338—346
吳明燦, 張 立, 潘 英等, 2015. 糙海參卵子發生及卵黃發生的超微結構. 熱帶海洋學報, 34(3): 68—74
鄒 杰, 彭慧婧, 蔣 艷等, 2010. 方格星蟲親體培育試驗.漁業現代化, 37(3): 30—33
應雪萍, 2002. 文蛤卵母細胞卵黃發生的超微結構. 中國水產科學, 9(2): 125—128
張 琴, 童 潼, 童萬平等, 2012. 飼料蛋白水平對方格星蟲稚蟲日增重和消化酶活性的影響. 漁業現代化, 39(2):41—47
張永泉, 尹家勝, 杜 佳等, 2013. 哲羅魚(Hucho taimen)卵巢濾泡細胞發生的超微結構研究. 海洋與湖沼, 44(1):262—266
陳振國, 班廷輝, 2015. 方格星蟲捕捉機: 中國,CN201420615340.0
竺俊全, 王 偉, 丁理法, 2012. 可口革囊星蟲(Phascolosoma esculenta)卵黃合成期卵母細胞發育及卵黃發生與卵膜形成的超微結構. 海洋與湖沼, 43(4): 870—876
金春華, 竺俊全, 許式見等, 2011. 可口革囊星蟲(Phascolosoma esculenta)胚胎及幼蟲發育研究. 海洋與湖沼, 42(1): 94—100
姜永華, 顏素芬, 宋振榮, 2005. 凡納濱對蝦卵母細胞卵黃發生的超微結構. 動物學報, 51(1): 133—141
洪水根, 1992. 長毛對蝦卵子發生的研究 II. 卵黃發生. 海洋與湖沼, 23(3): 280—284
顧曉英, 竺俊全, 許式見等, 2009. 可口革囊星蟲(Phascolosoma esculenta)卵子發生的組織學研究. 海洋與湖沼, 40(3):283—288
顧海龍, 林志華, 沈偉良等, 2013. 泥蚶初級卵母細胞發育及卵黃發生的超微結構研究. 海洋科學, 37(1): 49—53
郭學武, 李復雪, 1993. 光裸星蟲生殖周期的研究. 熱帶海洋,12(2): 69—76
堵南山, 賴 偉, 陳鵬程等, 1999. 中華絨螯蟹卵黃形成的研究. 動物學報, 45(1): 88—92
蔣 艷, 蔡德建, 鄒 杰等, 2010. 方格星蟲苗種池塘中間培育試驗研究. 廣西科學, 17(2): 175—177
曾志南, 李復雪, 1991. 青蛤卵母細胞發育過程和卵黃發生的超微結構研究. 海洋學報, 13(5): 679—683
操向文, 王慶恒, 黃若男等, 2009. 光裸星蟲消化系統的解剖學和組織學研究. 廣東海洋大學學報, 29(3): 6—10
Du X D, Chen Z, Deng Y W et al, 2008. Genetic Diversity and Population Structure of the Peanut Worm (Sipunculus nudus)in Southern China as Inferred from Mitochondrial 16S rRNA Sequences. The Israeli Journal of Aquaculture-Bamidgeh,60(4): 237—242
Du X D, Chen Z, Deng Y W et al, 2009. Comparative analysis of genetic diversity and population structure of Sipunculus nudus as revealed by mitochondrial COI sequences.Biochemical Genetics, 47(11—12): 884—891
Gonse P H, 1956. L’ovogenese chez Phascolosoma vulgare: Ⅱ.Recherches biométriques sur les ovocytes. Acta Zoologica,37(3): 225—233
Rice M E, 1974. Gametogenesis in three species of Sipuncula:Phascolosoma agassizi, Golfingia pugettensis, and Themiste pyroides. La Cellule, 79(2—3): 259—313
Rice M E, 1988. Observations on development and metamorphosis of Siphonosoma cumanense with comparative remarks on Sipunculus nudus (Sipuncula, Sipunculidae). Bulletin of Marine Science, 42(1): 1—15
Rice M E, 1989. Comparative observations of gametes,fertilization, and maturation in sipunculans. In: Ryland J S ed. Reproduction, Genetics and Distributions of Marine Organisms: 23rd European Marine Biology Symposium.Swansea: School of Biological Sciences, University of Wales, 167—183
Rice M E, Reichardt H F, Piraino J et al, 2012. Reproduction,development, growth, and the length of larval life of Phascolosoma turnerae, a wood-dwelling deep-sea sipunculan. Invertebrate Biology, 131(3): 204—215
Sawada N, 1975. An electron microscopic study on the oogenesis of Sipunculan worms. In: Mary E, Todorovic M, eds.Proceedings of the International Symposium on the Biology of the Sipuncula and Echiura. Belgrade: Institute for Biological Research, 169—176
