嗜水氣單胞菌cpxRA缺失株構建及生物學特性研究
2018-01-24 02:24:07張?zhí)m梨張倩倩王謝昊吳振兵馮宇晴李愛華
水生生物學報 2018年1期
關鍵詞:系統(tǒng)
張?zhí)m梨 張倩倩 陳 輝 王謝昊 吳振兵 馮宇晴 李愛華
(1. 中國科學院水生生物研究所, 淡水生態(tài)與生物技術國家重點實驗室, 武漢 430072; 2. 中國科學院大學, 北京 100049;3. 淡水水產(chǎn)健康養(yǎng)殖湖北省協(xié)同創(chuàng)新中心, 武漢 430070; 4. 江蘇省漁業(yè)技術推廣中心, 南京 210036)
嗜水氣單胞菌(Aeromonas hydrophila)屬于弧菌科氣單胞菌屬, 是一種革蘭氏陰性短桿菌, 廣泛分布于自然界各種水體及淤泥、土壤中, 對水產(chǎn)動物、畜禽和人均具有致病性[1—4], 尤其是對水產(chǎn)動物危害嚴重, 可導致水產(chǎn)動物患敗血癥、腸炎、爛尾病、腹水病等[5], 造成水產(chǎn)養(yǎng)殖業(yè)嚴重的經(jīng)濟損失。
細菌生存和繁殖的前提是適應環(huán)境[6], 這就需要細菌多種分子調控系統(tǒng)來有效整合外界信息并控制細菌行為應對方式。細菌分子調控系統(tǒng)包括單組分、雙組分及三組分調控系統(tǒng), 其中雙組分調控系統(tǒng)處于中心地位。CpxRA系統(tǒng)是革蘭氏陰性菌中普遍存在的一種雙組分系統(tǒng), 由細胞質內膜上的組氨酸蛋白激酶CpxA和細胞質內的反應調節(jié)蛋白CpxR組成。當CpxA蛋白感知到外界信號分子變化時, 通過自磷酸化使一個保守的組氨酸殘基帶上磷酸基團, 然后將磷酸基團轉移到細胞質內反應調節(jié)蛋白CpxR的天冬氨酸殘基上, 磷酸化的CpxR被激活, 會與靶基因啟動子區(qū)的特異序列結合, 激活轉錄[7]。已有學者對大腸桿菌、霍亂弧菌、遲緩愛德華氏菌等的CpxRA雙組分系統(tǒng)進行深入研究, 但有關嗜水氣單胞菌中Cpx系統(tǒng)的作用則鮮有報道。因此本文通過構建嗜水氣單胞菌株DBHS101的cpxRA雙基因缺失突變株Δcpx來研究Cpx系統(tǒng)在嗜水氣單胞菌的功能, 為后續(xù)深入認識該系統(tǒng)提供參考。
1 材料與方法
1.1 菌株及質粒
嗜水氣單胞菌DBHS101由本實驗室于2009年6月在患病鳙魚腎臟分離得到[8], 具有氨芐青霉素抗性; 大腸桿菌S17-1λpir和pACYC184質粒由中國科學院水生生物研究所謝海俠博士饋贈; pMD18-T質粒購自大連TaKaRa公司; pRE112質粒由華中農(nóng)業(yè)大學林蠡博士饋贈。
1.2 主要試劑和儀器
細菌基因組提取試劑盒、DNA Marker、2×TaqMasterMix購自北京康為世紀生物科技有限公司;質粒提取試劑盒購于OMEGA公司; 限制性內切酶、T4 DNA連接酶、PrimeSTAR Max DNA Polymerase、SYBR GreenⅠ購自大連TaKaRa公司; 蔗糖、氯霉素、氨芐青霉素購自Sigma公司。PCR儀購于Thermo公司; 核酸電泳儀購于Bio-Rad公司; 引物合成于北京擎科新業(yè)生物技術有限公司(表 1)。
1.3 基因缺失株及互補株的構建和鑒定
基因缺失株的構建和鑒定根據(jù)Ho等[9]的方法利用融合PCR獲得基因缺失片段, 通過同源重組篩選出缺失突變株。以嗜水氣單胞菌DBHS101的基因組為模板, 根據(jù)已公布的Aeromonas hydrophilaML09-119 (CP005966)、JBN2301 (CP013178)、NJ-35 (CP006870)全基因組序列, 同源克隆擴增得到DBHS101的cpxRA基因簇全序列, 用Primer Premier 5.0對序列設計引物。用引物cpx-5O/5I和cpx-3I/3O分別擴增cpx基因的上下游片段F1、F2,然后用F1、F2片段為模板, 用cpx-5O/3O進行融合PCR得到大片段F1F2, 將F1F2片段連接到pMD18-T載體上送測序, 驗證片段序列正確后, 在限制性內切酶XbaⅠ和KpnⅠ及T4 DNA連接酶的作用下將片段與自殺性質粒pRE112連接起來構建重組質粒pRE-Δcpx; 把重組質粒轉入大腸桿菌S17-1λpir感受態(tài)中, 然后利用接合轉移法將重組質粒pRE-Δcpx轉入受體菌DBHS101中進行同源重組, 通過含氨芐青霉素(Amp 100 μg/mL)、氯霉素(Cm 25 μg/mL)或蔗糖(SacB 0.1 g/mL) 的TSA平板篩選出SacBRCms表型菌株, 即為陽性菌株, 用cpx-5O/3O及兩對檢測引物cpx-outcheck F/R、cpx-incheck F/R對陽性菌株進行PCR驗證, 條帶大小符合期望值的即為cpxR-A基因缺失突變株Δcpx。雙交換及篩選過程如圖1。
互補株CΔcpx的構建以DBHS101的基因組為模板, 用互補引物com-F/com-R擴增cpx互補片段, 連T載測序正確后經(jīng)限制性內切酶Hind Ⅲ 和SphⅠ 雙酶切后連接pACYC184載體構成互補質粒pACYC-cpx, 互補質粒電轉入缺失突變株Δcpx中,即可構建互補株, 經(jīng)氯霉素抗性平板篩選和PCR驗證挑出陽性克隆, 送測序。
1.4 掃描電鏡觀察
劃線后分別挑取野生株、突變株和互補株的單菌落在TSB中過夜培養(yǎng), 按照1%比例轉接至底部放有蓋玻片的新鮮的TSB培養(yǎng)基中, 28℃靜置培養(yǎng)24h, 取出蓋玻片用無菌PBS沖洗3次; 接著用2.5%戊二醛4℃固定4h, 之后依次用10%、30%、50%、70%、90%和無水乙醇脫水, 叔丁醇進行置換, 最后真空冷凍干燥, 噴金進行掃描電鏡觀察。
1.5 生長曲線、運動能力及生物膜形成能力測定
挑取野生株、突變株的單菌落在5 mL TSB中過夜培養(yǎng), 調節(jié)吸光度值A600=1.0, 按照1%比例分別轉接至新鮮的TSB培養(yǎng)基中28℃ 180 r/min進行振蕩培養(yǎng), 每1h取樣測定OD600, 連續(xù)觀察12h并繪制生長曲線; 當菌株長至A600=1.0時, 取2 μL接種至含0.35%瓊脂的半固體TSB平板, 28℃ 靜置培養(yǎng)24h后觀察并測量泳動圈大小; 采用96孔法[10]進行生物膜的測定, 在菌株按照1%比例轉接至新鮮TSB后, 立即將含菌的培養(yǎng)基以200 μL/孔的量轉移到三個96孔細胞培養(yǎng)板中, 分別在24h、48h、96h各取出一板, 棄掉培養(yǎng)基, 用PBS洗滌3次自然風干后, 以Bouin’s固定液固定1h之后加入1%結晶紫染液, 染色30min。最后用蒸餾水洗滌3次, 待干燥后加入95%乙醇溶解, 靜置10min; 以無菌培養(yǎng)基做空白對照, 用酶標儀檢測570 nm波長(結晶紫最大吸收波長)各孔的光吸收值從而比較生物膜含量差異。

表 1 本研究中使用的引物Tab. 1 Primers used in this study

圖1 cpxR-A基因缺失株的構建過程Fig. 1 Construction of cpxR-A mutant strain
1.6 刺激耐受實驗
急性刺激挑野生株和突變株的單菌落活化后按照1%比例擴大培養(yǎng)至A600=0.5, 4℃ 8000×g離心5min, 加入TSB重懸; 之后取100 μL菌液加入到900 μL TSB (對照組)或含刺激因子的TSB (實驗組)中, 28℃, 180 r/min培養(yǎng)30min; 對照組和實驗組分別用TSB進行連續(xù)十倍倍比稀釋, 每個濃度稀釋液各取10 μL在含Amp的TSA平板上點樣[11], 28℃培養(yǎng)12—14h后計數(shù), 比較野生株和突變株的存活率差異(存活率%=實驗組CFU/對照組CFU×100%)。
刺激因子及終濃度0.02 mol/L HCl、0.02 mol/L NaOH、0.5 mol/L KCl、0.5 mg/mL SDS、5 mg/mL EDTA,2 μm孔徑濾膜過濾除菌。
慢性刺激挑單菌落活化后, 培養(yǎng)至A600=1.0時, 按照1%比例轉接至30 mL含刺激因子的TSB培養(yǎng)基(對照組不含刺激因子)中28℃ 180 r/min培養(yǎng)12h, 每小時測定A600值。刺激因子及終濃度5 mg/mL EDTA溶液, 20 μg/mL多黏菌素B溶液。
1.7 致病性相關實驗
斑馬魚浸泡感染實驗挑取野生株和突變株的單菌落活化后擴大培養(yǎng)至對數(shù)期, 用PBS洗滌3次后調節(jié)菌液濃度為108CFU/mL, 分別用1 L的野生株、突變株菌懸液或PBS各浸泡15條健康斑馬魚24h, 之后每天正常換水, 但不喂食觀察1周內魚的生長狀態(tài)及存活情況。
斑馬魚腹腔注射攻毒實驗挑取野生株和突變株的單菌落活化后擴大培養(yǎng)至對數(shù)期(培養(yǎng)約2.5h), 用PBS調節(jié)菌液濃度(野生株濃度分別是1.29×105、6.45×104、3.23×104和1.62×104CFU/mL;突變株濃度為1.32×105、6.59×104、3.30×104和1.65×104CFU/mL)[12], 然后各取50 μL分別腹腔注射斑馬魚(平均長度為3 cm, 隨機分為9組, 每組15條)對照組注射等量PBS, 統(tǒng)計2周內斑馬魚存活情況, 根據(jù)軟件SPSS 16.0 通過幾率單位加權回歸法(Bliss)計算斑馬魚半數(shù)致死量LD50。
1.8 實時熒光定量PCR
為了進一步驗證Cpx系統(tǒng)相關基因表達量的變化, 篩選了與Cpx系統(tǒng)可能相關的基因序列設計引物[11,13,14](表 2)進行熒光定量分析。培養(yǎng)野生株和突變株至對數(shù)期, 然后提細菌RNA, 測定其濃度及電泳檢測質量后進行反轉錄, 以野生株作為對照組,選取穩(wěn)定表達的16S rRNA基因作內參, 進行熒光定量PCR。反轉錄和熒光定量PCR過程嚴格按照試劑盒說明書進行操作。
根據(jù)前期實驗, 選取5 mg/mL EDTA作為誘導劑, 當菌株培養(yǎng)至對數(shù)前期(A600為0.2—0.3)時, 加入EDTA溶液或等量無菌雙蒸水(對照組), 繼續(xù)培養(yǎng)30min, 提取細菌RNA, 迅速反轉錄得到cDNA后,選取外膜相關的5個基因進行熒光定量PCR, 比較野生株、突變株中這些基因經(jīng)EDTA處理后的表達量變化。
實時熒光定量PCR反應體系為20 μL: SYBR GreenⅠMix 10 μL, 上、下游引物(10 μmol/L)各0.8 μL, 模板cDNA 1 μL, DEPC H2O 7.4 μL。
PCR反應條件為: 95℃ 預變性2min; 95℃ 變性10s, 58℃ 退火30s, 72℃ 延伸30s, 40個循環(huán)。
野生株和突變株各3個生物學重復, 每1個樣品重復3次, 基因轉錄水平的變化根據(jù)2-ΔΔCt方法[15]計算。

表 2 熒光定量PCR引物Tab. 2 Primers used in qRT-PCR
1.9 數(shù)據(jù)分析
實驗結果圖通過GraphPad Prism 5.01繪制, 采用雙尾t檢驗方法, 但斑馬魚浸泡實驗的結果采用的是log-rank (Mantel-Cox) 檢驗, 顯著性水平定義為P<0.05, 在圖中用“*”標出,P<0.01, 用“**”標出。
2 結果
2.1 基因缺失株、互補株的鑒定
以嗜水氣單胞菌野生株DBHS101為模板, 通過PCR分別獲得cpxRA基因的上下游同源臂F1、F2(理論值F1=646 bp F2=505 bp), 然后進行融合PCR得到缺失cpxRA基因部分片段的上下游片段F1F2 (理論值F1F2=1116 bp), 結果如圖2A所示。對構建的重組質粒進行XbaⅠ/KpnⅠ雙酶切驗證(圖2B)并送測序, 測序結果正確。通過接合轉移和同源重組, 在氨芐、蔗糖和氯霉素的壓力條件下逐步篩選得到了嗜水氣單胞菌cpxRA基因缺失株Δcpx,以cpx-incheck F/R、cpx-5O/3O、cpx-outcheck F/R三對引物同時對野生株和突變株進行PCR驗證,結果如圖2C所示, 和理論值相符(理論值: 野生株做模板時3對引物擴增產(chǎn)物大小應該為2649、2921和3325 bp, 陽性突變株做模板3對引物擴增產(chǎn)物大小應該為826、1116和1501 bp), 說明構建的缺失株Δcpx正確。
以嗜水氣單胞菌野生株DBHS101為模板, 以com-F/com-R為引物擴增得到cpxRA互補片段, 將片段與質粒pACYC184酶切連接后電轉入突變株Δcpx中, 篩選在含Cm的TSA平板上生長的單菌落,以com-F/com-R為引物, 野生株和突變株為對照進行PCR來驗證, 結果如圖3所示, 符合預期值(理論值: 野生株為模板時PCR產(chǎn)物應該為2238 bp, 突變株為模板時PCR產(chǎn)物應該為415 bp), 將互補株PCR產(chǎn)物送測序, 序列正確, 說明互補株CΔcpx構建成功。
2.2 掃描電鏡觀察
載有菌液的玻片在固定、冷凍、干燥后用掃描電鏡進行觀察, 結果如圖4(進行了3次重復, 未顯示所有重復圖), 結果顯示在20000倍放大下, 野生株、突變株和互補株的形態(tài)沒有明顯差異。
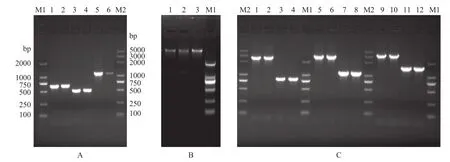
圖2 基因缺失株的構建和鑒定Fig. 2 Construction and identification of cpxRA mutant strain
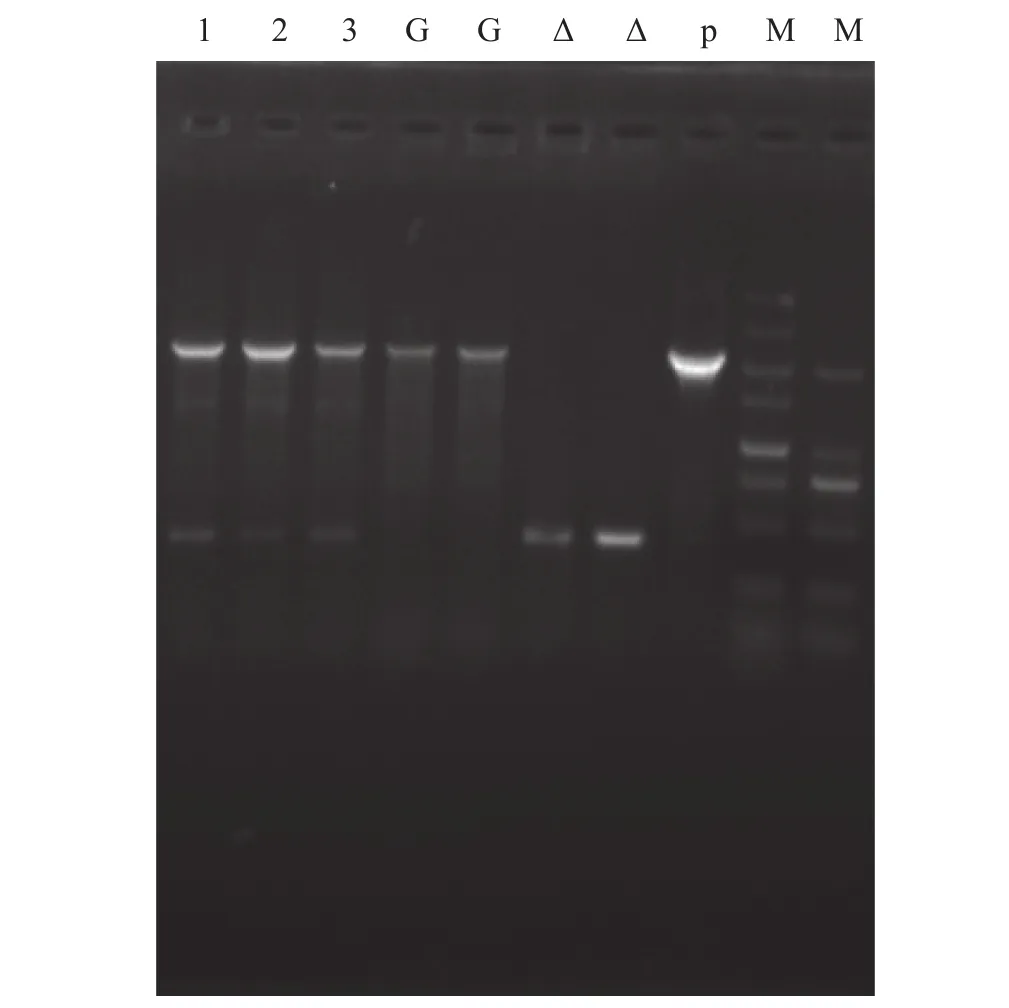
圖3 互補株的鑒定Fig. 3 Identification of cpxRA complementary strain
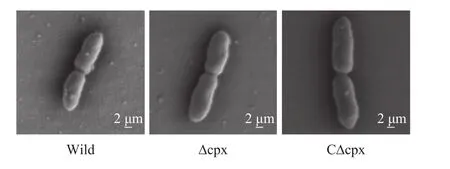
圖4 菌株掃描電鏡圖Fig. 4 SEM images of strains
2.3 生長曲線、運動能力及生物膜形成能力測定結果
野生株和缺失株生長曲線如圖5A, 兩者的生長曲線無明顯差異, 說明cpxRA基因同時缺失不影響嗜水氣單胞菌DBHS101正常條件下的生長; 運動能力結果如圖5B, 對數(shù)據(jù)進行統(tǒng)計分析, 野生株、缺失株和互補株間泳動能力無明顯差異。
在生物膜形成能力測定中, 通過測定生物膜吸附的結晶紫含量來分析菌株在靜置培養(yǎng)24h、48h和96h后形成的生物膜濃度, 結果如圖6所示, 突變株生物膜形成能力較野生株稍低, 且在96h兩者差距更大, 但沒有達到顯著性水平; 而互補株則在24h時與野生株相似, 但在48h、96h菌膜形成較野生株少,這可能是因為后期菌株營養(yǎng)缺乏時互補株表達抗性基因會耗用能量。從總的結果分析,cpxRA基因簇對生物膜形成有一定作用但不是其關鍵因子。
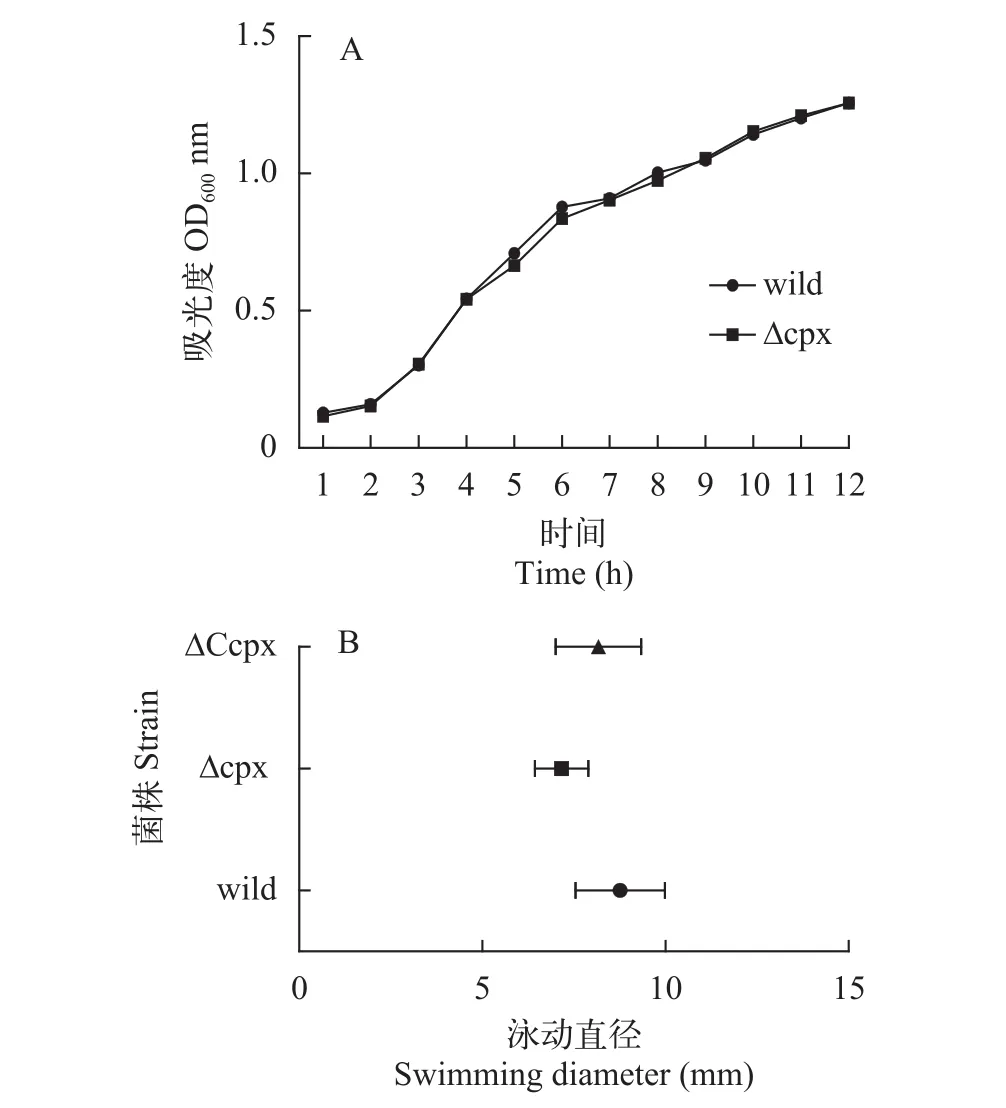
圖5 菌株的生長曲線及泳動直徑Fig. 5 The growth curve of strains and motility assays

圖6 生物膜形成能力Fig. 6 The assay of biofilm formation
2.4 刺激耐受實驗
急性刺激實驗分別選取酸性、堿性、高滲透壓及去污劑和螯合劑刺激作為實驗對象, 比較這些刺激對野生株和缺失株存活率的影響, 從而了解Cpx系統(tǒng)和這些刺激應對機制間的關系。結果如圖7, 去污劑SDS存在環(huán)境下突變株較野生株存活率顯著增加, 而高滲透壓環(huán)境下突變株相比于野生株存活率則是顯著下降的, 在其他環(huán)境中兩者差異不顯著, 這說明在嗜水氣單胞菌中Cpx雙組分系統(tǒng)參與細菌對高滲透壓和SDS刺激的應答。
慢性刺激實驗為了了解菌株對刺激的一個動態(tài)反應過程, 采用了測定吸光度值繪制生長曲線的方法來觀察突變株和野生株對影響外膜結構的刺激的表現(xiàn)。結果如圖8所示, 可以看出野生株和突變株在這2種刺激條件下生長情況有著顯著差異, 在觀察的12h內, 突變株在含有EDTA的環(huán)境中依然能緩慢生長, 而野生株則一直處于生長停滯狀態(tài); 在多黏菌素B刺激下, 突變株反而停滯生長, 而野生株則是持續(xù)生長。
2.5 毒力試驗結果
斑馬魚浸泡攻毒試驗以108CFU/mL濃度的野生株和缺失株菌液1 L同時各浸泡15條健康斑馬魚24h, 之后每天換水, 觀察1周內斑馬魚的存活情況, 以PBS做空白對照。1周后根據(jù)實驗結果繪制斑馬魚累積存活曲線(圖9)并用log-rank (Mantel-Cox)檢驗進行差異分析, 結果顯示通過菌液浸泡感染, 野生株對斑馬魚毒力較缺失株較大, 但差異不顯著(P=0.283>0.05)。
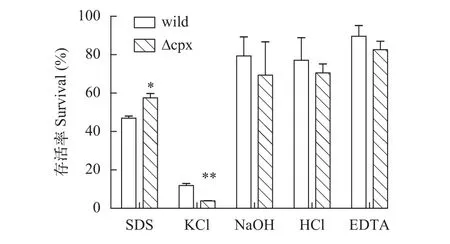
圖7 不同急性刺激下菌株存活率Fig. 7 The survival of strains in different acute stimulus
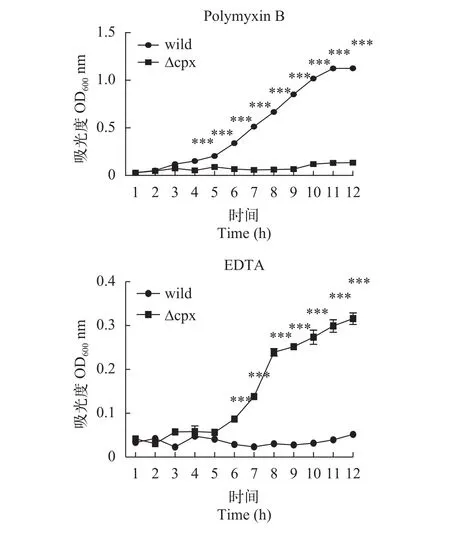
圖8 在不同刺激下菌株的生長曲線Fig. 8 The growth curve of strains in different stimulus
斑馬魚腹腔注射攻毒試驗分別用4個濃度菌液各50 μL腹腔注射斑馬魚(n=15), 同時以PBS作對照。斑馬魚死亡情況如表 3, 經(jīng)計算LD50(wild)=6.19×102cfu/尾,LD50(Δcpx)= 7.00×102cfu/尾, 說明野生株和缺失株腹腔注射對斑馬魚的毒力均很強,并且缺失株毒力較野生株稍弱。
2.6 實時熒光定量PCR
提取細菌對數(shù)期的RNA進行反轉錄, 然后選取嗜水氣單胞菌毒力蛋白基因、其他雙組分系統(tǒng)基因及菌毛基因等共15個基因設計引物進行熒光定量PCR檢測, 結果如圖10。通過熒光定量PCR, 再次驗證了突變株的cpxR和cpxA基因缺失, 并且可以看到突變株的毒力基因hly、aer、alt相比野生株顯著上調, 但幅度不大; 另外可以發(fā)現(xiàn)CpxRA雙組分系統(tǒng)的缺失會導致OmpR-EnvZ雙組分系統(tǒng)上調, 說明CpxRA雙組分系統(tǒng)可能和OmpR-EnvZ在功能上存在協(xié)調關系; 但CpxRA缺失對QseBC無顯著影響,QseBC雙組分系統(tǒng)與生物膜形成相關, 這和前面的實驗結果一致, 說明cpxRA基因簇缺失對細菌生物膜形成影響不大; 當加入誘導劑EDTA后, 菌株的部分基因表達如圖11所示, 經(jīng)EDTA處理0.5h后, 野生株和突變株dsbA/exbD1/histidine kinase gene (AHML_03870)/hly基因均上調表達, 其中有顯著差異的是exbD1和histidine kinase gene。
3 討論
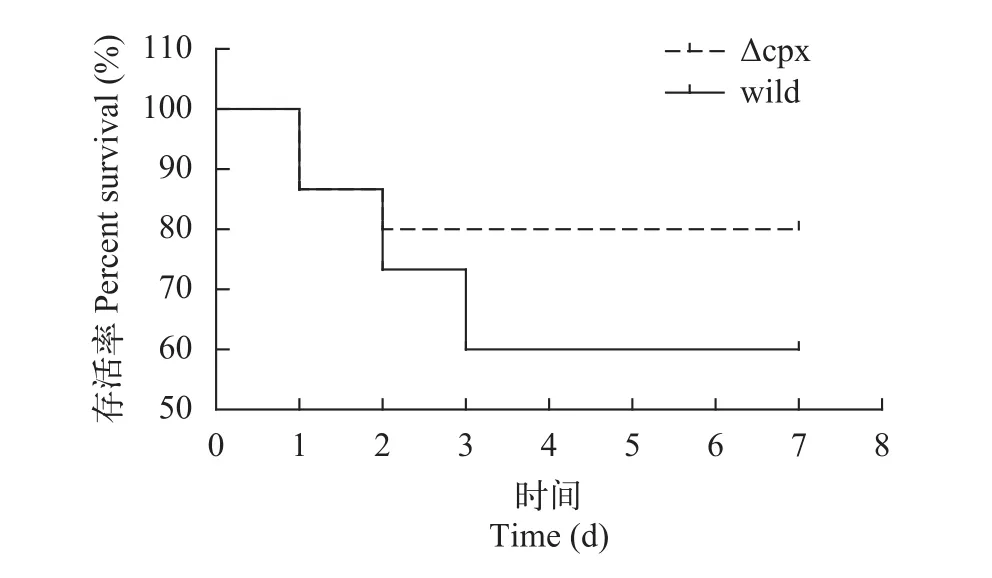
圖9 斑馬魚浸泡攻毒累積存活曲線Fig. 9 The survival curve of zebrafish attacked by Aeromonas hydrophila strains soak
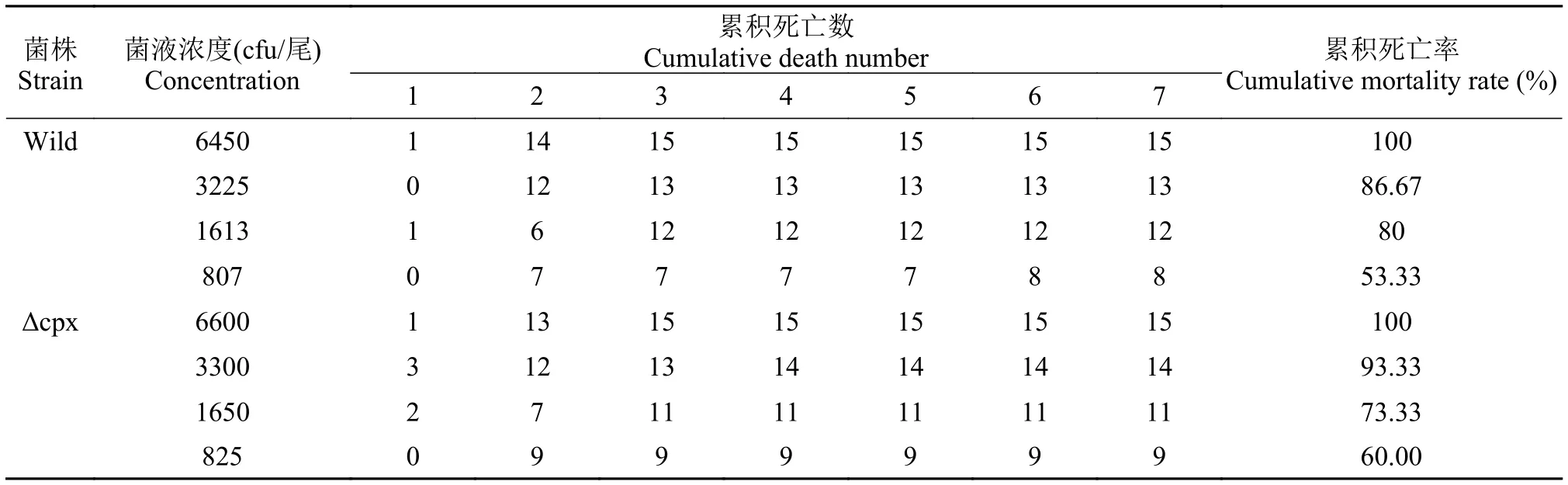
表 3 斑馬魚腹腔注射累積死亡情況Tab. 3 The cumulative death of zebrafish injected A. hydrophila strains by intraperitoneal
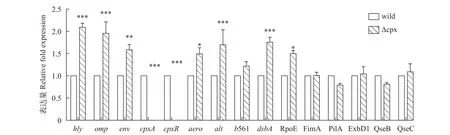
圖10 相關基因表達量變化Fig. 10 Related genes expression level
自從1980年McEwen等[16]研究大腸桿菌基因缺失株特征時發(fā)現(xiàn)CpxRA系統(tǒng), 這個雙組分系統(tǒng)逐漸受到研究者的關注, 它的生物功能在越來越多的細菌中被揭示。Cosma等[17]研究發(fā)現(xiàn)Cpx系統(tǒng)的激活能緩和膜蛋白的錯誤折疊; De Wulf等[18,19]認為cpx系統(tǒng)能使細菌獲得對不良應激的抵抗力; Gal-Mor等[20]研究表明Cpx系統(tǒng)可以調節(jié)嗜肺軍團菌(Legionella pneumophila)毒力基因icm和dot的表達;Jubelin等[21]研究發(fā)現(xiàn), Cpx系統(tǒng)可以調控大腸桿菌菌毛的合成, 間接影響病原菌的粘附和侵襲。本實驗通過融合PCR構建嗜水氣單胞菌CpxRA雙基因缺失片段, 利用自殺質粒pRE112和同源重組獲得cpx缺失突變株Δcpx來研究Cpx系統(tǒng)在嗜水氣單胞菌中的作用。
結果顯示突變株的外部形態(tài)以及在正常環(huán)境下攝取養(yǎng)分生長、運動、形成生物膜和野生株相比沒有顯著差異, 這說明cpxRA基因缺失對這些過程的影響程度較小; 斑馬魚腹腔注射實驗結果顯示野生株和缺失株相對于斑馬魚而言均為強毒株, 兩者毒力沒有明顯差異, 而在浸泡感染過程中, 缺失株毒力較野生株稍弱, 但差異還沒有達到顯著水平。因此我們推測cpxRA基因簇不是主要毒力基因。在這些結果中, 突變株和野生株表現(xiàn)一致, 根據(jù)qRT-PCR比較, 可能有2種原因, 一種是cpxRA基因簇確實不參與嗜水氣單胞菌的這些過程, 對毒力基因影響也不大; 另一種可能是CpxRA被敲除后,有其他系統(tǒng)取代了該系統(tǒng)在這些過程中的作用, 因而細菌表現(xiàn)與野生株無差異, 可以看到突變株的ompR、envZ雙組分系統(tǒng)基因以及經(jīng)EDTA誘導后類似于CpxA的組氨酸激酶(AHML_03870)是上調的, 這說明這些基因是受Cpx系統(tǒng)影響的[22]。
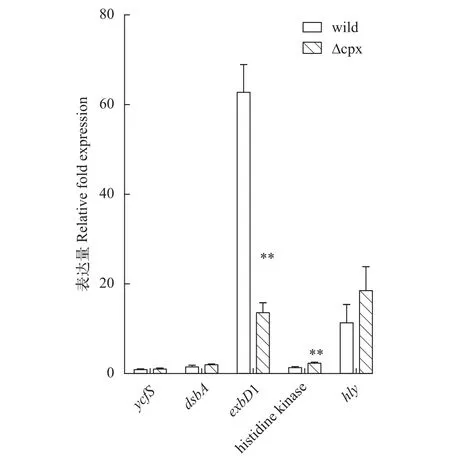
圖11 加入EDTA誘導劑后菌株熒光定量結果Fig. 11 The result of qRT-PCR after EDTA induction
在急性刺激實驗中, 突變株在0.5 mg/mL SDS刺激下存活率顯著高于野生株, 而在0.5 mol/L KCl的高滲透壓環(huán)境下則是顯著低于野生株的; 同樣的, 當給予慢性刺激時, 突變株在5 mg/mL的EDTA刺激下存活率顯著高于野生株, 在20 μg/mL多黏菌素B刺激下甚至停滯生長。這些結果說明Cpx雙組分系統(tǒng)參與嗜水氣單胞菌對外界刺激的應答, 且對不同刺激的表現(xiàn)不同。突變株在SDS刺激反應中存活率較野生株高, 這可能是因為Cpx系統(tǒng)增加細菌對SDS的敏感性, 所以當失去這個系統(tǒng)后, 細菌敏感性反而減弱, 但這也許會對突變株其他方面帶來不利; 突變株在高滲透條件下存活率顯著降低表明Cpx系統(tǒng)參與嗜水氣單胞菌抗高滲透壓的調控過程。對于EDTA刺激, 綜合急性和慢性刺激結果: 短時間的刺激沒有明顯降低菌株的存活率, 當刺激時間延長至12h, 野生株生長停滯, 突變株表現(xiàn)出緩慢生長, 由此說明5 mg/mL濃度的EDTA對菌株并不會帶來明顯致死效應, 但這種刺激環(huán)境的持續(xù)會干擾菌株的正常活動過程, 突變株由于缺少Cpx系統(tǒng),對這種刺激反應遲鈍, 因而生長較快, 但EDTA對細菌的累積損傷可能會造成突變株其他方面如離子吸收的改變。在熒光定量實驗中, 經(jīng)EDTA誘導后野生株exbD1基因的上調表達倍數(shù)高達62.7, 是突變株上調的3倍多,exbD1基因編碼的蛋白是ExbBExbD-TonB轉運系統(tǒng)的一部分, 該轉運系統(tǒng)是能量依賴的運輸外界陽離子進入機體進行生命活動的一個通道[23,24]。野生株在受刺激后大量表達exbD1基因, 有助于機體同EDTA爭奪陽離子, 為細菌的大量繁殖做準備。多黏菌素B刺激結果說明野生株DBHS101對20 μg/mL多黏菌素B有一定耐藥性, 突變株在這種條件下生長停滯可能是因為缺失Cpx雙組分系統(tǒng), 機體不能迅速啟動耐藥機制所以生長受阻, 由此我們推測Cpx系統(tǒng)參與多黏菌素B耐藥過程, 當然這還需要后續(xù)進一步驗證。本研究通過實驗說明敲除cpxRA基因簇并不影響嗜水氣單胞菌DBHS101毒力, 這與沙門氏菌[25]、大腸桿菌[26]等是不同的; 因此我們需要從嗜水氣單胞菌的不同雙組分系統(tǒng)間的關系及毒力基因表達譜方面繼續(xù)探究來分析原因; 此外比較缺失突變株在其他環(huán)境壓力、代謝等方面與野生株的差異來系統(tǒng)了解Cpx在嗜水氣單胞菌中發(fā)揮的功能也是后續(xù)值得討論的問題。
[1]Thune R L, Stanley L A, Cooper R K. Pathogenesis of gram-negative bacterial infections in warm water fish [J].Annual Reviews of Fish Diseases, 1993, 3: 145—185
[2]Popoff M. Aeromonas. Krieg N R, editor. Bergy’s Manual of Systematic Bacteriology, vol. 1. Baltimore [M]. Williams &Wilkins, 1984, 545—548
[3]Altwegg M, Geiss H K. Aeromonas as a human pathogen[J].Critical Reviews in Microbiology, 1989, 16: 253—286
[4]Austin B, Austin D A. Bacterial fish pathogens. Disease of Farmed and Wild Fish [M]. 5th ed Chichester: Springer Praxis, 2012, 119—146
[5]Cipriano R C.Aeromonas hydrophilaand Motile Aeromonad Septicemias of Fish [M]. Fish Disease Leaflet, 2001, 68
[6]Conrad M. Cross-scale information processing in evolution, development and intelligence [J].Biosystems, 1996,38(2—3): 97—109
[7]Raivio T L, Silhavy T J. Transduction of envelope stress in Escherichia coli by the Cpx two-component system [J].Journal of Bacteriology, 1997, 179(24): 7724—7733
[8]Zhang X J, Yang W M, Li T T,et al. The genetic diversity and virulence characteristics ofAeromonas hydrophilaisolated from fishponds with disease outbreaks in Hubei province [J].Acta Hydrobiologica Sinica, 2013,37(3): 458—466 [張旭杰, 楊五名, 李彤彤, 等. 湖北地區(qū)暴發(fā)病池塘中嗜水氣單胞菌的遺傳多樣性和毒力特征研究. 水生生物學報, 2013, 37(3): 458—466]
[9]Ho S N, Hunt H D, Horton R M,et al. Site-directed mutagenesis by overlap extension using the polymerase chainreaction [J].Gene, 1989, 77(1): 51—59
[10]Hu Y H, Liu C S, Hou J H,et al. Identification, characterization, and molecular application of a virulence-associated autotransporter from a pathogenic Pseudomonas fluorescens strain [J].Applied and Environmental Microbiology, 2009, 75(13): 4333—4340
[11]Kristin S, Emina C, Elke H,et al. Molecular and proteome analyses highlight the importance of the Cpx envelope stress system for acid stress and cell wall stability inEscherichia coli[J].Microbiology Open, 2016, 5(4): 582—596
[12]Li A H. The study of the drug resistance, resistance plasmid of fish pathogenic bacteria, and the antibacterial action of several drug in China [D]. Ph D thesis, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan.1998 [李愛華. 我國魚類病原菌耐藥性、耐藥質粒及幾種藥物抗菌作用的研究. 中國科學院水生生物研究所博士學位論文. 武漢. 1998]
[13]Acosta N, Pukatzki S, Raivio T L. TheVibrio choleraeCpx envelope stress response senses and mediates adaptation to low iron [J].Journal of Bacteriology, 2015, 197:262—276
[14]Raivio T L, Leblanc S K, Price N L,et al. The Escherichia coli Cpx envelope stress response regulates genes of diverse function that impact antibiotic resistance and membrane integrity [J].Journal of Bacteriology, 2013,195: 2755—2767
[15]Livak K J, Schmittgen T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-ΔΔCt)method [J].Methods, 2001, 25(4): 402—408
[16]McEwen J, Silverman P. Chromosomal mutations of Escherichia coli that alter expression of conjugative plasmid functions [J].Proceedings of the National Academy of Sciences of the United States of America, 1980, 77:513—517
[17]Cosma C L, Danese P N, Carlson J H,et al. Mutational activation of the Cpx signal transduction pathway ofEscherichia colisuppresses the toxicity conferred by certain envelope-associated stresses [J].Molecular Microbiology, 1995, 18: 491—505
[18]De W P, Lin E C. Cpx Two-Component Signal Transduction in Escherichia coli: Excessive CpxR-P levels underlie CpxA* phenotypes [J].Journal of Bacteriology, 2000,182(5): 1423—1426
[19]De W P, Kwon O, Lin E C. The CpxRA signal transduction system ofEscherichia coli: Growth-related autoactivation and control of unanticipated target operons [J].Journal of Bacteriology, 1999, 181(21): 6772—6778
[20]Gal-Mor O, Segal G. Identification of cpxR as a positive regulator of icm and dot virulence genes ofLegionella pneumophila[J].Journal of Bacteriology, 2003, 185(16):4908—4919
[21]Jubelin G, Vianney A, Beloin C,et al. Cpx R/Omp R interplay regulates curli gene expression in response to osmolarity inEscherichia coli[J].Journal of Bacteriology,2005, 187(6): 2038—2049
[22]Gerken H, Misra R. MzrA-EnvZ interactions in the periplasm influence the EnvZ/OmpR two-component regulon[J].Journal of Bacteriology, 2010, 192(23): 6271—6278
[23]Blanvillain S, Meyer D, Boulanger A,et al. Plant carbohydrate scavenging through tonB-dependent receptors: a feature shared by phytopathogenic and aquatic bacteria[J].PLoS One, 2007, 2: e224
[24]Schauer K, Gouget B, Carrière M,et al. Novel nickel transport mechanism across the bacterial outer membrane energized by the TonB/ExbB/ExbD machinery [J].Molecular Microbiology, 2007, 63: 1054—1068
[25]Humphreys S, Rowley G, Stevenson A,et al. Role of the two-component regulator Cpx AR in the virulence of Salmonella entetica serotype typhimurium [J].Infection andImmunity, 2004, 72(8): 4654—4661
[26]Debnath I, Norton J P, Barber A E,et al. The Cpx stress response system potentiates the fitness and virulence of uropathogenicEscherichia coli[J].Infection and Immunity, 2013, 81(5): 1450—1459
猜你喜歡
工業(yè)設計(2022年8期)2022-09-09 07:43:20
軍民兩用技術與產(chǎn)品(2021年10期)2021-03-16 06:05:30
北京測繪(2020年12期)2020-12-29 01:33:58
裝備制造技術(2019年12期)2019-12-25 03:06:46
制造技術與機床(2019年10期)2019-10-26 02:47:06
中國洗滌用品工業(yè)(2019年4期)2019-05-11 09:27:34
鐵道通信信號(2018年5期)2018-06-28 03:06:24
家庭影院技術(2017年9期)2017-09-26 03:41:45
知識經(jīng)濟·中國直銷(2017年5期)2017-06-15 20:28:19
通信電源技術(2016年6期)2016-04-20 06:21:32
