不同魚類肌間骨的骨化模式研究
2018-01-24 02:24:30聶春紅陳祖萱戴彩嬌萬世明高澤霞
水生生物學報 2018年1期
聶春紅 陳祖萱 戴彩嬌 萬世明 高澤霞,
(1. 華中農業大學水產學院, 農業部淡水生物繁育重點實驗室/農業動物遺傳育種與繁殖教育部重點實驗室, 武漢 430070;2. 淡水水產健康養殖湖北省協同創新中心, 武漢 430070; 3. 池塘健康養殖湖北省工程實驗室, 武漢 430070)
肌間骨(Intermucsular bone, IB)由肌膈中的間充質細胞骨化而來, 是一種膜性硬骨, 根據附著部位可分為髓弓小骨、脈弓小骨、椎體小骨三種類型[1,2]。肌間骨的存在與魚類的分類地位有極大的關系, 其僅存在低等真骨魚類中, 系統演化過程表現為數目由少到多, 多到少, 再到無的過程[2—4]。近年來有關魚類肌間骨的研究大多集中在研究肌間骨的數目及形態, 如Patterson和Johnson[2]對125屬真骨魚的代表種肌間骨進行了詳細的觀察和分析; 呂耀平等[3]對骨舌魚總目、海鰱總目、鯡形總目和骨鰾總目等低等真骨魚類中的代表性魚類的肌間骨數量、形態和分布進行了比較分析; 董在杰等[5]對幾種常見的鯉科養殖魚類肌間骨形態及數目進行了系統的分析。然而, 對魚類肌間骨的骨化過程研究的較少。在鯉科魚類上, 僅秉志[6]、Bird和Mabee[7]、柯中和等[8]、萬世明等[9]、呂耀平等[10,11]分別研究了鯉(Cyprinus carpio)、斑馬魚(Danio rerio)、鰱(Hypophthalmichthys molitrix)、團頭魴(Megalobrama amblycephala)、唇逕(Hemibarbus labeo)和甌江彩鯉(Cyprinus carpio var. color)等魚類的肌間骨骨化過程, 結果顯示均是從尾部向頭部發育。鯉科魚類肌間骨的形態比較復雜, 基本被分為6種形態, 且其他復雜的形態都是由“I”形演化而來的。姚文杰等[12,13]對日本鰻鱺(Anguilla japonica)肌間骨骨化過程研究卻發現其是從頭部向尾部發育, 這與已報道的鯉科魚類的截然相反, 據此推測魚類肌間小骨的骨化可能與其不同的游動方式有關。
為進一步探討魚類肌間骨的骨化方式是否與游動方式相關, 或者是與魚類的進化相關, 鯉科魚類表現為從尾部向頭部的骨化方向, 而非鯉科魚類的日本鰻鱺卻表現為從頭部向尾部的骨化方向。本研究選擇了合鰓魚目合鰓魚科的黃鱔(Monopte-rus albus)和鯉形目鰍科的泥鰍(Misgurnus anguillicaudatus), 系統研究了2種魚類的肌間骨發生發育過程。此外, 本研究還采用成骨細胞特異性轉錄因子(osterix)綠色轉基因熒光蛋白斑馬魚, 對其肌間骨的發生發育進行了活體的系統觀察。研究結果將為魚類肌間骨的發生發育模式的探討提供更豐富的材料, 為今后研究魚類肌間骨發生發育的分子機制奠定基礎。
1 材料與方法
1.1 實驗材料
黃鱔樣本來自荊州長江水產研究所的友情提供, 仔、稚魚樣本從受精卵孵出后開始采集, 每隔5d采集一次樣本, 每次5—10尾, 一直采集到孵出后60d; 黃鱔成體為一齡魚。泥鰍樣本來自華中農業大學水產學院養殖基地自繁群體, 樣本采集從受精卵孵出后開始一直到孵出后40d, 每隔3天采集一次,每次5—10尾; 泥鰍成體為一齡魚。
1.2 骨骼染色方法
幼體染色方法: 整體骨骼染色的方法參照柯中和等[8]方法, 但其步驟稍有改動。用MS-222將魚麻醉(100 mg/L)后用95%的酒精保存標本; 標本經75%、50%和30%的酒精梯度復水, 每次15—20min;轉移至1%雙氧水中漂白30—60min, 蒸餾水浸泡30min左右, 1% KOH溶液中透明20—30min; 后轉移至新的1% KOH溶液中, 加適量的茜素紅染液, 直至溶液變為深紫色, 染色20min左右后將標本轉移至新的1%KOH溶液中, 洗去標本表面的染色液; 然后將標本轉移至50%的甘油中, 直至體表的染色褪去; 最后將標本放在100%的甘油中保存。
成體染色方法: 整體骨骼染色的方法參照柯中和等[8]方法, 但其步驟稍有改動。用MS-222將魚麻醉(200 mg/L)后用95%的酒精保存標本; 經50%和30%酒精梯度復水; 將標本轉移至1%的雙氧水中漂白10h左右后轉移至蒸餾水過夜處理; 轉移至1%KOH溶液中透明8h左右, 后轉移至新的1% KOH溶液中, 加適量的茜素紅染液, 直至溶液變為深紫色,染色至少6h, 直至骨骼被染成紫紅色后將標本轉移至新的1%KOH溶液中, 洗去標本表面的染色液; 然后將標本轉移至50%的甘油中, 直至體表的染色褪去; 最后將標本放在100%的甘油中保存。
骨骼染色標本通過SZX-16型日本奧康體視顯微鏡觀察和拍照, 記錄肌間骨數目、位置及形態。
1.3 轉基因斑馬魚肌間骨發育觀察
成骨細胞特異性轉錄因子綠色轉基因熒光蛋白(Osterix GFP)斑馬魚親本由臺灣中原大學蕭崇德教授提供, 本實驗室繁殖子代后進行觀察。自孵出后開始一直到孵出后30d(體長20 mm), 每隔3d選取5—10尾幼魚, 用MS-222將魚麻醉(100 mg/L)后, 放置于體式熒光顯微鏡(Leica, M205 FA)下進行觀察拍照。斑馬魚成體染色參照1.2中描述的成體染色方法進行。
2 結果
2.1 黃鱔的肌間骨骨化過程
黃鱔幼魚孵出25d (體長約35 mm)前肌間骨還沒有出現, 但其主軸骨骼及附肢骨骼已骨化完成(圖1A); 在孵出30d (體長約40 mm)時, 肌間骨從頭部開始少量出現, 形態為“I”形, 且只有椎體小骨(圖1B); 在孵出45d (體長約55 mm)時, 黃鱔的主軸骨骼進一步發育, 椎體小骨從頭部向尾部開始大量出現(圖1C); 在孵出55d (體長約100 mm), 肌間骨基本全部出現, 形態清晰, 附著在椎體上(圖1D)。由此可見黃鱔的椎體小骨的骨化順序是從頭部向尾部骨化的, 形態僅有“I”形, 無其他形態的肌間骨發育。此外, 黃鱔成魚經骨骼染色后, 其肌間骨的形態及位置可以被清楚地觀察到(圖2A), 僅有“I”形的椎體小骨, 成體黃鱔共有180—188根小骨, 肌節數為150—166, 平均每一肌節的肌間骨數目為1.08—1.25。李仲輝[14]在對黃鱔的骨骼研究中對黃鱔的骨骼做了詳細的觀察, 發現黃鱔僅存在上肋骨,沒有下肋骨。為了區別黃鱔上肋骨與椎體小骨, 圖3顯示了成體黃鱔的椎體小骨與上肋骨分別存在的位置。
2.2 泥鰍的肌間骨骨化過程
泥鰍幼魚在孵出后24d (體長15 mm), 其骨骼包括主軸骨骼及附肢骨骼已骨化完全, 但肌間骨還沒有出現(圖4A)。孵出后27d (體長17 mm), 肌間骨從尾部開始出現, 有1—2根, 形態為“I”形(圖4B),為脈弓小骨。泥鰍孵化33d左右(體長20 mm), 大量肌間骨已出現在尾部, 包括髓弓小骨和脈弓小骨,但沒有發育到背部(圖4C)。孵出后37d (體長25 mm), 肌間骨出現于背鰭下方, 形態都為“I”形。孵出后40d (體長35 mm), 肌間骨全部出現, 尾部肌間骨形態發生分化, 少量的“卜”形出現(圖4D)。此外, 泥鰍成魚骨骼經染色后觀察發現其具有髓弓小骨和脈弓小骨(圖2B), 肌間骨的形態有“I”、“Y”和“卜”形三種形態; 成體泥鰍肌間骨的數目約100—104根, 肌節數為40—44, 平均每肌節的肌間骨數目為2.27—2.60。
2.3 熒光斑馬魚的肌間骨骨化過程
Osterix綠色轉基因熒光蛋白斑馬魚幼魚孵出后15d (體長5 mm), 魚體主軸骨骼已發育完全, 且肌間骨并沒有出現(圖5A); 孵出后21d (體長6.5 mm)時, 肌間骨從尾部開始出現, 形態為“I”形, 為脈弓小骨(圖5B); 孵出后27d (體長10 mm)時, 肌間骨在尾部大量出現, 但還沒有發育到背部, 髓弓小骨與脈弓小骨均出現(圖5C); 孵出后30d (體長15 mm), 肌間骨已生長到背鰭下方, 且形態開始復雜化發育(圖5D)。隨著熒光斑馬魚的生長, 鱗片的出現覆蓋了骨骼的熒光。因此, 我們利用整體骨骼染色方法對成魚的斑馬魚進行骨骼染色觀察(圖2C), 結果顯示斑馬魚存在髓弓小骨和脈弓小骨兩種類型, 肌間骨的形態有“I”、“卜”、“Y”和一端多叉形。

圖2 黃鱔、泥鰍和斑馬魚成魚骨骼染色顯示成魚肌間骨的形態Fig. 2 Whole-mount staining of skeleton for M. albu (A), M. anguillicaudatus (B) and D. rerio (C) to show different morphological types of intermuscular bones
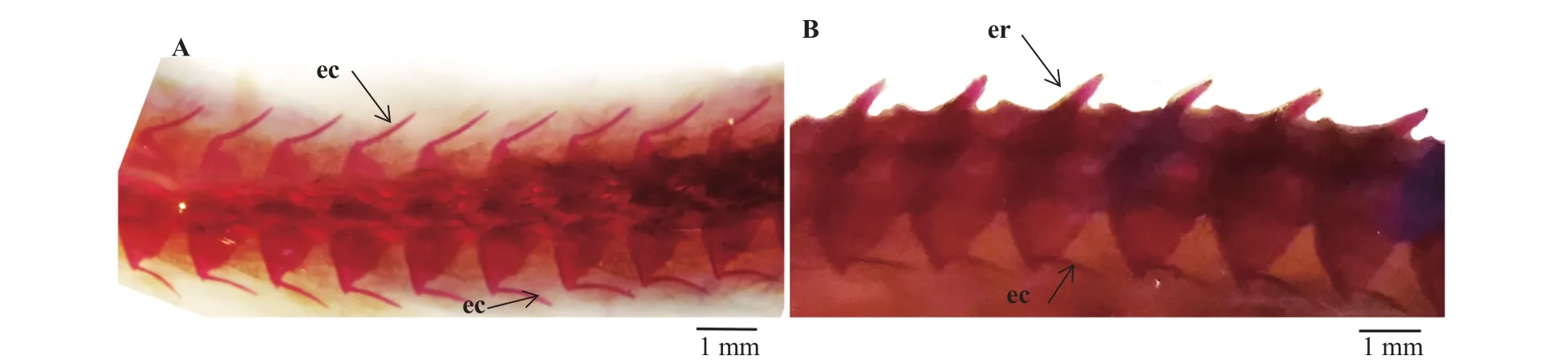
圖3 成體黃鱔椎體小骨和上肋骨的位置差異Fig. 3 Position difference of epicentrals and epipleural ribs in M. albu
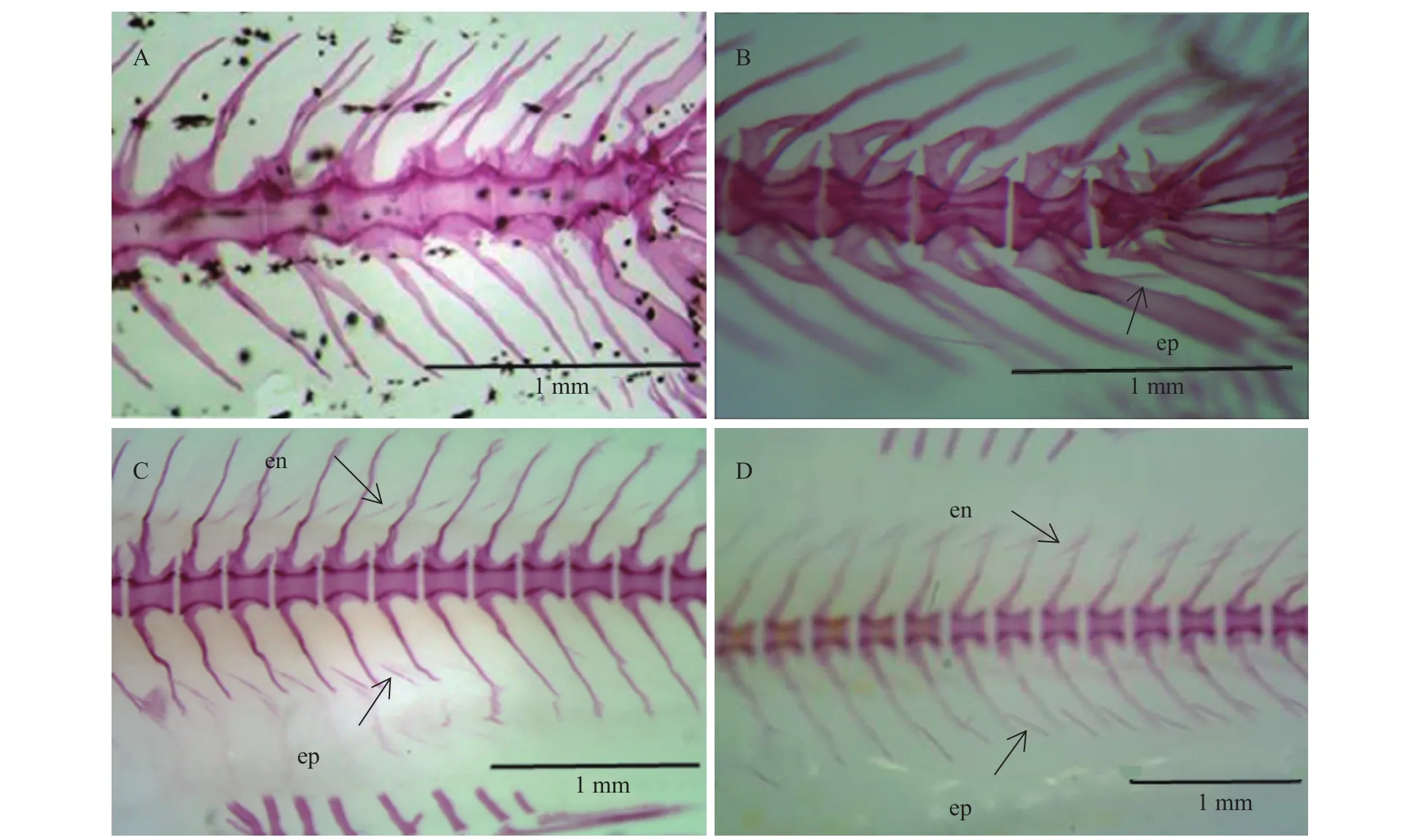
圖4 不同發育時期泥鰍肌間骨骨化過程Fig. 4 The ossification process of intermuscular bones at different stages of Misgurnus anguillicaudatus
3 討論
魚類骨骼的骨化是一個復雜的過程, 不同魚類有不同的骨化模式。本文研究了3種魚類的肌間骨骨化模式, 黃鱔與泥鰍和斑馬魚的骨化模式是相反的。泥鰍屬于鯉形目鰍科花鰍亞科, 斑馬魚屬于鯉科魚丹亞科, 兩種魚的肌間骨的骨化順序是從尾部向頭部依次出現, 有髓弓小骨和脈弓小骨。這種肌間骨骨化方式與已報道的分別屬于鯉科的鲌亞科團頭魴、逗亞科唇魚骨和鰱亞科鰱的肌間骨骨化順序是一致的[7—10]。黃鱔, 屬于合鰓魚目, 合鰓魚科, 肌間骨的骨化順序是從頭向尾部依次骨化的;這與已報道的日本鰻鱺的骨化順序是一致的[12]。表 1是近年來所有已研究魚類的肌間骨骨化模式。由此可見, 泥鰍和斑馬魚、團頭魴、唇魚骨、鰱都屬于鯉形目, 分屬鰍科和鯉科, 但具有相同的肌間骨骨化順序。而非鯉形目的日本鰻鱺表現為從頭部向尾部依次骨化的肌間骨骨化方式, 姚文杰等[12]據此推測魚類肌間骨的骨化可能與其不同的游動方式有關。本文研究的黃鱔的游動方式與日本鰻鱺相同, 都屬于鰻鱺模式, 而其他已報到肌間骨從尾向頭部骨化的魚類的游動方式均為鲹科模式。本文結果進一步證實了魚類肌間骨的骨化可能與其不同的游動方式有關。但另一方面, 因為目前已報道的肌間骨從尾向頭部骨化的魚類都屬于鯉形目, 而肌間骨從頭部向尾部骨化的黃鱔和日本鰻鱺分別屬于合鰓魚目和鰻鱺目, 因此魚類肌間骨骨化方式是否與進化有一定關系, 還需要進一步采集更多屬于不同分類地位的魚類進行更全面的研究。

圖5 不同發育時期斑馬魚肌間骨骨化過程Fig. 5 The ossification process of intermuscular bones at different stages of Danio rerio
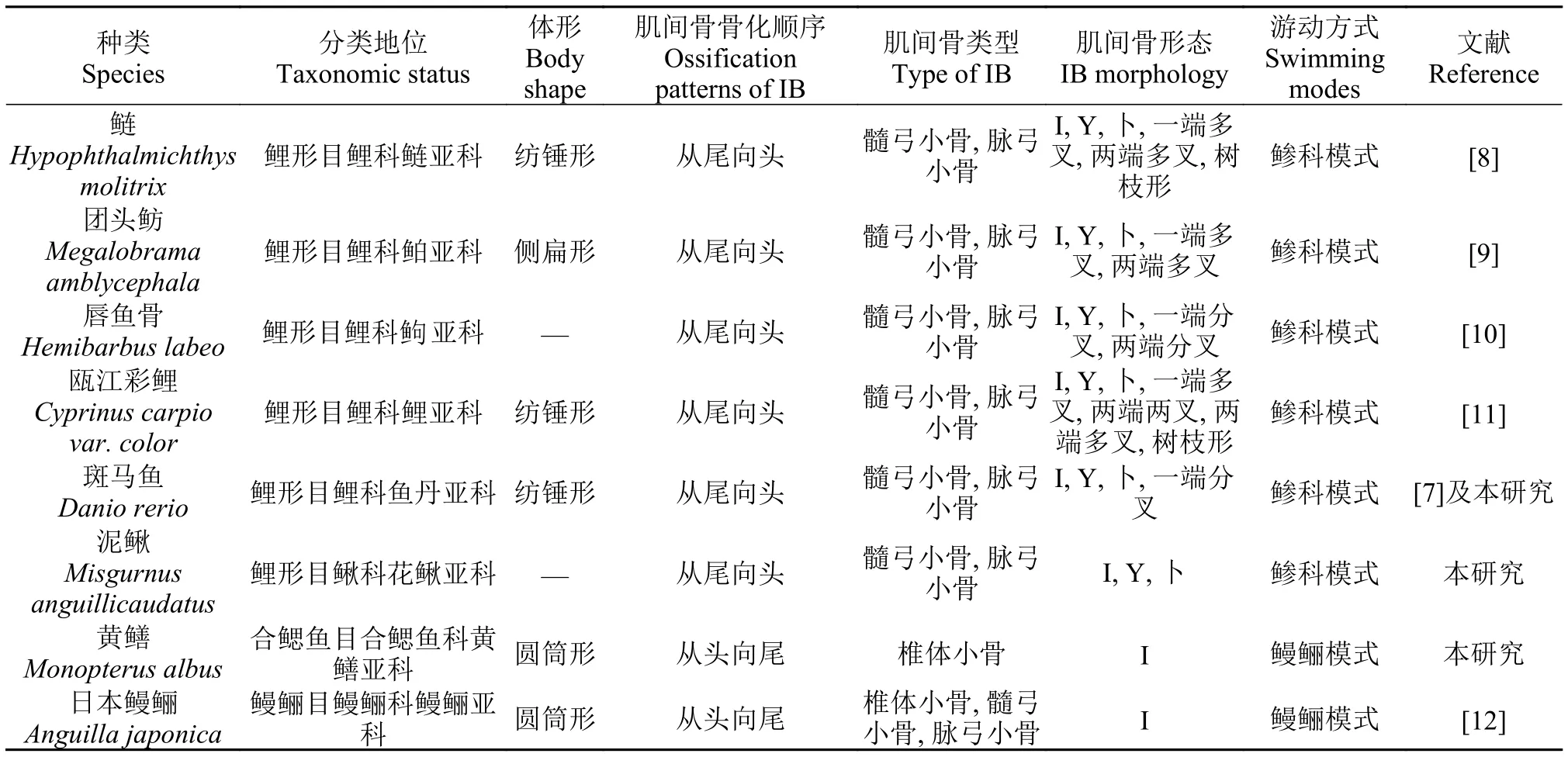
表 1 本文及已報道的不同魚類肌間骨骨化模式的比較Tab. 1 The documents about ossification patterns of intermuscular bones among different fish species in this study and other previous reports
呂耀平等[3]和蔣燕等[15]分別對骨舌魚總目、海鰱總目、鯡形總目和骨鰾總目等低等真骨魚類肌間骨的類型分析結果顯示, 在低等真骨魚類的系統演化過程中, 骨舌魚總目只有髓弓小骨和脈弓小骨,不包含椎體小骨, 而海鰱總目與鯡形總目的魚類含有三種類型的肌間骨, 而隨后的骨鰾總目中椎體小骨已退化。由此可見, 椎體小骨在魚類進化上出現的時間比髓弓小骨和脈弓小骨要晚, 但其退化比髓弓小骨和脈弓小骨卻早很多。本研究中的泥鰍和斑馬魚屬于骨鰾總目, 僅有髓弓小骨和脈弓小骨存在, 與上述研究結果一致。黃鱔屬于棘鰭總目, 卻含有椎體小骨, 而無髓弓小骨和脈弓小骨, 表明椎體小骨在骨鰾總目退化后又在棘鰭總目中出現, 這與蔣燕等[15]報道的椎體小骨退化比髓弓小骨要早很多, 且椎體小骨的演化史要比髓弓小骨和脈弓小骨短得多的結論是相悖的, 表明椎體小骨的演化史還需要進一步探討。
肌間骨是存在肌膈間的一種膜性小骨, 其形態表現出多樣性[16]。呂耀平等[3]將肌間骨(髓弓小骨和脈弓小骨)的形態分為有“I”形、“卜”形、“Y”形、一端多叉形、兩端兩叉形、兩端多叉形、樹枝形等七種形態, 認為肌間骨的復雜形態是由“I”形演化而來的, 且從骨舌魚總目到骨鰾總目, 肌間骨形態表現為從簡單向復雜形態演化, 然后出現退化的現象。本文研究的泥鰍的肌間骨形態有“I”、“卜”和“Y”形三種形態, 與呂耀平等[3]報道的泥鰍的肌間骨研究結果一致。黃鱔的椎體小骨僅有“I”形,與日本鰻鱺的肌間骨形態一致[12], 但海鰱總目中海鰻的椎體小骨的形態相對復雜, 包括“I”、“Y”和一端多叉形[3], 表明魚類椎體小骨形態經歷了從簡單到復雜, 再退化到簡單的演化過程。此外, 從已報道的魚類肌間骨的形態來看, 黃鱔、日本鰻鱺、海鰻等體型為圓筒形且游動方式為鰻鱺模式的魚類相較于鰱、團頭魴、甌江彩鯉等體型為紡錘形或側扁形且游動方式為鲹科模式的魚類, 肌間骨的形態類型要簡單得多, 絕大部分以“I”形為主。因此,推測魚類肌間骨的形態與其體型和游動方式可能有一定的關系。
此外, 本文首次采用成骨細胞特異性轉錄因子(osterix)綠色轉基因熒光蛋白斑馬魚, 對其肌間骨的發生發育進行了活體的系統觀察, 結果顯示肌間骨發生發育過程都可以通過此熒光魚觀察到, 這為今后通過斑馬魚在活體水平研究基因在肌間骨發生發育中的功能奠定了很好的材料基礎。以此嘗試通過基因敲除、遺傳改良、添加基因抑制劑等技術抑制肌間骨形成, 從分子水平上探討培育高產、優質、肌間刺少的養殖新品種的技術手段。
[1]Meng Q W, Su Q W, Li W D. Comparative Anatomy of Fishes [M]. Beijing: Science Press. 1987, 102—103 [孟慶聞, 蘇錦祥, 李婉端. 魚類比較解剖. 北京: 科學出版社. 1987, 102—103]
[2]Patterson C, Johnson G D. The intermuscular bones and ligaments of teleostean fishes [J].Smithsonian Contribution to Zoology, 1995, 559: 1—85
[3]Lü Y P, Bao B L, Jiang Y,et al. Comparative analysis of intermuscular bones in lower teleosts [J].Journal of Fisheries of China, 2007, 31(5): 661—668 [呂耀平, 鮑寶龍,蔣燕, 等. 低等真骨魚類肌間骨的比較分析. 水產學報,2007, 31(5): 661—668]
[4]Nelson J S. Fishes of the World (Third edition) [M]. John Wiley & Sons, Inc., New York, 1994, 600
[5]Dong Z J, Huang D Z, Li L J,et al. Preliminary study on intermuscular bones of several cultured cyprinids [J].Journal of Shanghai Fisheries University, 2006, 15(4):425—429 [董在杰, 黃代中, 李麗娟, 等. 幾種常見鯉科魚類肌間刺的初步研究. 上海水產大學學報, 2006,15(4): 425—429]
[6]Bing Z. On the myoseptal spines of the carp (Cyprinus carpioL.) [J].Acta Zoologica Sinica, 1962, 14(2):175—178 [秉志. 幼鯉大側肌肌隔骨針的觀察. 動物學報, 1962, 14(2): 175—178]
[7]Bird N C, Mabee P M. Developmental morphology of the axial skeleton of the zebrafish,Danio rerio(Ostariophysi:Cyprinidae) [J].Developmental Dynamics, 2003, 228(3):337—357
[8]Ke Z H, Zhang W, Jiang Y,et al. Developmental morphology of the intermuscular bone inHypophthalmichthys molitrix[J].Chinese Journal of Zoology, 2008,43(6): 88—96 [柯中和, 張煒, 蔣燕, 等. 鰱肌間小骨發育的形態學觀察. 動物學雜志, 2008, 43(6): 88—96]
[9]Wan S M, Yi S K, Zhong J,et al. Developmental and morphological observation of intermuscular bones inMegalobrama amblycephala[J].Acta Hydrobiologica Sini-ca, 2014, 38(6): 1144—1152 [萬世明, 易少奎, 仲嘉, 等.團頭魴肌間骨發育的形態學觀察. 水生生物學報, 2014,38(6): 1144—1152]
[10]Lü Y P, Chen J, Bao B L,et al. The ossificational process of the intermuscular bones inHemibarbus labeo[J].Journal of Shanghai Fisheries University, 2012, 21(4):549—553 [呂耀平, 陳潔, 鮑寶龍, 等. 唇逕肌間小骨的骨化過程. 上海海洋大學學報, 2012, 21(4): 549—553]
[11]Lü Y P, Zhou C C, Yang L M,et al. Ossification pattern of the intermuscular bone inCyprinus carpiovar. color[J].Journal of Shanghai Ocean University, 2014, 23(1):58—63 [呂耀平, 周晨晨, 楊麗美, 等. 甌江彩鯉肌間小骨的骨化模式. 上海海洋大學學報, 2014, 23(1):58—63]
[12]Yao W J, Gong X L, Lü Y P,et al. The ossificational process of the intermuscular bones inAnguilla japonica[J].Journal of Shanghai Ocean University, 2015, 23(6):810—813 [姚文杰, 龔小玲, 呂耀平, 等. 日本鰻鱺肌間小骨的骨化過程. 上海海洋大學學報, 2015, 23(6):810—813]
[13]Yao W, Lü Y, Gong X,et al. Different ossification patterns of intermuscular bones in fish with different swimming modes [J].Biology Open, 2015, 4(12): 1727—1732
[14]Li Z H. Study on the bones ofMonopterus albus[J].Zoological Research, 1981, 2(3): 215—222 [李仲輝. 黃鱔(Monopterus albus(Zuiew)) 骨骼的研究. 動物學研究,1981, 2(3): 215—222]
[15]Jiang Y, Yang L L, Bao B L. The epicentrals in several lower teleosts [J].Journal of Shanghai Fisheries University, 2008, 21(4): 549—553 [蔣燕, 楊琳琳, 鮑寶龍.幾種低等魚類的椎體小骨. 上海海洋大學學報, 2008,21(4): 549—553]
[16]Li Z, Zhou L, Wang Z W,et al. Comparative analysis of intermuscular bones between clone A+and clone F strains of allogynogenetic gibel carp [J].Acta Hydrobiologica Sinica, 2017, 41(4): 860—869 [李志, 周莉, 王忠衛, 等.異育銀鯽A+系和F系肌間骨的比較分析. 水生生物學報, 2017, 41(4): 860—869]
